
Quantitative Analysis of Cell Aggregation Dynamics Identifies HDAC Inhibitors as Potential Regulators of Cancer Cell Clustering

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Clustering Assay
2.3. Time-Lapse Microscopy
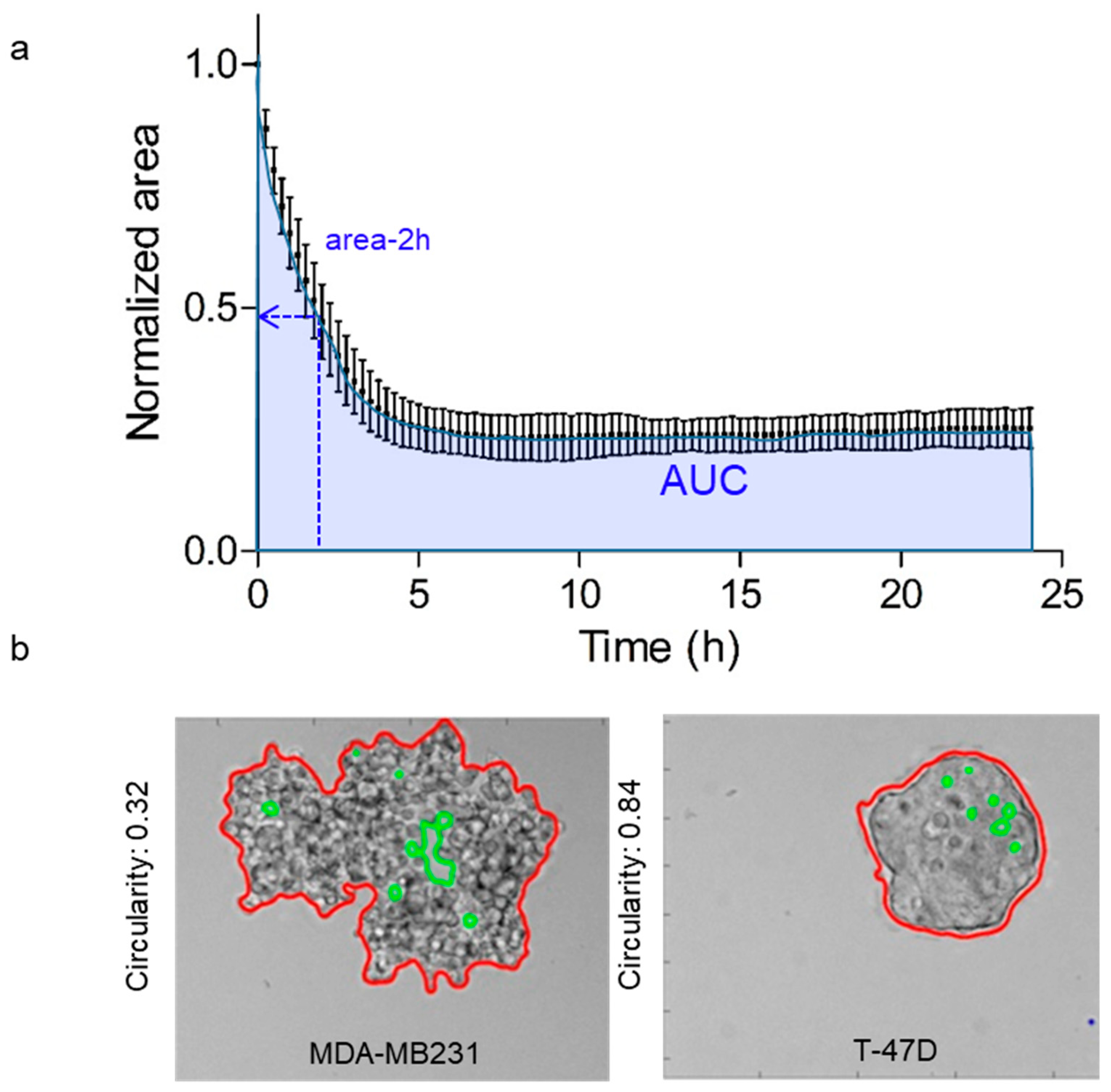
2.4. Quantification of Cell Clustering Parameters
- For each position/well/time, the whole stack of 2D images is fused into one single in-focus image. This process is based on a Laplacian of Gaussian filter to detect the in-focus image regions, followed by a Gaussian blending of these regions taken from the different focal planes.
