
Dysregulation of Translation Factors EIF2S1, EIF5A and EIF6 in Intestinal-Type Adenocarcinoma (ITAC)

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In silico Analysis of Publicly Available Data on Adenocarcinoma
2.2. Analysis and Validation of Significant eIFs In Vitro
2.3. Ethics Committee
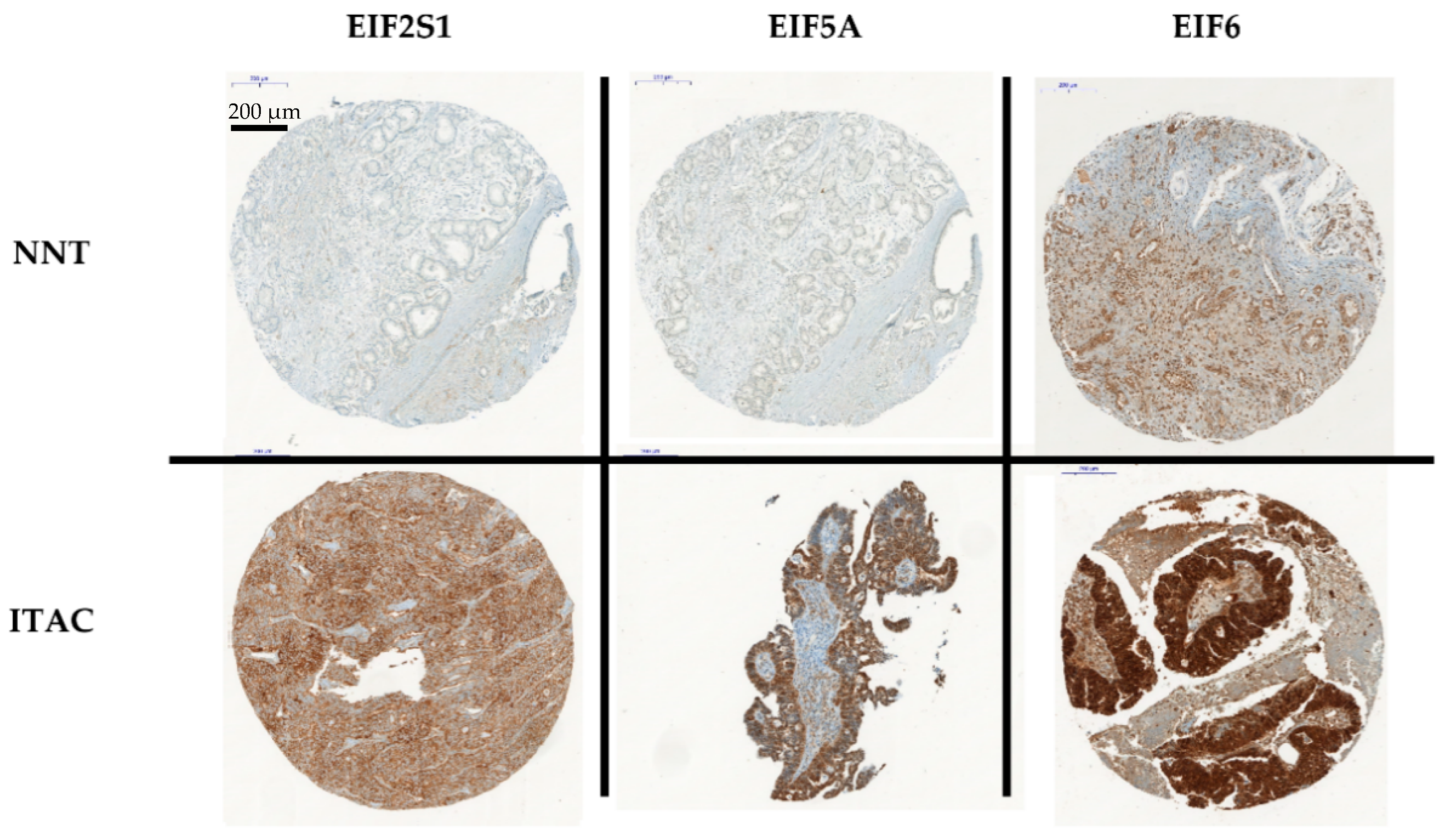
2.4. Immunohistochemistry
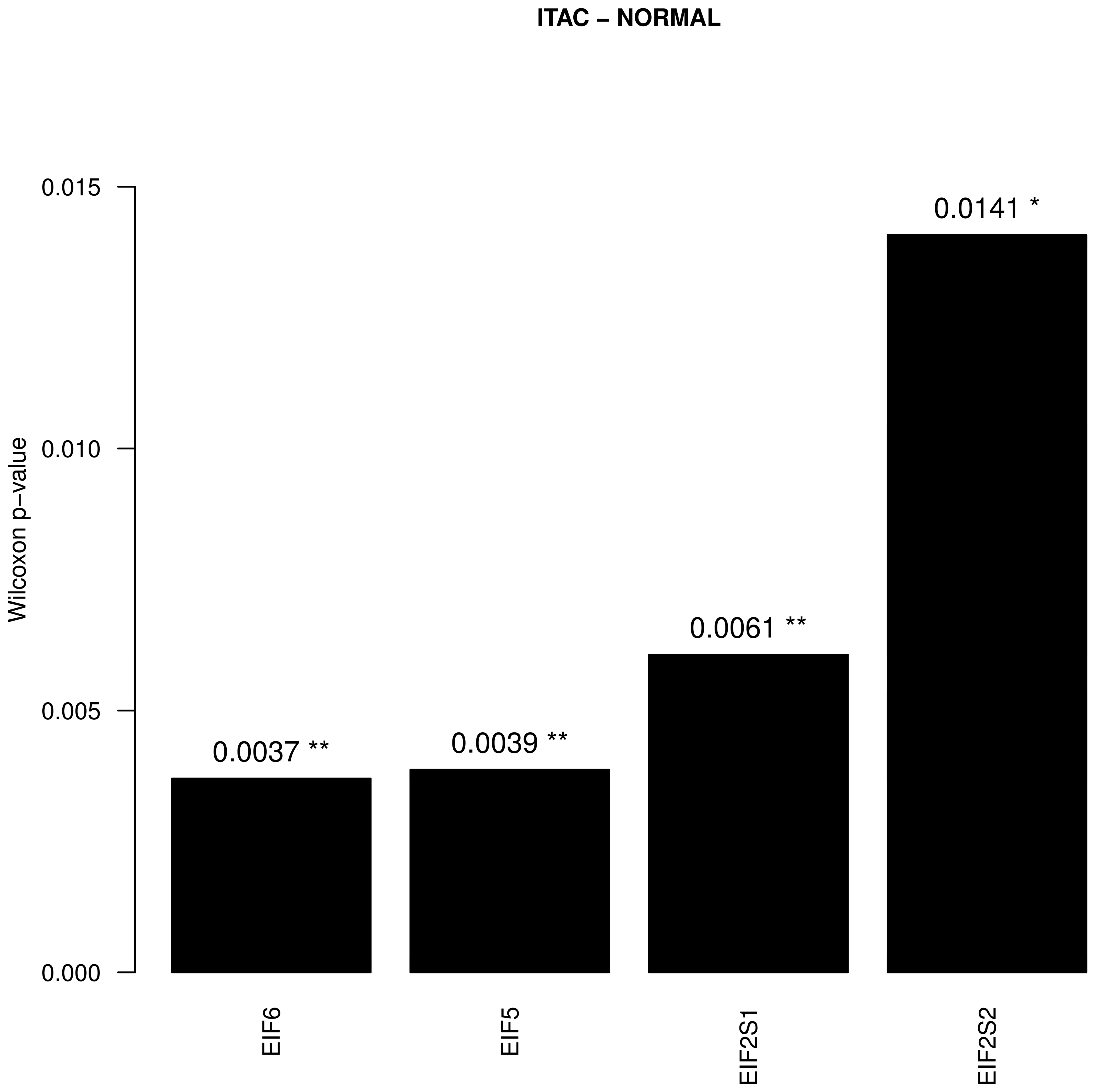
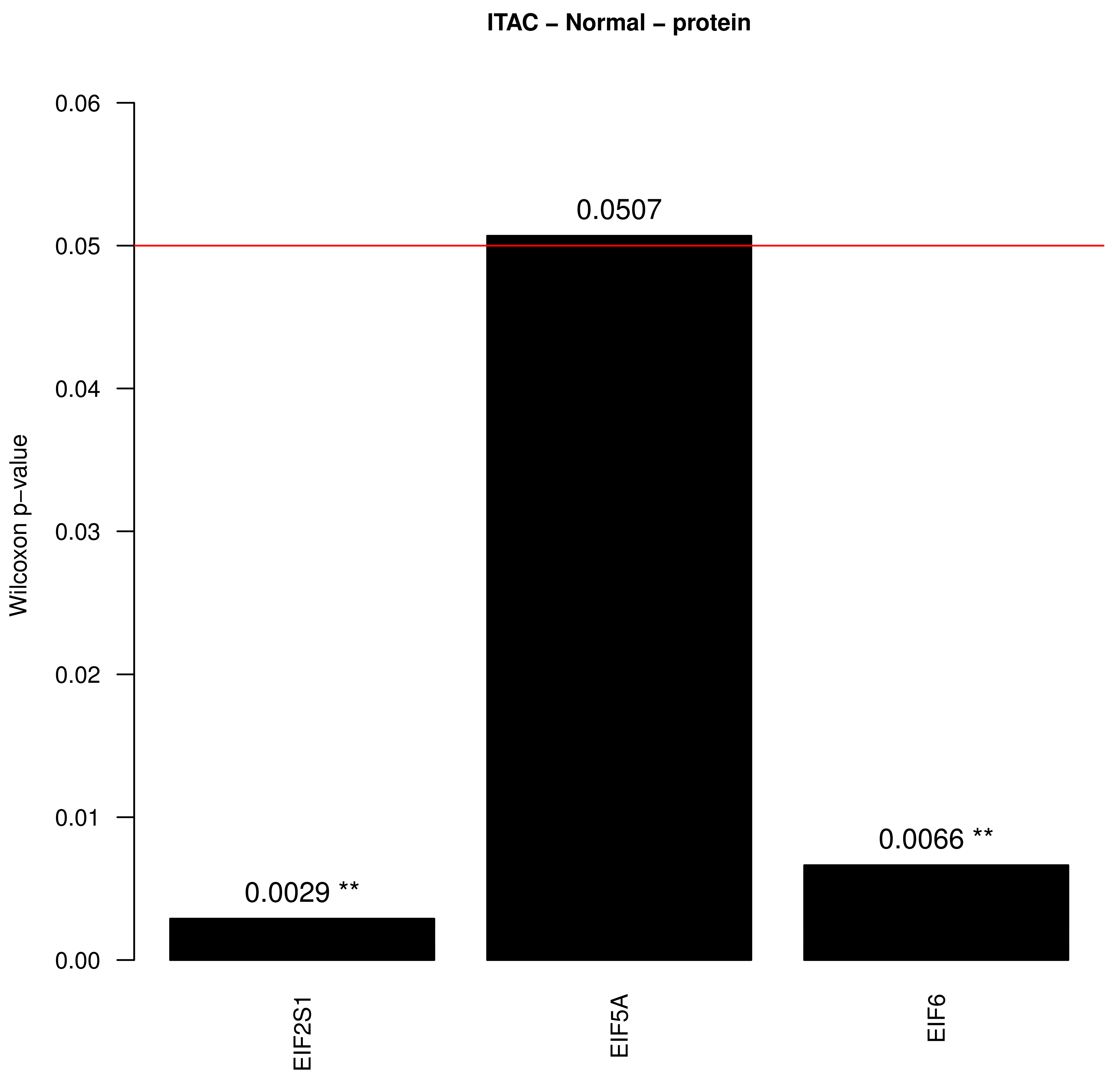
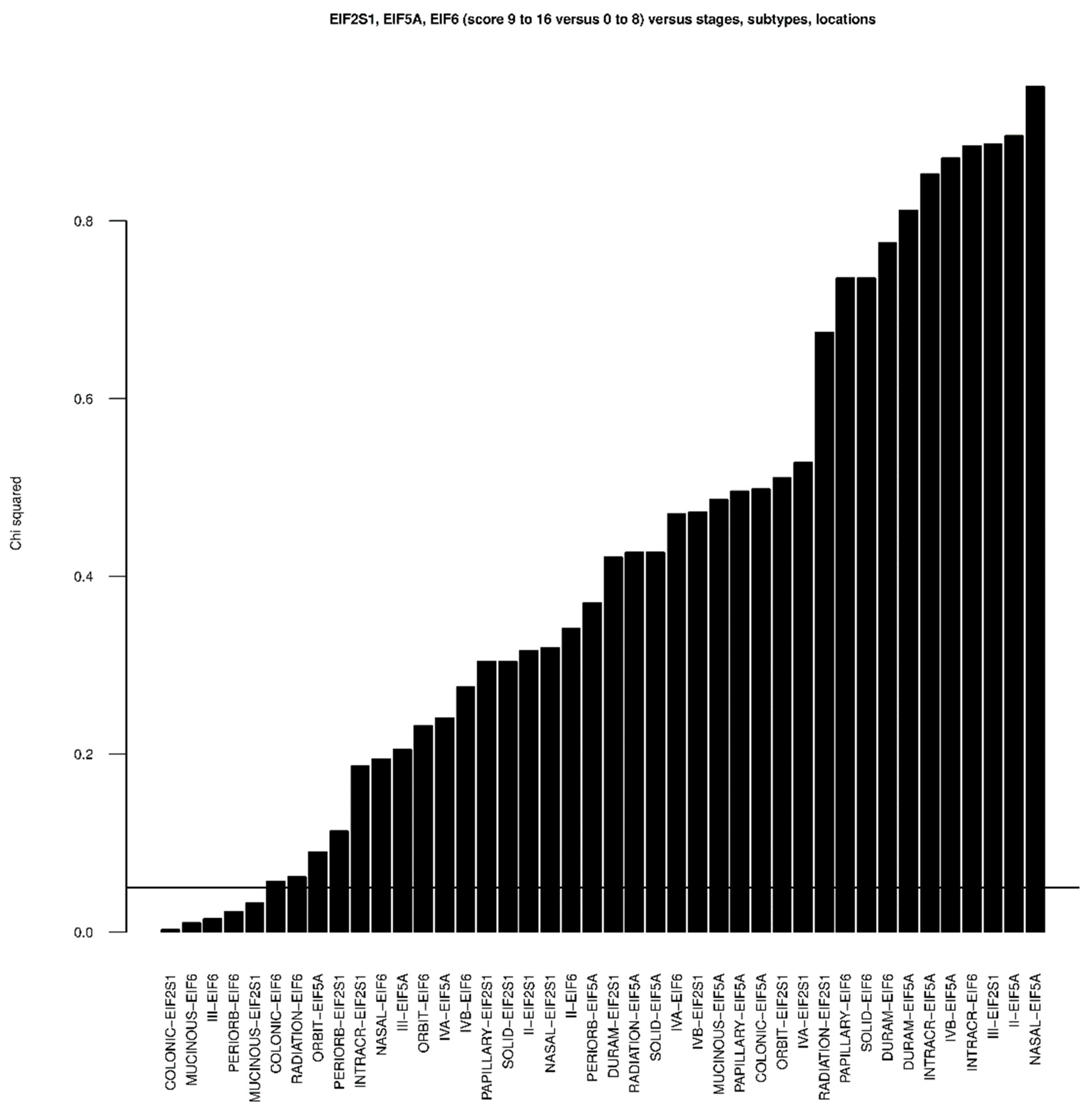
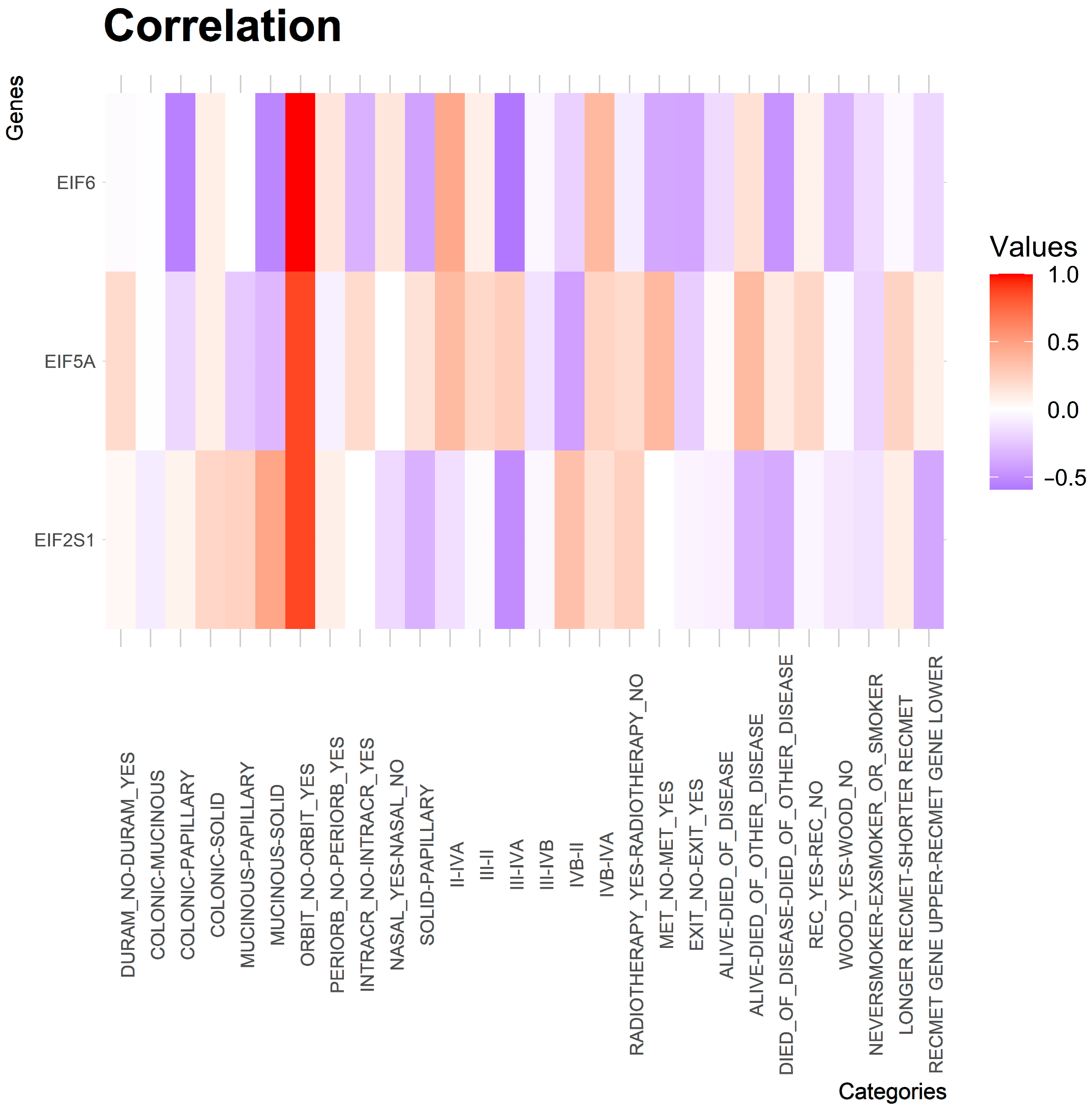
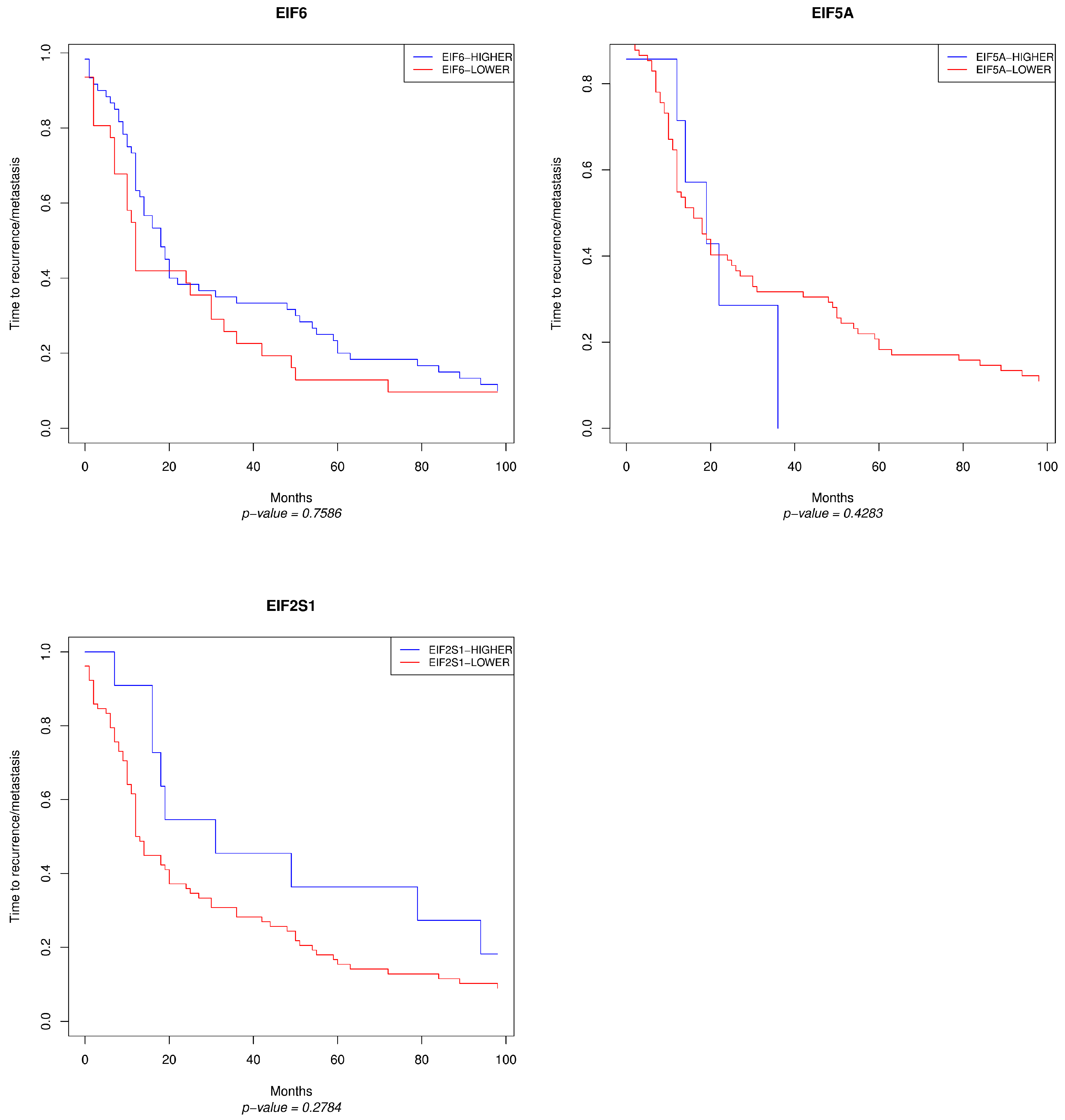
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rampinelli, V.; Ferrari, M.; Nicolai, P. Intestinal-Type Adenocarcinoma of the Sinonasal Tract: An Update. Curr. Opin. Otolaryngol. Head Neck Surg. 2018, 26, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Kazi, M.; Awan, S.; Junaid, M.; Qadeer, S.; Hassan, N.H. Management of Sinonasal Tumors: Prognostic Factors and Outcomes: A 10 Year Experience at a Tertiary Care Hospital. Indian J. Otolaryngol. Head Neck Surg. 2013, 65, 155–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivo, I. Intestinal-Type Adenocarcinoma: Classification, Immunophenotype, Molecular Features and Differential Diagnosis. Head Neck Pathol. 2017, 11, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolai, P.; Schreiber, A.; Villaret, A.B.; Lombardi, D.; Morassi, L.; Raffetti, E.; Donato, F.; Battaglia, P.; Turri–Zanoni, M.; Bignami, M.; et al. Intestinal Type Adenocarcinoma of the Ethmoid: Outcomes of a Treatment Regimen Based on Endoscopic Surgery with or without Radiotherapy. Head Neck 2016, 38, E996–E1003. [Google Scholar] [CrossRef] [PubMed]
