
Surfaceome Profiling of Rhabdomyosarcoma Reveals B7-H3 as a Mediator of Immune Evasion

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Reagents
2.2. Cell surface Biotinylation and Mass Spectrometry
2.3. Bioinformatic Annotation of Cell Surface Proteins
2.4. Expression Analysis of RMS-Enriched Cell Surface Proteins in RMS Tumors and Normal Tissue
2.5. Deconvolution Analysis of Bulk RNA-seq
2.6. T-Cell Cytotoxicity Assay
2.7. Statistical Analysis
3. Results
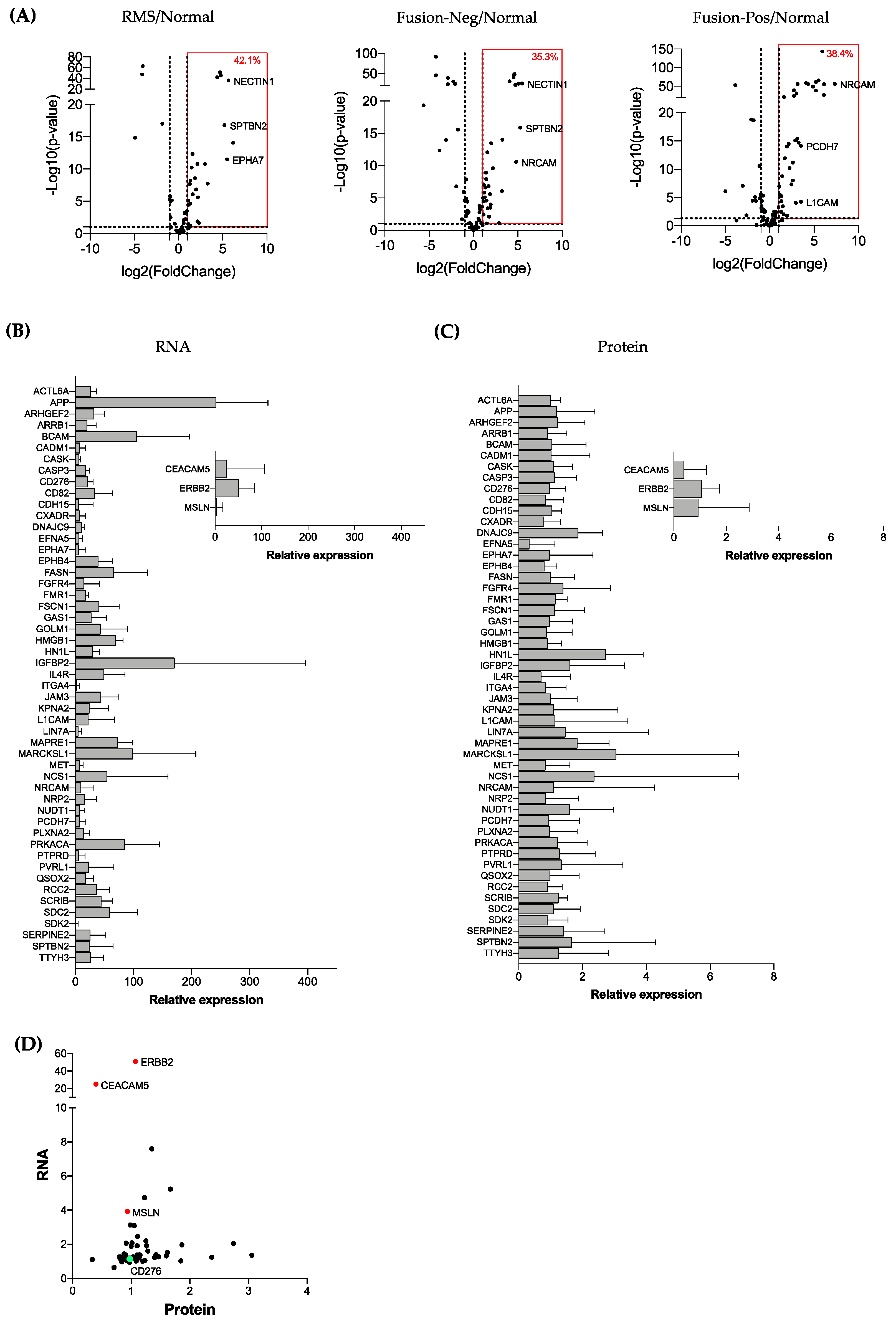
3.1. Identification of Cell Surface Protein Repertoires in RMS and Normal Muscle
3.2. Validation of RMS-Enriched Cell Surface Proteins by Combined Proteomic and Transcriptomic Analysis
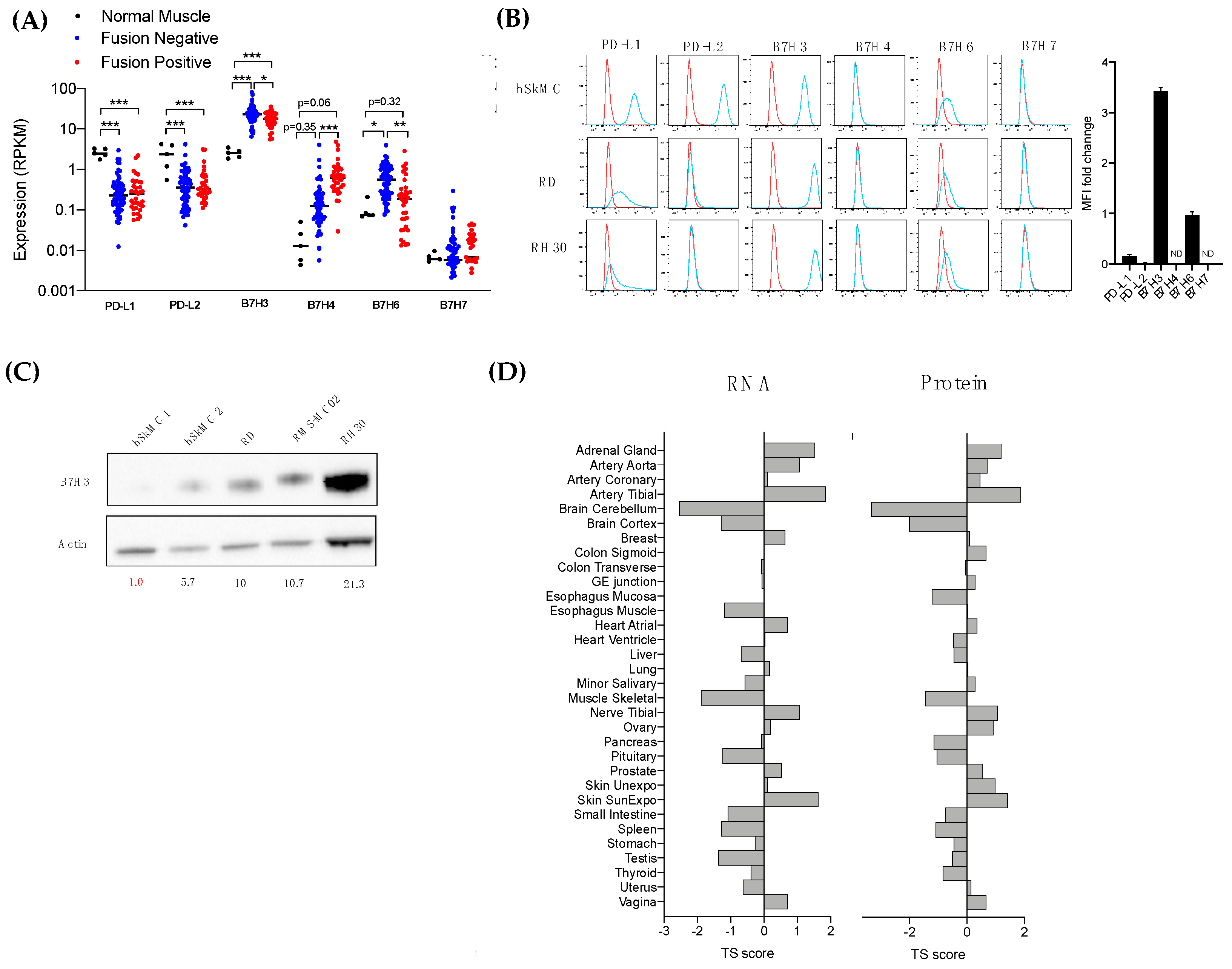
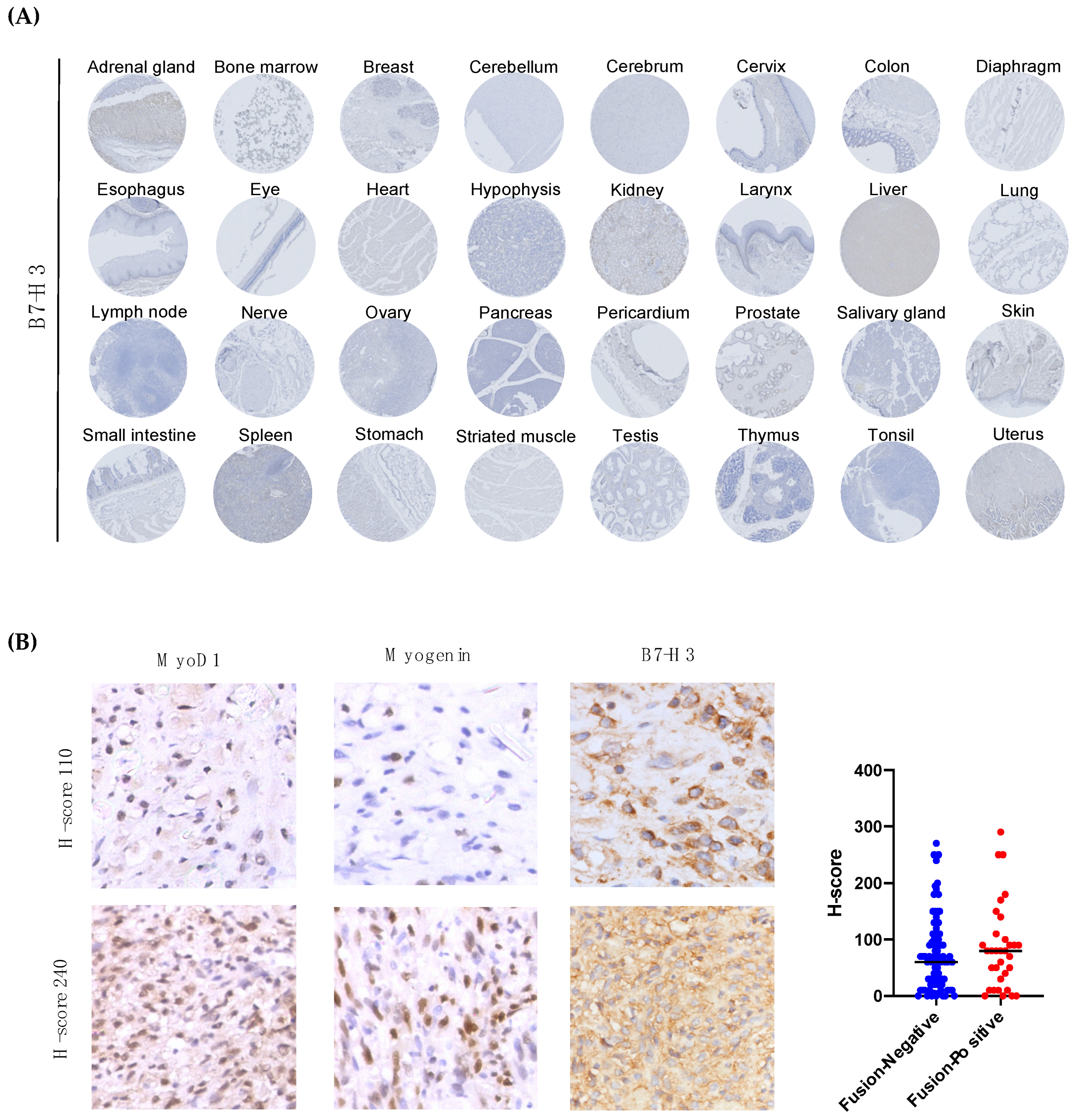
3.3. The Immune Checkpoint Molecule B7-H3 Is Upregulated in Both Fusion-Negative and Fusion-Positive RMS
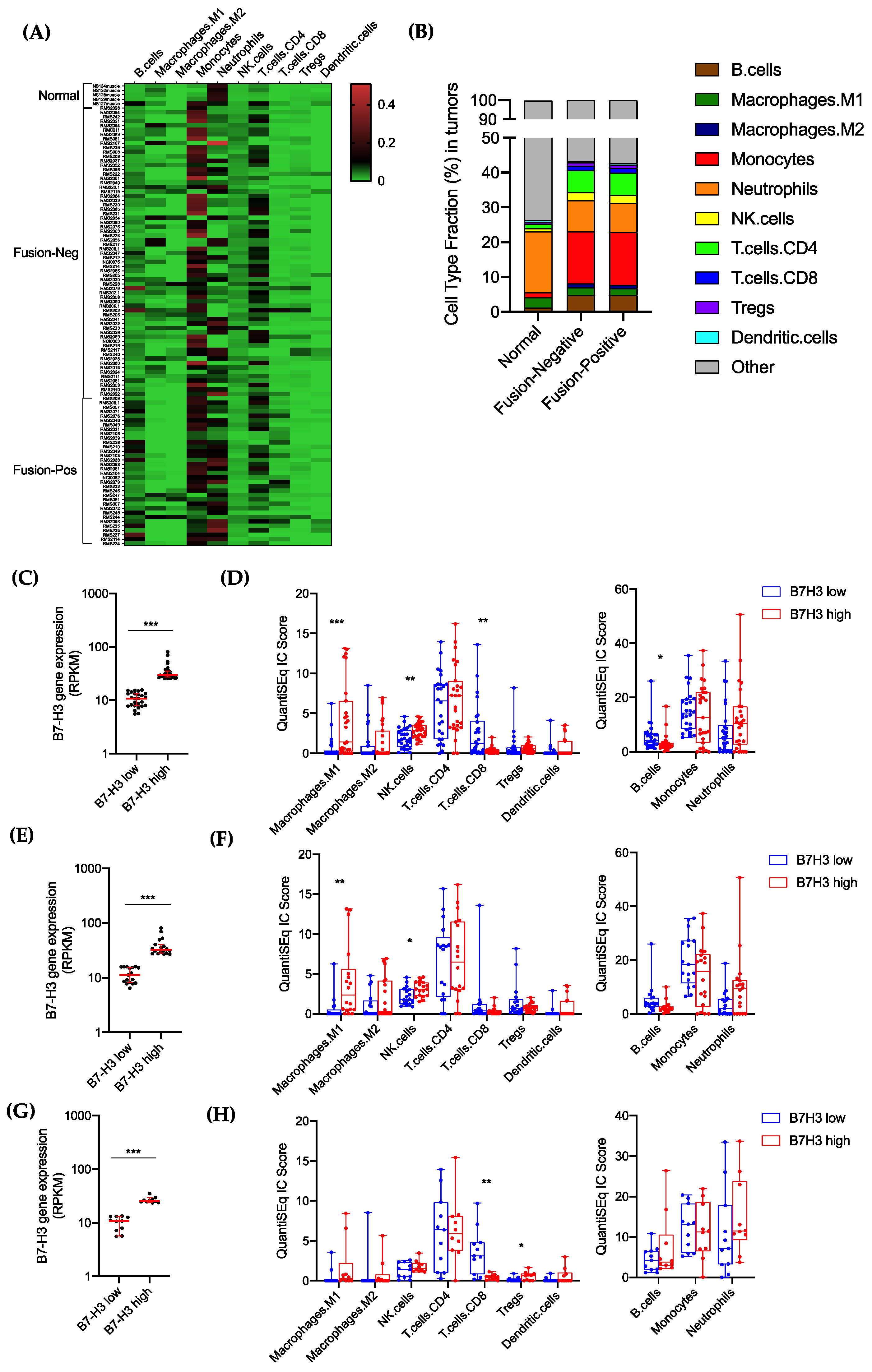
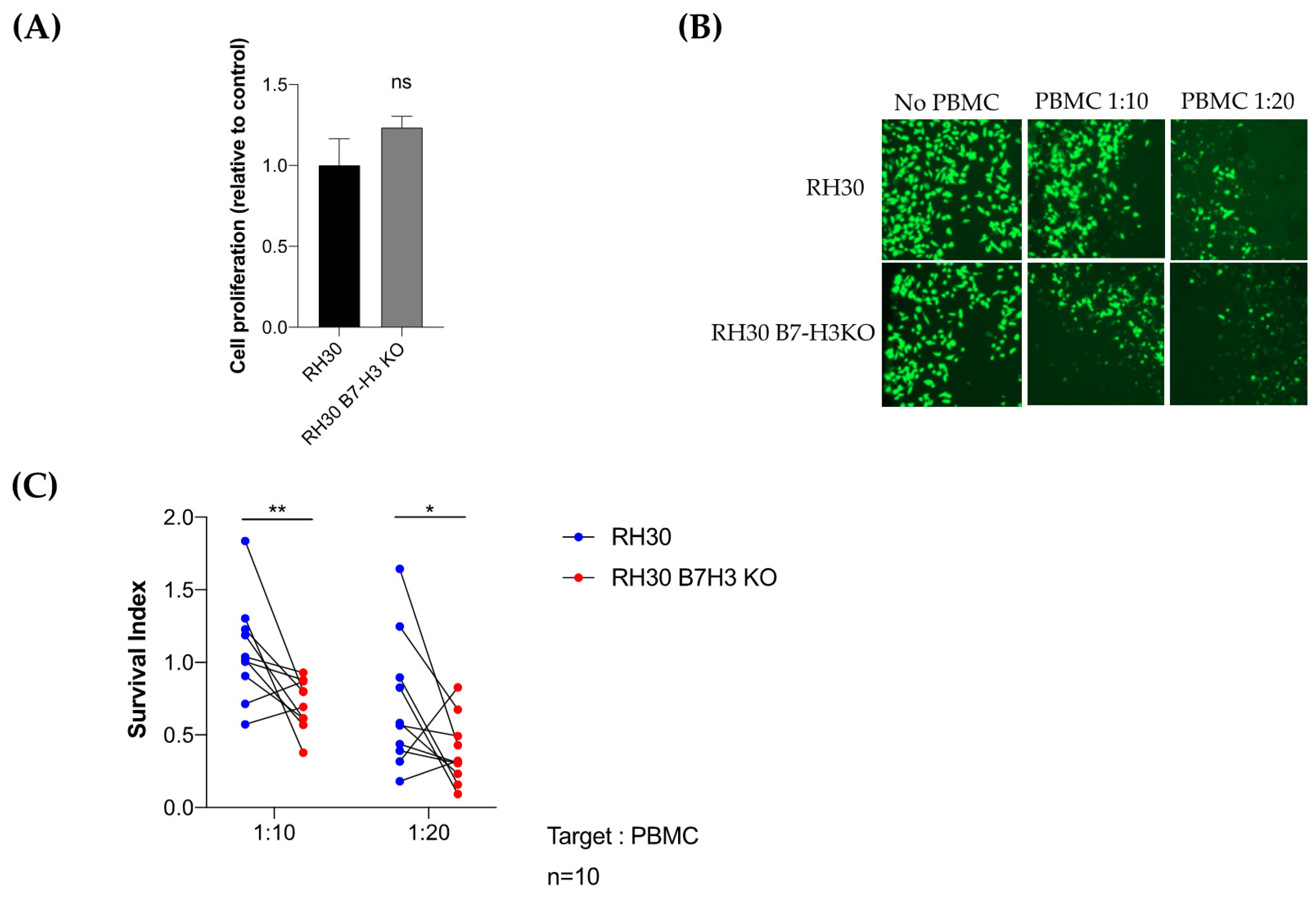
3.4. Tumor B7-H3 Overexpression Is Associated with Low Infiltration of CD8+-T Cells in RMS Tumors and Impaired Antitumor Immune Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, R.J.; D’Cruz, C.M.; Lovell, A.M.; Biegel, A.J.; Barr, F.G. Fusion of PAX7 to FKHR by the Variant t(1;13)(p36;q14) Translocation in Alveolar Rhabdomyosarcoma. Cancer Res. 1994, 54, 2869–2872. [Google Scholar]
