
Lymphoma-Associated Biomarkers Are Increased in Current Smokers in Twin Pairs Discordant for Smoking

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Cotinine and Immune-Related Biomarker Measurement
2.3. Statistical Analysis
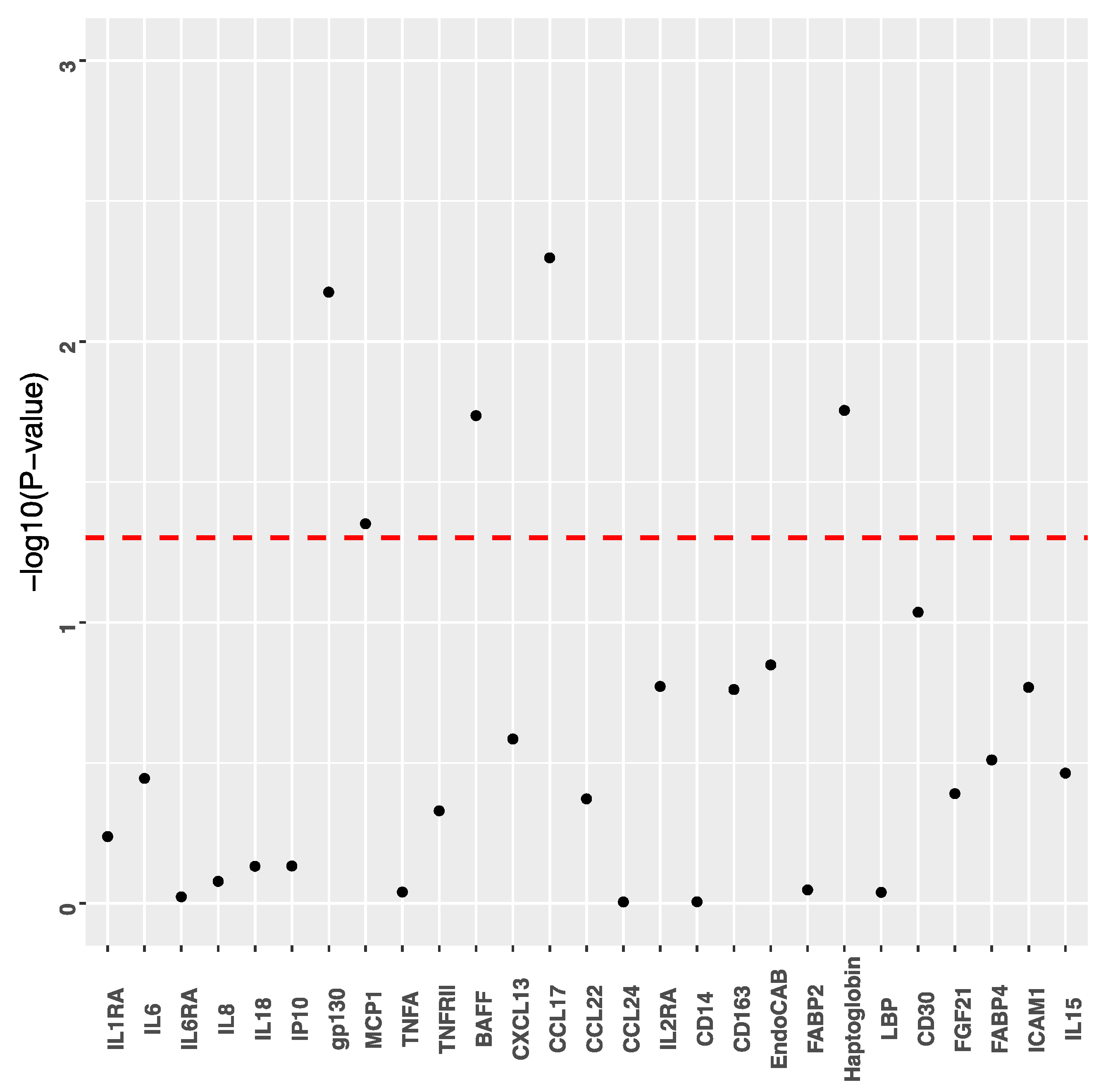
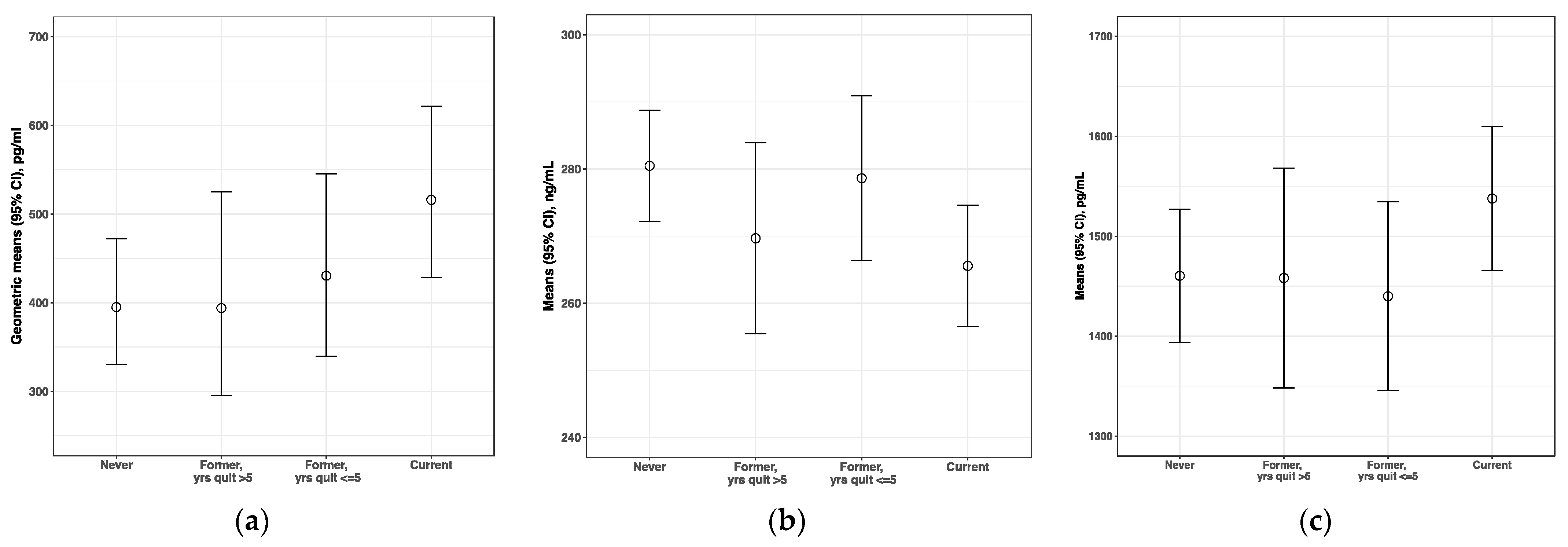
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vineis, P.; Alavanja, M.; Buffler, P.; Fontham, E.; Franceschi, S.; Gao, Y.; Gupta, P.C.; Hackshaw, A.; Matos, E.; Samet, J.; et al. Tobacco and cancer: Recent epidemiological evidence. J. Natl. Cancer Inst. 2004, 96, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kamper-Jørgensen, M.; Rostgaard, K.; Glaser, S.; Zahm, S.; Cozen, W.; Smedby, K.; Sanjosé, S.; Chang, E.; Zheng, T.; La Vecchia, C.; et al. Cigarette smoking and risk of Hodgkin lymphoma and its subtypes: A pooled analysis from the International Lymphoma Epidemiology Consortium (InterLymph). Ann. Oncol. 2013, 24, 2245–2255. [Google Scholar] [CrossRef] [PubMed]
- Cozen, W.; Hamilton, A.; Zhao, P.; Salam, M.; Deapen, D.; Nathwani, B.; Weiss, L.M.; Mack, T.M. A protective role for early childhood exposures and young adult Hodgkin lymphoma. Blood J. Am. Soc. Hematol. 2009, 114, 4014–4020. [Google Scholar]
- Morton, L.M.; Hartge, P.; Holford, T.R.; Holly, E.A.; Chiu, B.C.; Vineis, P.; Stagnaro, E.; Willett, E.V.; Franceschi, S.; La Vecchia, C.; et al. Cigarette smoking and risk of non-Hodgkin lymphoma: A pooled analysis from the International Lymphoma Epidemiology Consortium (InterLymph). Cancer Epidemiol. Biomark. Prev. 2005, 14, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Gern, J.E.; Lemanske, R.F.; Busse, W.W. Early life origins of asthma. J. Clin. Invest. 1999, 104, 837–843. [Google Scholar] [CrossRef] [Green Version]
- Wurster, A.; Rodgers, V.; White, M.; Rothstein, T.; Grusby, M. Interleukin-4-mediated protection of primary B cells from Apoptosis through Stat6-dependent Up-regulation of Bcl-xL. J. Biol. Chem. 2002, 277, 27169–27175. [Google Scholar] [CrossRef] [Green Version]
- Noakes, P.S.; Holt, P.G.; Prescott, S.L. Maternal smoking in pregnancy alters neonatal cytokine responses. Allergy 2003, 58, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Byron, K.A.; Varigos, G.A.; Wootton, A.M. IL-4 production is increased in cigarette smokers. Clin. Exp. Immunol. 1994, 95, 333–336. [Google Scholar] [CrossRef]