- Bonzini, M.; Battaglia, P.; Parassoni, D.; Casa, M.; Facchinetti, N.; Turri-Zanoni, M.; Borchini, R.; Castelnuovo, P.; Ferrario, M.M. Prevalence of Occupational Hazards in Patients with Different Types of Epithelial Sinonasal Cancers. Rhinology 2013, 51, 31–36. [Google Scholar] [CrossRef]
- Fritsch, V.A.; Camp, E.R.; Lentsch, E.J. Sentinel Lymph Node Status in Merkel Cell Carcinoma of the Head and Neck: Not a Predictor of Survival. Head Neck 2014, 36, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Vergez, S.; du Mayne, M.D.; Coste, A.; Gallet, P.; Jankowski, R.; Dufour, X.; Righini, C.; Reyt, E.; Choussy, O.; Serrano, E.; et al. Multicenter Study to Assess Endoscopic Resection of 159 Sinonasal Adenocarcinomas. Ann. Surg. Oncol. 2014, 21, 1384–1390. [Google Scholar] [CrossRef] [PubMed]
- EI-Naggar, A.K.; Chan, J.K.C.; Grandis, J.R.; Takata, T.; Slootweg, P.J. WHO Classification of Head and Neck Tumours, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2017; ISBN 978-92-832-2438-9. [Google Scholar]
- Barnes, L. Intestinal-Type Adenocarcinoma of the Nasal Cavity and Paranasal Sinuses. Am. J. Surg. Pathol. 1986, 10, 192–202. [Google Scholar] [CrossRef]
- Franchi, A.; Miligi, L.; Palomba, A.; Giovannetti, L.; Santucci, M. Sinonasal Carcinomas: Recent Advances in Molecular and Phenotypic Characterization and Their Clinical Implications. Crit. Rev. Oncol. Hematol. 2011, 79, 265–277. [Google Scholar] [CrossRef]