- The background signal of the resulting in-focus image is estimated using a morphological opening and then subtracted.
- A Gaussian smoothing filter is then applied and the image intensities adjusted (histogram equalization performed on the pixel intensity to obtain a saturation of 1% of the extreme values), followed by a simple automated thresholding, resulting in a binary mask that corresponds to all cell aggregates.
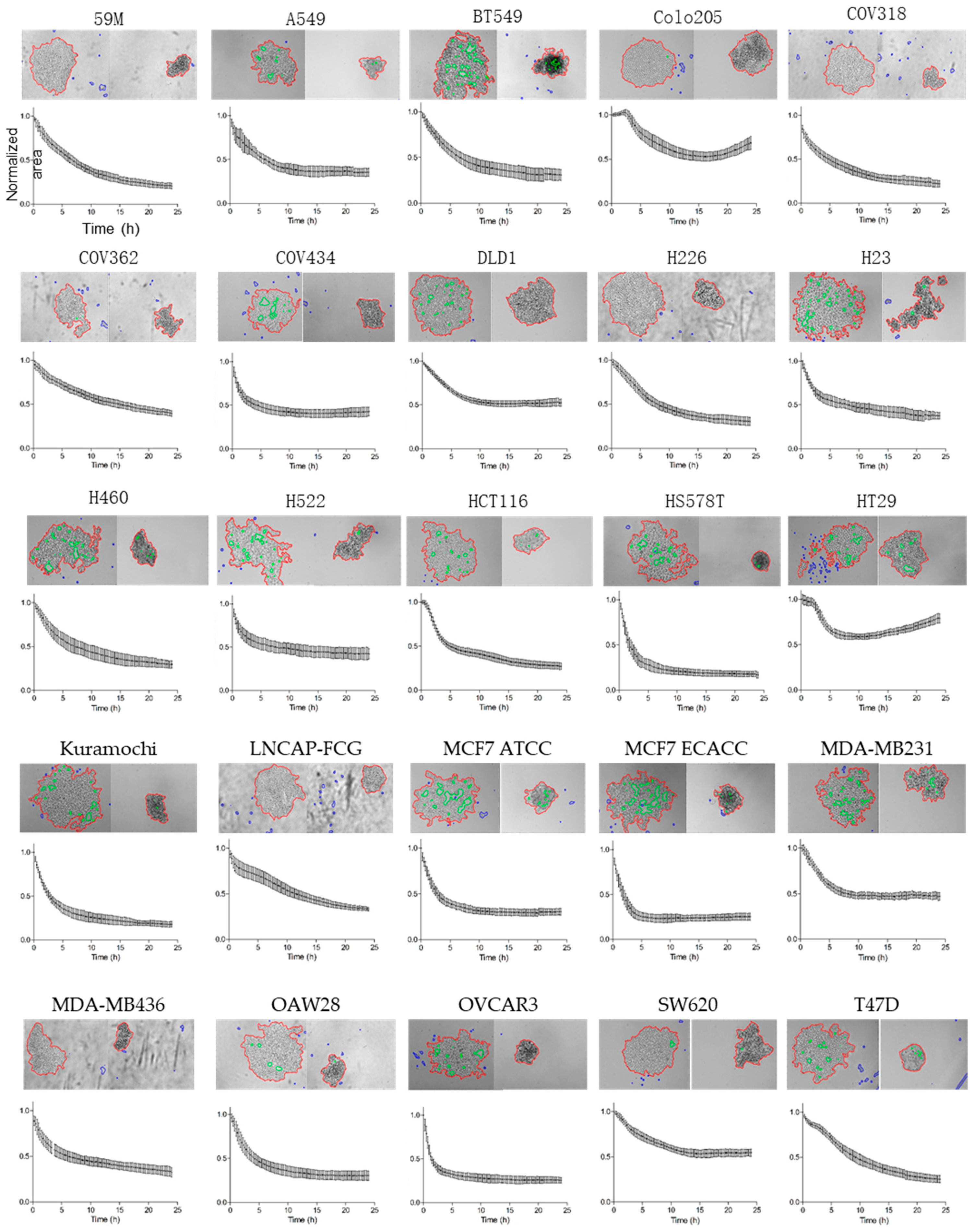
- Cell aggregates are individually detected in the binary image as connected components that match several criteria, and are divided in two groups according to their area using a threshold of 10,000 pixels (or 4160 µm2, because the XY resolution is 0.645 µm at 10× magnification). Detected objects with an area smaller than this threshold are excluded from the analysis. Holes inside the binary objects are excluded in the two object categories to measure standard parameters: number of aggregates, normalized area to the initial time point, and circularity. This image analysis pipeline was developed within a parallelized processing architecture using multi-core processors.
2.5. Analysis of the Relation between Cell Aggregation Parameters and CCLE Transcriptional Data
2.6. Connectivity Map Analysis
2.7. Statistical Analysis
3. Results
3.1. Investigation of the Aggregation Parameters in a Panel of 25 Cancer Cell Lines
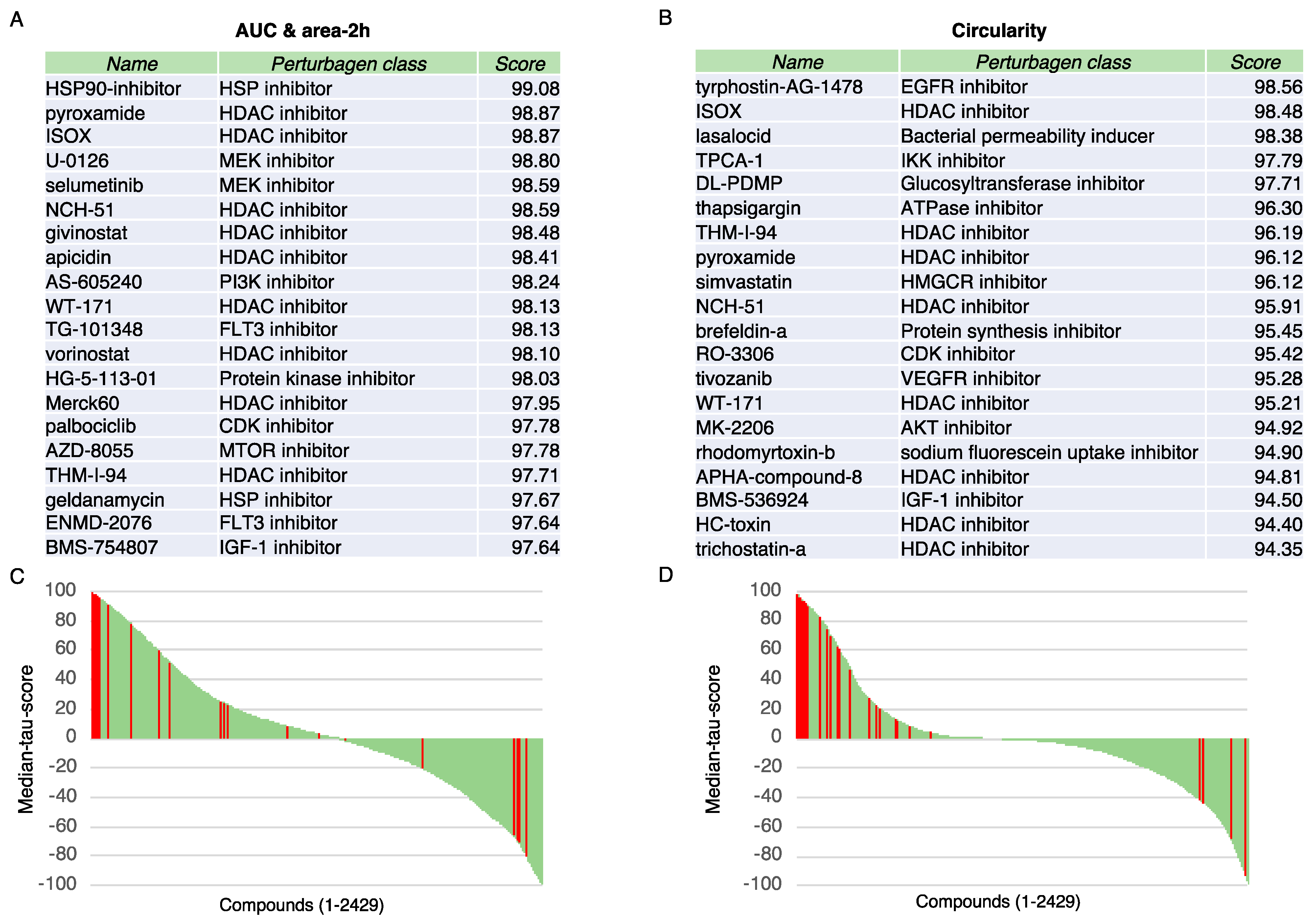
3.2. Identification of Genes Associated with the Aggregation Quantitative Parameters
3.3. Connectivity Map Analysis Identifies Strong Relationship with HDAC Inhibitors