- Barr, F.G.; Galili, N.; Holick, J.; Biegel, J.A.; Rovera, G.; Emanuel, B.S. Rearrangement of the PAX3 Paired Box Gene in the Paediatric Solid Tumour Alveolar Rhabdomyosarcoma. Nat. Genet. 1993, 3, 113–117. [Google Scholar] [CrossRef]
- Skapek, S.X.; Anderson, J.; Barr, F.G.; Bridge, J.A.; Gastier-Foster, J.M.; Parham, D.M.; Rudzinski, E.R.; Triche, T.; Hawkins, D.S. PAX-FOXO1 Fusion Status Drives Unfavorable Outcome for Children with Rhabdomyosarcoma: A Children’s Oncology Group Report. Pediatr. Blood Cancer 2013, 60, 1411–1417. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Yu, Y.; Bilke, S.; Walker, R.L.; Mayeenuddin, L.H.; Azorsa, D.O.; Yang, F.; Pineda, M.; Helman, L.J.; Meltzer, P.S. Genome-Wide Identification of PAX3-FKHR Binding Sites in Rhabdomyosarcoma Reveals Candidate Target Genes Important for Development and Cancer. Cancer Res. 2010, 70, 6497–6508. [Google Scholar] [CrossRef] [Green Version]
- Shern, J.F.; Chen, L.; Badgett, T.; Getz, G.; Chmielecki, J.; Mora, J.; Anderson, J.R.; Skapek, S.X.; Barr, F.G.; Meyerson, M.; et al. Comprehensive Genomic Analysis of Rhabdomyosarcoma Reveals a Landscape of Alterations Affecting a Common Genetic Axis in Fusion-Positive and Fusion-Negative Tumors. Cancer Discov. 