- Kleinsasser, O.; Schroeder, H.-G. Adenocarcinomas of the Inner Nose after Exposure to Wood Dust. Eur. Arch. Oto-Rhino-Laryngol. 1988, 245, 1–15. [Google Scholar] [CrossRef]
- Cantu, G.; Solero, C.L.; Miceli, R.; Mattana, F.; Riccio, S.; Colombo, S.; Pompilio, M.; Lombardo, G.; Formillo, P.; Quattrone, P. Anterior Craniofacial Resection for Malignant Paranasal Tumors: A Monoinstitutional Experience of 366 Cases. Head Neck 2012, 34, 78–87. [Google Scholar] [CrossRef]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of Translation Initiation in Eukaryotes: Mechanisms and Biological Targets. Cell 2009, 136, 731–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doerfel, L.K.; Wohlgemuth, I.; Kothe, C.; Peske, F.; Urlaub, H.; Rodnina, M.V. EF-P Is Essential for Rapid Synthesis of Proteins Containing Consecutive Proline Residues. Science 2013, 339, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, E.; Shin, B.-S.; Woolstenhulme, C.J.; Kim, J.-R.; Saini, P.; Buskirk, A.R.; Dever, T.E. EIF5A Promotes Translation of Polyproline Motifs. Mol. Cell 2013, 51, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Horsey, E.W.; Jakovljevic, J.; Miles, T.D.; Harnpicharnchai, P.; Woolford, J.L. Role of the Yeast Rrp1 Protein in the Dynamics of Pre-Ribosome Maturation. RNA 2004, 10, 813–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, D.M.; Chaudhuri, A.; Maitra, U. Eukaryotic Ribosomal Subunit Anti-Association Activity of Calf Liver Is Contained in a Single Polypeptide Chain Protein of Mr=25,500 (Eukaryotic Initiation Factor 6). J. Biol. Chem. 1982, 257, 7712–7719. [Google Scholar] [CrossRef]