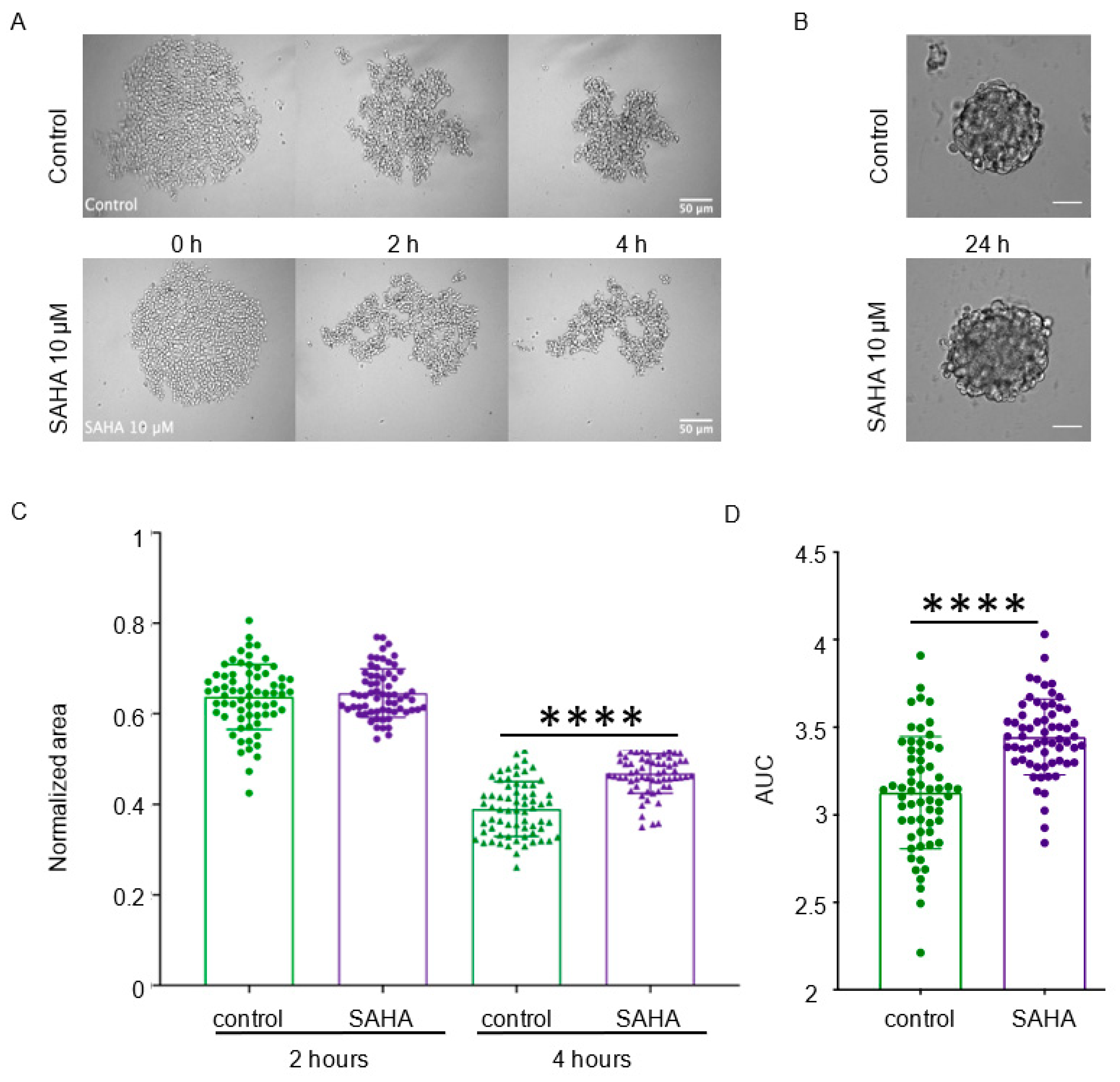
3.4. Effect of HDAC Inhibitors on In Vitro Cancer Cell Aggregation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro-Giner, F.; Aceto, N. Tracking cancer progression: From circulating tumor cells to metastasis. Genome Med. 2020, 12, 31. [Google Scholar] [CrossRef] [Green Version]
- Kelley, S.O.; Pantel, K. A New Era in Liquid Biopsy: From Genotype to Phenotype. Clin. Chem. 2020, 66, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Chitty, J.L.; Filipe, E.C.; Lucas, M.C.; Herrmann, D.; Cox, T.R.; Timpson, P. Recent advances in understanding the complexities of metastasis. F1000Research 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Aceto, N.; Bardia, A.; Miyamoto, D.T.; Donaldson, M.C.; Wittner, B.S.; Spencer, J.A.; Yu, M.; Pely, A.; Engstrom, A.; Zhu, H.; et al. Circulating tumor cell clusters are oligoclonal precursors of breast cancer metastasis. Cell 2014, 158, 1110–1122. [Google Scholar] [CrossRef] [Green Version]
- Szczerba, B.M.; Castro-Giner, F.; Vetter, M.; Krol, I.; Gkountela, S.; Landin, J.; Scheidmann, M.C.; Donato, C.; Scherrer, R.; Singer, J.; et al. Neutrophils escort circulating tumour cells to enable cell cycle progression. Nature 2019, 566, 553–557. [Google Scholar] [CrossRef]
- Foty, R.A.; Steinberg, M.S. Measurement of tumor cell cohesion and suppression of invasion by E- or P-cadherin. Cancer Res. 1997, 57, 5033–5036. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topal, B.; Roskams, T.; Fevery, J.; Penninckx, F. Aggregated colon cancer cells have a higher metastatic efficiency in the liver compared with nonaggregated cells: An experimental study. J. Surg. Res. 2003, 112, 31–37. [Google Scholar] [CrossRef]