2014, 4, 216–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crist, W.; Gehan, A.E.; Ragab, A.H.; Dickman, P.S.; Donaldson, S.S.; Fryer, C.; Hammond, D.; Hays, D.M.; Herrmann, J.; Heyn, R. The Third Intergroup Rhabdomyosarcoma Study. J. Clin. Oncol. 1995, 13, 610–630. [Google Scholar] [CrossRef]
- Maurer, H.M.; Beltangady, M.; Gehan, E.A.; Crist, W.; Hammond, D.; Hays, D.M.; Heyn, R.; Lawrence, W.; Newton, W.; Ortega, J.; et al. The Intergroup Rhabdomyosarcoma Study-I. A Final Report. Cancer 1988, 61, 209–220. [Google Scholar] [CrossRef]
- Maurer, H.M.; Gehan, E.A.; Beltangady, M.; Crist, W.; Dickman, P.S.; Donaldson, S.S.; Fryer, C.; Hammond, D.; Hays, D.M.; Herrmann, J.; et al. The Intergroup Rhabdomyosarcoma Study-II. Cancer 1993, 71, 1904–1922. [Google Scholar] [CrossRef]
- Punyko, J.A.; Mertens, A.C.; Gurney, J.G.; Yasui, Y.; Donaldson, S.S.; Rodeberg, D.A.; Raney, R.B.; Stovall, M.; Sklar, C.A.; Robison, L.L.; et al. Long-Term Medical Effects of Childhood and Adolescent Rhabdomyosarcoma: A Report from the Childhood Cancer Survivor Study. Pediatr. Blood Cancer 2005, 44, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Reulen, R.C.; Winter, D.L.; Frobisher, C.; Lancashire, E.R.; Stiller, C.A.; Jenney, M.E.; Skinner, R.; Stevens, M.C.; Hawkins, M.M.; British