- Russell, D.W.; Spremulli, L.L. Purification and Characterization of a Ribosome Dissociation Factor (Eukaryotic Initiation Factor 6) from Wheat Germ. J. Biol. Chem. 1979, 254, 8796–8800. [Google Scholar] [CrossRef]
- Pestova, T.V.; Kolupaeva, V.G.; Lomakin, I.B.; Pilipenko, E.V.; Shatsky, I.N.; Agol, V.I.; Hellen, C.U. Molecular Mechanisms of Translation Initiation in Eukaryotes. Proc. Natl. Acad. Sci. USA 2001, 98, 7029–7036. [Google Scholar] [CrossRef] [Green Version]
- Kapp, L.D.; Lorsch, J.R. The Molecular Mechanics of Eukaryotic Translation. Annu. Rev. Biochem. 2004, 73, 657–704. [Google Scholar] [CrossRef] [PubMed]
- Weis, F.; Giudice, E.; Churcher, M.; Jin, L.; Hilcenko, C.; Wong, C.C.; Traynor, D.; Kay, R.R.; Warren, A.J. Mechanism of EIF6 Release from the Nascent 60S Ribosomal Subunit. Nat. Struct. Mol. Biol. 2015, 22, 914–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceci, M.; Gaviraghi, C.; Gorrini, C.; Sala, L.A.; Offenhäuser, N.; Marchisio, P.C.; Biffo, S. Release of EIF6 (P27BBP) from the 60S Subunit Allows 80S Ribosome Assembly. Nature 2003, 426, 579–584. [Google Scholar] [CrossRef]
- Komar, A.A.; Merrick, W.C. A Retrospective on EIF2A—and Not the Alpha Subunit of EIF2. Int. J. Mol. Sci. 2020, 21, 2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golob-Schwarzl, N.; Puchas, P.; Gogg-Kamerer, M.; Weichert, W.; Göppert, B.; Haybaeck, J. New