- Updyke, T.V.; Nicolson, G.L. Malignant melanoma cell lines selected in vitro for increased homotypic adhesion properties have increased experimental metastatic potential. Clin. Exp. Metastasis 1986, 4, 273–284. [Google Scholar] [CrossRef]
- Tabaries, S.; Siegel, P.M. The role of claudins in cancer metastasis. Oncogene 2017, 36, 1176–1190. [Google Scholar] [CrossRef]
- Schuster, E.; Taftaf, R.; Reduzzi, C.; Albert, M.K.; Romero-Calvo, I.; Liu, H. Better together: Circulating tumor cell clustering in metastatic cancer. Trends Cancer 2021, 7, 1020–1032. [Google Scholar] [CrossRef]
- Saias, L.; Gomes, A.; Cazales, M.; Ducommun, B.; Lobjois, V. Cell-Cell Adhesion and Cytoskeleton Tension Oppose Each Other in Regulating Tumor Cell Aggregation. Cancer Res. 2015, 75, 2426–2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gava, F.; Rigal, L.; Mondesert, O.; Pesce, E.; Ducommun, B.; Lobjois, V. Gap junctions contribute to anchorage-independent clustering of breast cancer cells. BMC Cancer 2018, 18, 221. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, J.; Rigal, L.; Mondesert, O.; Morin, R.; Corsaut, G.; Vigneau, M.; Ducommun, B.; Lobjois, V. Mitotic arrest affects clustering of tumor cells. Cell Div. 2021, 16, 2. [Google Scholar] [CrossRef]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehar, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e1417. [Google Scholar] [CrossRef]