Childhood Cancer Survivor Study Steering Group. Long-term Cause-Specific Mortality among Survivors of Childhood Cancer. JAMA 2010, 304, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.-M.; Hwang, Y.-C.; Liu, I.-J.; Lee, C.-C.; Tsai, H.-Z.; Li, H.-J.; Wu, H.-C. Development of Therapeutic Antibodies for the Treatment of Diseases. J. Biomed. Sci. 2020, 27, 1–30. [Google Scholar] [CrossRef]
- Leung, K.K.; Wilson, G.M.; Kirkemo, L.L.; Riley, N.M.; Coon, J.J.; Wells, J.A. Broad and Thematic Remodeling of the Surfaceome and Glycoproteome on Isogenic Cells Transformed with Driving Proliferative Oncogenes. Proc. Natl. Acad. Sci. USA 2020, 117, 7764–7775. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Mou, K.Y.; Solomon, P.; Aggarwal, R.; Leung, K.K.; Wells, J.A. Large Remodeling of the Myc-Induced Cell Surface Proteome in B Cells and Prostate Cells Creates New Opportunities for Immunotherapy. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Martinko, A.J.; Truillet, C.; Julien, O.; Diaz, E.J.; Horlbeck, A.M.; Whiteley, G.; Blonder, J.; Weissman, J.S.; Bandyopadhyay, S.; Evans, M.J.; et al. Targeting RAS-Driven Human Cancer Cells with Antibodies to Upregulated and Essential Cell-Surface Proteins. eLife 