Pancreatic Cancer Biomarkers EIF1, EIF2D, EIF3C and EIF6 Play a Major Role in Translational Control in Ductal Adenocarcinoma. Anticancer. Res. 2020, 40, 3109–3118. [Google Scholar] [CrossRef] [PubMed]
- Spilka, R.; Ernst, C.; Mehta, A.K.; Haybaeck, J. Eukaryotic Translation Initiation Factors in Cancer Development and Progression. Cancer Lett. 2013, 340, 9–21. [Google Scholar] [CrossRef]
- Golob-Schwarzl, N.; Schweiger, C.; Koller, C.; Krassnig, S.; Gogg-Kamerer, M.; Gantenbein, N.; Toeglhofer, A.M.; Wodlej, C.; Bergler, H.; Pertschy, B.; et al. Separation of Low and High Grade Colon and Rectum Carcinoma by Eukaryotic Translation Initiation Factors 1, 5 and 6. Oncotarget 2017, 8, 101224–101243. [Google Scholar] [CrossRef] [Green Version]
- Detre, S.; Saclani Jotti, G.; Dowsett, M. A “Quickscore” Method for Immunohistochemical Semiquantitation: Validation for Oestrogen Receptor in Breast Carcinomas. J. Clin. Pathol. 1995, 48, 876–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brierley, J.D.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours, 7th ed.; Wiley Blackwell: Hoboken, NJ, USA, 2017. [Google Scholar]
- Ali, M.U.; Ur Rahman, M.S.; Jia, Z.; Jiang, C. Eukaryotic Translation Initiation Factors and Cancer. Tumour Biol. 2017, 39, 1010428317709805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, B.; Sanz, R.; Aragüés, R.; Oliva, B.; Sierra, A. Functional Clustering of Metastasis Proteins Describes Plastic Adaptation Resources of Breast-Cancer Cells to New Microenvironments. J. Proteome Res. 2008, 7, 3242–3253. [Google Scholar] [CrossRef]