- Ghandi, M.; Huang, F.W.; Jane-Valbuena, J.; Kryukov, G.V.; Lo, C.C.; McDonald, E.R., 3rd; Barretina, J.; Gelfand, E.T.; Bielski, C.M.; Li, H.; et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 2019, 569, 503–508. [Google Scholar] [CrossRef]
- Le Cao, K.A.; Rossouw, D.; Robert-Granie, C.; Besse, P. A sparse PLS for variable selection when integrating omics data. Stat. Appl. Genet. Mol. Biol. 2008, 7, 35. [Google Scholar] [CrossRef]
- Tibshirani, R. Regression Shrinkage and Selection via the Lasso. J. R. Stat. Society Ser. B (Methodol.) 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.A. mixOmics: An R package for ′omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- R: A Language and Environment for Statistical Computing, Version 1.2.5033. R Foundation for Statistical Computing. R Core Team: Vienna, Austria, 2021. Available online: https://www.R-project.org(accessed on 1 November 2021).
- Andersson, C.R.; Selvin, T.; Blom, K.; Rubin, J.; Berglund, M.; Jarvius, M.; Lenhammar, L.; Parrow, V.; Loskog, A.; Fryknas, M.; et al. Mebendazole is unique among tubulin-active drugs in activating the MEK-ERK pathway. Sci. Rep. 2020, 10, 13124. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Sugimoto, Y.; Nakagawa, C.; Hosomi, K.; Takada, M. Integrative analysis of clinical and bioinformatics databases to identify anticancer properties of digoxin. Sci. Rep. 2019, 9, 16597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, A.T.; Weitz, M.; Breheny, P.J.; Ear, P.H.; Darbro, B.; Brown, B.J.; Braun, T.A.; Li, G.; Umesalma, S.; Kaemmer, C.A.; et al. Gene Expression Signatures Identify Novel Therapeutics for Metastatic Pancreatic Neuroendocrine Tumors. Clin. Cancer Res. 2020, 26, 2011–2021. [Google Scholar] [CrossRef]