- Cozen, W.; Diaz-Sanchez, D.; James Gauderman, W.; Zadnick, J.; Cockburn, M.; Gill, P.; Masood, R.; Hamilton, A.S.; Jyrala, M.; Mack, T.M. Th1 and Th2 cytokines and IgE levels in identical twins with varying levels of cigarette consumption. J. Clin. Immunol. 2004, 24, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Andreoli, C.; Bassi, A.; Gregg, E.O.; Nunziata, A.; Puntoni, R.; Corsini, E. Effects of cigarette smoking on circulating leukocytes and plasma cytokines in monozygotic twins. Clin. Chem. Lab. Med. 2015, 53, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Cozen, W.; Gill, P.; Salam, M.; Nieter, A.; Masood, R.; Cockburn, M.; Gauderman, W.J.; Martinez-Maza, O.; Nathwani, B.N.; Pike, M.C.; et al. Interleukin-2, interleukin-12 and interferon-γ levels and risk of young adult Hodgkin lymphoma. Blood J. Am. Soc. Hematol. 2008, 111, 3377–3382. [Google Scholar] [CrossRef]
- Cozen, W.; Timofeeva, M.; Li, D.; Diepstra, A.; Hazelett, D.; Delahaye-Sourdeix, M.; Edlund, C.K.; Franke, L.; Rostgaard, K.; Van Den Berg, D.J.; et al. A meta-analysis of Hodgkin lymphoma reveals 19p13.3 TCF3 as a novel susceptibility locus. Nat. Comm. 2014, 25, 3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steidl, C.; Lee, T.; Shah, S.P.; Farinha, P.; Han, G.; Nayar, T.; Delaney, A.; Jones, S.J.; Iqbal, J.; Weisenburger, D.D.; et al. Tumor-associated macrophages and survival in classic Hodgkin’s lymphoma. N. Engl. J. Med. 2010, 362, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Grulich, A.E.; Vajdic, C.; Cozen, W. Altered immunity as a risk factor for non-hodgkin lymphoma. Cancer Epidemiol. Biomark. Prev. 2007, 16, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Roos, A.; Mirick, D.; Edlefsen, K.; LaCroix, A.; Kopecky, K.; Madeleine, M.; Magpantay, L.; Martinez-Maza, O. Markers of B-cell activation in relation to risk of non-Hodgkin lymphoma. Cancer Res. 2012, 72, 4733–4743. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Plütschow, A.; Jachimowicz, R.D.; Kleefisch, D.; Reiners, K.S.; Ponader, S.; Engert, A.; von Strandmann, E.P. Baseline serum TARC levels predict therapy outcome in patients with Hodgkin lymphoma. Am. J. Hematol. 2012, 88, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Van der Weyden, C.; Pileri, S.; Feldman, A.; Whisstock, J.; Prince, H. Understanding CD30 biology and therapeutic targeting: A historical perspective providing insight into future directions. Blood Cancer J. 2017, 7, e603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozen, W.; Gill, P.S.; Ingles, S.A.; Masood, R.; Martínez-Maza, O.; Cockburn, M.G.; Gauderman, W.J.; Pike, M.C.; Bernstein, L.; Nathwani, B.N.; et al. IL-6 levels and genotype are associated with risk of young adult Hodgkin lymphoma. Blood 2004, 103, 3216–3221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Liew, L.N.; Kuo, I.C.; Huang, C.H.; Goh, D.L.