- Harris, M.N.; Ozpolat, B.; Abdi, F.; Gu, S.; Legler, A.; Mawuenyega, K.G.; Tirado-Gomez, M.; Lopez-Berestein, G.; Chen, X. Comparative Proteomic Analysis of All-Trans-Retinoic Acid Treatment Reveals Systematic Posttranscriptional Control Mechanisms in Acute Promyelocytic Leukemia. Blood 2004, 104, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Flavin, R.J.; Smyth, P.C.; Finn, S.P.; Laios, A.; O’Toole, S.A.; Barrett, C.; Ring, M.; Denning, K.M.; Li, J.; Aherne, S.T.; et al. Altered EIF6 and Dicer Expression Is Associated with Clinicopathological Features in Ovarian Serous Carcinoma Patients. Mod. Pathol. 2008, 21, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Biffo, S.; Sanvito, F.; Costa, S.; Preve, L.; Pignatelli, R.; Spinardi, L.; Marchisio, P.C. Isolation of a Novel Beta4 Integrin-Binding Protein (P27(BBP)) Highly Expressed in Epithelial Cells. J. Biol. Chem. 1997, 272, 30314–30321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Haybaeck, J.; Yang, Z. Therapeutic Potential of PI3K/AKT/MTOR Pathway in Gastrointestinal Stromal Tumors: Rationale and Progress. Cancers 2020, 12, 2927. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, L.; Chakraborty, A.; Robert, C.; Vagner, S. The Plasticity of MRNA Translation during Cancer Progression and Therapy Resistance. Nat. Rev. Cancer 2021, 21, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Yu, J.; Ward, R.; Liu, Y.; Hao, Q.; An, S.; Xu, T. Eukaryotic Translation Initiation Factors as Promising Targets in Cancer Therapy. Cell Commun. Signal. 