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A Statistical Model for Identifying Proteins by Tandem Mass Spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Deng, N.; Li, Z.; Pan, C.; Duan, H. freeQuant: A Mass Spectrometry Label-Free Quantification Software Tool for Complex Proteome Analysis. Sci. World J. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lowerison, M.; Huang, C.; Lucien, F.; Chen, S.; Song, P. Ultrasound Localization Microscopy of Renal Tumor Xenografts in Chicken Embryo is Correlated to Hypoxia. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Wang, M.; Lin, S.; Jian, R.; Li, X.; Chan, J.; Dong, G.; Fang, H.; Robinson, A.E.; Snyder, M.P.; et al. A Quantitative Proteome Map of the Human Body. Cell 2020, 183, 269–283.e19. [Google Scholar] [CrossRef]
- Bonifant, C.; Jackson, H.J.; Brentjens, R.J.; Curran, K.J. Toxicity and Management in CAR T-Cell Therapy. Mol. Ther.-Oncol. 2016, 3, 16011. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Hallström, B.M.; Lindskog, C.; Mardinoglu, A.; Pontén, F.; Nielsen, J. Transcriptomics Resources of Human Tissues and Organs. Mol. Syst. Biol. 2016, 12, 862. [Google Scholar] [CrossRef]
- Mackay, M.; Afshinnekoo, E.; Rub, J.; Hassan, C.; Khunte, M.; Baskaran, N.; Owens, B.; Liu, L.; Roboz, G.J.; Guzman, M.L.; et al. The Therapeutic Landscape for Cells Engineered with Chimeric Antigen Receptors. Nat. Biotechnol. 2020, 38, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Hegde, M.; Joseph, S.K.; Pashankar, F.; DeRenzo, C.; Sanber, K.; Navai, S.; Byrd, T.T.; Hicks, J.; Xu, M.L.; Gerken, C.; et al. Tumor Response and Endogenous Immune Reactivity after Administration of HER2 CAR T Cells in a Child with Metastatic Rhabdomyosarcoma. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Dong, C. New B7 Family Checkpoints in Human Cancers. Mol. Cancer Ther. 2017, 16, 1203–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontos, F.; Michelakos, T.; Kurokawa, T.; Sadagopan, A.; Schwab, J.H.; Ferrone, C.R.; Ferrone, S. B7-H3: An Attractive Target for Antibody-based Immunotherapy. Clin. Cancer Res. 2020, 27, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A Costimulatory Molecule for T Cell Activation and IFN-γ Production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Martin-Orozco, N.; Zheng, P.; Li, J.; Zhang, P.; Tan, H.; Park, H.J.; Jeong, M.; Chang, S.H.; Kim, B.-S.; et al. Inhibition of the B7-H3 Immune Checkpoint Limits Tumor Growth by Enhancing Cytotoxic Lymphocyte Function. Cell Res. 2017, 27, 1034–1045. [Google Scholar] [CrossRef]