-M.; Chua, K.Y. The modulatory effects of lipopolysaccharide-stimulated B cells on differential T-cell polarization. Immunology 2008, 125, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Brinkhoff, A.; Zeng, Y.; Sieberichs, A.; Dolff, S.; Shilei, X.; Sun, M.; Engler, H.; Benson, S.; Korth, J.; Schedlowski, M.; et al. B-cell dynamics during experimental endotoxemia in humans. Biosci. Rep. 2019, 39, BSR20182347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epeldegui, M.; Magpantay, L.; Guo, Y.; Halec, G.; Cumberland, W.G.; Yen, P.K.; Macatangay, B.; Margolick, J.B.; Rositch, A.F.; Wolinsky, S.; et al. A prospective study of serum microbial translocation biomarkers and risk of AIDS-related non-Hodgkin lymphoma. AIDS 2018, 32, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Jungnickel, C.; Wonnenberg, B.; Karabiber, O.; Wolf, A.; Voss, M.; Wolf, L.; Honecker, A.; Kamyschnikow, A.; Herr, C.; Bals, R.; et al. Cigarette smoke-induced disruption of pulmonary barrier and bacterial translocation drive tumor-associated inflammation and growth. Am. J. Physiol. Cell. Mol. Physiol. 2015, 309, L605–L613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.J.; Salvatore, J.E.; Aaltonen, S.; Barr, P.B.; Bogl, L.H.; Byers, H.A.; Heikkilä, K.; Korhonen, T.; Latvala, A.; Palviainen, T.; et al. FinnTwin12 cohort: An updated review. Twin Res. Hum. Genet. 2019, 22, 302–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaprio, J.; Bollepalli, S.; Buchwald, J.; Iso-Markku, P.; Korhonen, T.; Kovanen, V.; Kujala, U.; Laakkonen, E.K.; Latvala, A.; Leskinen, T.; et al. The older finnish twin cohort—45 years of follow-up. Twin Res. Hum. Genet. 2019, 22, 240–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaidesoja, M.; Aaltonen, S.; Bogl, L.; Heikkilä, K.; Kaartinen, S.; Kujala, U.; Kärkkäinen, U.; Masip, G.; Mustelin, L.; Palviainen, T.; et al. FinnTwin16: A longitudinal study from age 16 of a population-based Finnish twin cohort. Twin. Res. Hum. Genet. 2019, 22, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, J.