2020, 18, 175. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, J.; Zhou, J.; Xiao, Z.; Ding, N.; Duan, Y.; Li, W.; Sun, L. EIF2A Promotes Cell Survival during Paclitaxel Treatment in Vitro and in Vivo. J. Cell Mol. Med. 2019, 23, 6060–6071. [Google Scholar] [CrossRef] [PubMed]
- Golob-Schwarzl, N.; Wodlej, C.; Kleinegger, F.; Gogg-Kamerer, M.; Birkl-Toeglhofer, A.M.; Petzold, J.; Aigelsreiter, A.; Thalhammer, M.; Park, Y.N.; Haybaeck, J. Eukaryotic Translation Initiation Factor 6 Overexpression Plays a Major Role in the Translational Control of Gallbladder Cancer. J. Cancer Res. Clin. Oncol. 2019, 145, 2699–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Tissues for Biochemical Analyses (n = 145) |
|---|---|
| Age (Median) | 66 years |
| Sex | |
| Male | 143 (98.62%) |
| Female | 2 (1.38%) |
| Subtype | |
| Papillary | 13 (8.97%) |
| Colonic | 88 (60.69%) |
| Solid | 10 (6.9%) |
| Mucinous | 13 (8.97%) |
| Stage | |
| Stage I | 31 (22.3%) |
| Stage II | 17 (12.33%) |
| Stage III | 48 (34.53%) |
| Stage IVA | 17 (12.33%) |
| Stage IVB | 26 (18.71%) |
| Intracranial spread | |
| No | 123 (88.49%) |
| Yes | 16 (11.51%) |
| Dural infiltration | |
| No | 114 (82.01%) |
| Yes | 25 (17.99%) |
| Orbital extension | |
| No | 135 (97.12%) |
| Yes | 4 (2.88%) |
| Periorbital extension | |
| No | 120 (86.33%) |
| Yes | 19 (13.67%) |
| Nasal cavity only | |
| No | 55 (39.86) |