- Finotello, F.; Mayer, C.; Plattner, C.; Laschober, G.; Rieder, D.; Hackl, H.; Krogsdam, A.; Loncova, Z.; Posch, W.; Wilflingseder, D.; et al. Molecular and Pharmacological Modulators of the Tumor Immune Contexture Revealed by Deconvolution of RNA-seq Data. Genome Med. 2019, 11, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Wollscheid, B.; Bausch-Fluck, D.; Henderson, C.; O’Brien, R.; Bibel, M.; Schiess, R.; Aebersold, R.; Watts, J.D. Mass-Spectrometric Identification and Relative Quantification of N-Linked Cell Surface Glycoproteins. Nat. Biotechnol. 2009, 27, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Nix, M.A.; Mandal, K.; Geng, H.; Paranjape, N.; Lin, Y.-H.T.; Rivera, J.M.; Marcoulis, M.; White, K.L.; Whitman, J.D.; Bapat, S.P.; et al. Surface Proteomics Reveals CD72 as a Target for In Vitro–Evolved Nanobody-Based CAR-T Cells in KMT2A/MLL1-Rearranged B-ALL. Cancer Discov. 2021. [Google Scholar] [CrossRef]
- Heider, M.; Eichner, R.; Stroh, J.; Morath, V.; Kuisl, A.; Zecha, J.; Lawatscheck, J.; Baek, K.; Garz, A.-K.; Rudelius, M.; et al. The IMiD Target CRBN Determines HSP90 Activity Toward Transmembrane Proteins Essential in Multiple Myeloma. Mol. Cell 2021, 81, 1170–1186.e10. [Google Scholar] [CrossRef]