- Goyette, M.A.; Cusseddu, R.; Elkholi, I.; Abu-Thuraia, A.; El-Hachem, N.; Haibe-Kains, B.; Gratton, J.P.; Cote, J.F. AXL knockdown gene signature reveals a drug repurposing opportunity for a class of antipsychotics to reduce growth and metastasis of triple-negative breast cancer. Oncotarget 2019, 10, 2055–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkountela, S.; Castro-Giner, F.; Szczerba, B.M.; Vetter, M.; Landin, J.; Scherrer, R.; Krol, I.; Scheidmann, M.C.; Beisel, C.; Stirnimann, C.U.; et al. Circulating Tumor Cell Clustering Shapes DNA Methylation to Enable Metastasis Seeding. Cell 2019, 176, 98–112.e114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Kumar, B.; Pan, K.; Dhawan, P.; Singh, A.B. HDAC-4 regulates claudin-2 expression in EGFR-ERK1/2 dependent manner to regulate colonic epithelial cell differentiation. Oncotarget 2017, 8, 87718–87736. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, M.; Singh, A.B.; Smith, J.J.; Sharma, A.; Chen, X.; Eschrich, S.; Yeatman, T.J.; Beauchamp, R.D.; Dhawan, P. HDAC inhibitors regulate claudin-1 expression in colon cancer cells through modulation of mRNA stability. Oncogene 2010, 29, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area-2h | AUC | Circularity | |
|---|---|---|---|
| 59M | 0.774 | 9.853 | 0.6451 |
| A549 | 0.724 | 10.88 | 0.4763 |
| BT549 | 0.786 | 10.97 | 0.4133 |
| COLO205 | 1.03 | 16.3 | 0.5596 |
| COV318 | 0.646 | 9.018 | 0.6619 |
| COV362 | 0.838 | 14.01 | 0.4843 |
| COV434 | 0.596 | 11.04 | 0.5506 |
| DLD1 | 0.85 | 14.26 | 0.6126 |