-A.; Novalen, M.; Jatlow, P.; Huestis, M.A.; Murphy, S.E.; Kaprio, J.; Kankaanpää, A.; Galanti, L.; Stefan, C.; George, T.P.; et al. Nicotine metabolite ratio (3-Hydroxycotinine/Cotinine) in plasma and urine by different analytical methods and laboratories: Implications for clinical implementation. Cancer Epidemiol. Biomark. Prev. 2015, 24, 1239–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, M.M.; Breen, E.C.; Magpantay, L.; Detels, R.; Lepone, L.; Penugonda, S.; Bream, J.; Jacobson, L.P.; Martínez-Maza, O.; Birmann, B.M. Temporal stability of serum concentrations of cytokines and soluble receptors measured across two years in low-risk HIV-seronegative men. Cancer Epidemiol. Biomark. Prev. 2013, 22, 2009–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benowitz, N.L.; Bernert, J.T.; Caraballo, R.S.; Holiday, D.B.; Wang, J. Optimal serum cotinine levels for distinguishing cigarette smokers and nonsmokers within different racial/ethnic groups in the United States between 1999 and 2004. Am. J. Epidemiol. 2008, 169, 236–248. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Maxwell, S.; Delaney, H.D.; Kelley, K. Designing Experiments and Analyzing Data: A Model Comparison Perspective, 3rd ed.; Routledge: New York, NY, USA, 2017. [Google Scholar]
- Dare, S.; Mackay, D.; Pell, J. Relationship between smoking and obesity: A cross-sectional study of 499,504 middle-aged adults in the UK general population. PLoS ONE 2015, 10, e0123579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piirtola, M.; Jelenkovic, A.; Latvala, A.; Sund, R.; Honda, C.; Inui, F.; Watanabe, M.; Tomizawa, R.; Iwatani, Y.; Ordoñana, J.R.; et al. Association of current and former smoking with body mass index: A study of smoking discordant twin pairs from 21 twin cohorts. PLoS ONE 2018, 13, e0200140. [Google Scholar] [CrossRef] [PubMed]
- Buchwald, J.; Chenoweth, M.J.; Palviainen, T.; Zhu, G.; Benner, C.; Gordon, S.; Korhonen, T.; Ripatti, S.; Madden, P.A.F.; Lehtimäki, T.; et al. Genome-wide association meta-analysis of nicotine metabolism and cigarette consumption measures in smokers of European descent. Mol. Psychiatry 2021, 26, 2212–2223. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.S.; Lessov-Schlaggar, C.N.; Cockburn, M.G.; Unger, J.B.; Cozen, W.; Mack, T.M. Gender differences in determinants of smoking initiation and persistence in California Twins. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1189–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangino, M.; Roederer, M.; Beddall, M.H.; Nestle, F.O.; Spector, T.D. Innate and adaptive immune traits are differentially affected by genetic and environmental factors. Nat. Commun. 