| Yes | 83 (60.14%) |
| Exposure to wood dust | |
| No | 16 (11.51%) |
| Yes | 123 (88.49%) |
| Years exposure to wood dust (Median) | 35 |
| Tobacco exposure | |
| Never smoked | 61 (46.92%) |
| Formal smoker or smoker | 69 (53.08%) |
| Months of follow-up (Median) | 60 |
| Months to recurrence or metastasis (Median) | 16 |
| Adjuvant radiotherapy | |
| No | 58 (41.73%) |
| Yes | 81 (58.27%) |
| Recurrence | |
| No | 75 (53.96%) |
| Yes | 64 (46.04%) |
| Metastases | |
| No | 123 (88.49%) |
| Yes | 16 (11.51%) |
| Status | |
| Alive | 60 (43.17%) |
| Died of disease | 58 (41.73%) |
| Died of other cause | 21 (15.11%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schatz, C.; Sprung, S.; Schartinger, V.; Codina-Martínez, H.; Lechner, M.; Hermsen, M.; Haybaeck, J. Dysregulation of Translation Factors EIF2S1, EIF5A and EIF6 in Intestinal-Type Adenocarcinoma (ITAC). Cancers 2021, 13, 5649. https://doi.org/10.3390/cancers13225649
Schatz C, Sprung S, Schartinger V, Codina-Martínez H, Lechner M, Hermsen M, Haybaeck J. Dysregulation of Translation Factors EIF2S1, EIF5A and EIF6 in Intestinal-Type Adenocarcinoma (ITAC). Cancers. 2021; 13(22):5649. https://doi.org/10.3390/cancers13225649
Chicago/Turabian StyleSchatz, Christoph, Susanne Sprung, Volker Schartinger, Helena Codina-Martínez, Matt Lechner, Mario Hermsen, and Johannes Haybaeck. 2021. "Dysregulation of Translation Factors EIF2S1, EIF5A and EIF6 in Intestinal-Type Adenocarcinoma (ITAC)" Cancers 13, no. 22: 5649. https://doi.org/10.3390/cancers13225649