| H226 | 0.86 | 11.66 | 0.4747 |
| H23 | 0.693 | 11.26 | 0.3673 |
| H460 | 0.772 | 11.13 | 0.4898 |
| H522 | 0.644 | 11.99 | 0.4031 |
| HCT116 | 0.782 | 10.39 | 0.6901 |
| HS578T | 0.477 | 6.277 | 0.8117 |
| HT29 | 0.966 | 16.76 | 0.4627 |
| KURAMOCHI | 0.581 | 7.287 | 0.5922 |
| LNCAP | 0.779 | 12.76 | 0.6773 |
| MCF7 ATCC | 0.584 | 8.887 | 0.5947 |
| MCF7 ECACC | 0.47 | 6.93 | 0.5506 |
| MDAMB231 | 0.83 | 13.25 | 0.3204 |
| MDAMB436 | 0.675 | 11.14 | 0.4921 |
| OAW28 | 0.68 | 9.755 | 0.3956 |
| OVCAR 3 | 0.433 | 7.55 | 0.5445 |
| SW620 | 0.873 | 15.17 | 0.3807 |
| T47D | 0.854 | 11.76 | 0.8374 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gava, F.; Pignolet, J.; Déjean, S.; Mondésert, O.; Morin, R.; Agossa, J.; Ducommun, B.; Lobjois, V. Quantitative Analysis of Cell Aggregation Dynamics Identifies HDAC Inhibitors as Potential Regulators of Cancer Cell Clustering. Cancers 2021, 13, 5840. https://doi.org/10.3390/cancers13225840
Gava F, Pignolet J, Déjean S, Mondésert O, Morin R, Agossa J, Ducommun B, Lobjois V. Quantitative Analysis of Cell Aggregation Dynamics Identifies HDAC Inhibitors as Potential Regulators of Cancer Cell Clustering. Cancers. 2021; 13(22):5840. https://doi.org/10.3390/cancers13225840
Chicago/Turabian StyleGava, Fabien, Julie Pignolet, Sébastien Déjean, Odile Mondésert, Renaud Morin, Joseph Agossa, Bernard Ducommun, and Valérie Lobjois. 2021. "Quantitative Analysis of Cell Aggregation Dynamics Identifies HDAC Inhibitors as Potential Regulators of Cancer Cell Clustering" Cancers 13, no. 22: 5840. https://doi.org/10.3390/cancers13225840