2017, 8, 13850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Li, J.-Y.; Xu, W. Role of BAFF/BAFF-R axis in B-cell non-Hodgkin lymphoma. Crit. Rev. Oncol. 2014, 91, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Shiels, M.S.; Katki, H.A.; Freedman, N.D.; Purdue, M.; Wentzensen, N.; Trabert, B.; Kitahara, C.M.; Furr, M.; Li, Y.; Kemp, T.J.; et al. Cigarette smoking and variations in systemic immune and inflammation markers. J. Natl. Cancer Inst. 2014, 106, dju294. [Google Scholar] [CrossRef]
- Ritter, M.; Göggel, R.; Chaudhary, N.; Wiedenmann, A.; Jung, B.; Weith, A.; Seither, P. Elevated expression of TARC (CCL17) and MDC (CCL22) in models of cigarette smoke-induced pulmonary inflammation. Biochem. Biophys. Res. Commun. 2005, 334, 254–262. [Google Scholar] [CrossRef]
- Van den Berg, A.; Visser, L.; Poppema, S. High expression of the CC chemokine TARC in Reed-Sternberg cells. A possible explanation for the characteristic T-cell infiltrate in Hodgkin’s lymphoma. Am. J. Pathol. 1999, 154, 1685–1691. [Google Scholar] [CrossRef]
- Plattel, W.J.; Berg, A.V.D.; Visser, L.; Van Der Graaf, A.-M.; Pruim, J.; Vos, H.; Hepkema, B.; Diepstra, A.; Van Imhoff, G.W. Plasma thymus and activation-regulated chemokine as an early response marker in classical Hodgkin’s lymphoma. Haematologica 2011, 97, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Al-Bayaty, F.H.; Baharuddin, N.; Abdulla, M.A.; Ali, H.M.; Arkilla, M.B.; Albayaty, M.F. The influence of cigarette smoking on gingival bleeding and serum concentrations of haptoglobin and Alpha 1-Antitrypsin. BioMed Res. Int. 2013, 2013, 684154. [Google Scholar] [CrossRef] [PubMed]
- Komiyama, M.; Takanabe, R.; Ono, K.; Shimada, S.; Wada, H.; Yamakage, H.; Satoh-Asahara, N.; Morimoto, T.; Shimatsu, A.; Takahashi, Y.; et al. Association between monocyte chemoattractant protein-1 and blood pressure in smokers. J. Int. Med. Res. 2017, 46, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Morissette, M.C.; Gao, Y.; Shen, P.; Thayaparan, D.; Bérubé, J.-C.; Paré, P.D.; Brandsma, C.-A.; Hao, K.; Bossé, Y.; Ettinger, R.; et al. Role of BAFF in pulmonary autoantibody responses induced by chronic cigarette smoke exposure in mice. Physiol. Rep. 2016, 4, e13057. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Brooks, G.D.; McLeod, L.; Ruwanpura, S.; Jenkins, B. Differential involvement of gp130 signalling pathways in modulating tobacco carcinogen-induced lung tumourigenesis. Oncogene 2014, 34, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Kastelein, T.; Duffield, R.; Marino, F. Human insitu cytokine and leukocyte responses to acute smoking. J. Immunotoxicol. 2017, 14, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, J.H.; Etzerodt, A.; Svendsen, P.; Moestrup, S.K. The Haptoglobin-CD163-Heme Oxygenase-1 Pathway for Hemoglobin Scavenging. Oxidative Med. Cell. Longev. 2013, 2013, 523652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epeldegui, M.; Hussain, S. The role of microbial translocation and immune activation in AIDS-associated non-Hodgkin lymphoma pathogenesis: What have we learned? Crit. Rev. Immunol. 2020, 40, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Hjalgrim, H.; Ekström-Smedby, K.; Rostgaard, K.; Amini, R.-M.; Molin, D.; Hamilton-Dutoit, S.; Schöllkopf, C.; Chang, E.T.; Ralfkiaer, E.; Adami, H.-O.; et al. Cigarette smoking and risk of Hodgkin lymphoma: A population-based case-control study. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1561–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahola-Olli, A.V.; Würtz, P.; Havulinna, A.S.; Aalto, K.; Pitkänen, N.; Lehtimäki, T.; Kähönen, M.; Lyytikäinen, L.-P.; Raitoharju, E.; Seppälä, I.; et al. Genome-wide association study identifies 27 loci influencing concentrations of circulating cytokines and growth factors. Am. J. Hum. Genet. 2016, 100, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley-Hedgepeth, A.; Lloyd-Jones, D.M.; Colvin, A.; Matthews, K.A.; Johnston, J.; Sowers, M.R.; Sternfeld, B.; Pasternak, R.C.; Chae, C.U. Ethnic differences in C-reactive protein concentrations. Clin. Chem. 2008, 54, 1027–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Hertzen, L.; Surcel, H.; Kaprio, J.; Koskenvuo, M.; Bloigu, A.; Leinonen, M.; Saikku, P. Immune responses to Chlamydia pneumoniae in twins in relation to gender and smoking. J. Med. Microbiol. 1998, 47, 441–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Characteristics | Measure (SD or %) |
|---|---|
| 67 Twin Pairs | |
| Mean Age (SD) | 37.3 (18.9) |
| Female, N (%) | 76 (56.7) |
| 134 Individual Twins | |
| BMI kg/m2, mean (SD) | 25.0 (4.7) |
| Serum cotinine ng/mL, N (%) | |
| ≤3.08 (Non-current smoking) | 91 (67.9) |
| >3.08 (Current smoking) | |
| Low | 22 (15.7) |
| High | 21 (16.4) |
| Smoking status 1, N (%) | |
| Never | 55 (41.0) |
| Former | 36 (26.9) |
| Current | 43 (32.1) |
| Years since quit among former smokers, N (%) | |
| ≤5 | 20 (55.6) |
| >5 | 16 (44.4) |
| Smoking Level Comparison 1 | Difference in Least Squares Means 2 | % Difference | p | Padj3 |
|---|---|---|---|---|
| BAFF, pg/mL | ||||
| Current lighter smokers–Noncurrent | 48.2 | 3.3 | 0.27 | 0.51 |
| Current heavier smokers–Current lighter smokers | 80.7 | 5.4 | 0.17 | 0.35 |
| Current heavier smokers–Noncurrent | 128.9 | 8.9 | 0.01 | 0.02 |
| CCL17, pg/mL | ||||
| Current lighter smokers–Noncurrent | 27.4 | 6.9 | 0.51 | 0.78 |
| Current heavier smokers–Current lighter smokers | 223.3 | 52.4 | 0.002 | 0.006 |
| Current heavier smokers–Noncurrent | 250.6 | 62.9 | <0.0001 | 0.0001 |
| Haptoglobin, ng/mL | ||||
| Current lighter smokers–Noncurrent | 90.3 | 12.5 | 0.22 | 0.43 |
| Current heavier smokers–Current lighter smokers | 119.5 | 14.7 | 0.28 | 0.53 |
| Current heavier smokers–Noncurrent | 209.8 | 29.0 | 0.02 | 0.04 |
| sgp130, ng/mL | ||||
| Current lighter smokers–Noncurrent | −10.0 | −3.6 | 0.11 | 0.24 |
| Current heavier smokers–Current lighter smokers | −7.5 | −2.8 | 0.35 | 0.62 |
| Current heavier smokers–Noncurrent | −17.5 | −6.3 | 0.01 | 0.03 |
| MCP1, pg/mL | ||||
| Current lighter smokers–Noncurrent | 30.4 | 8.4 | 0.35 | 0.61 |
| Current heavier smokers–Current lighter smokers | 50.1 | 12.8 | 0.28 | 0.52 |
| Current heavier smokers–Noncurrent | 80.5 | 22.3 | 0.03 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Conti, D.V.; Epeldegui, M.; Ollikainen, M.; Tyndale, R.F.; Hwang, A.E.; Magpantay, L.; Mack, T.M.; Martinez-Maza, O.; Kaprio, J.; et al. Lymphoma-Associated Biomarkers Are Increased in Current Smokers in Twin Pairs Discordant for Smoking. Cancers 2021, 13, 5395. https://doi.org/10.3390/cancers13215395
Wang J, Conti DV, Epeldegui M, Ollikainen M, Tyndale RF, Hwang AE, Magpantay L, Mack TM, Martinez-Maza O, Kaprio J, et al. Lymphoma-Associated Biomarkers Are Increased in Current Smokers in Twin Pairs Discordant for Smoking. Cancers. 2021; 13(21):5395. https://doi.org/10.3390/cancers13215395
Chicago/Turabian StyleWang, Jun, David V. Conti, Marta Epeldegui, Miina Ollikainen, Rachel F. Tyndale, Amie Eunah Hwang, Larry Magpantay, Thomas McCulloch Mack, Otoniel Martinez-Maza, Jaakko Kaprio, and et al. 2021. "Lymphoma-Associated Biomarkers Are Increased in Current Smokers in Twin Pairs Discordant for Smoking" Cancers 13, no. 21: 5395. https://doi.org/10.3390/cancers13215395