- Lee, J.; Bangayan, N.J.; Chai, T.; Smith, B.A.; Pariva, T.E.; Yun, S.; Vashisht, A.; Zhang, Q.; Park, J.W.; Corey, E.; et al. Systemic Surfaceome Profiling Identifies Target Antigens for Immune-Based Therapy in Subtypes of Advanced Prostate Cancer. Proc. Natl. Acad. Sci. USA 2018, 115, E4473–E4482. [Google Scholar] [CrossRef] [Green Version]
- Davicioni, E.; Finckenstein, F.G.; Shahbazian, V.; Buckley, J.D.; Triche, T.J.; Anderson, M.J. Identification of a PAX-FKHR Gene Expression Signature that Defines Molecular Classes and Determines the Prognosis of Alveolar Rhabdomyosarcomas. Cancer Res. 2006, 66, 6936–6946. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Lu, J.; Zou, C.; Shao, Y.; Chen, Y.; Narala, S.; Fang, H.; Xu, H.; Wang, J.; Shen, J.; et al. CDH4 is a Novel Determinant of Osteosarcoma Tumorigenesis and Metastasis. Oncogene 2018, 37, 3617–3630. [Google Scholar] [CrossRef] [PubMed]
- Ceresa, D.; Alessandrini, F.; Bosio, L.; Marubbi, D.; Reverberi, D.; Malatesta, P.; Appolloni, I. Cdh4 Down-Regulation Impairs in Vivo Infiltration and Malignancy in Patients Derived Glioblastoma Cells. Int. J. Mol. Sci. 2019, 20, 4028. [Google Scholar] [CrossRef] [Green Version]
- Kucharczak, J.; Charrasse, S.; Comunale, F.; Zappulla, J.; Robert, B.; Teulon-Navarro, I.; Pèlegrin, A.; Gauthier-Rouviere, C. R-Cadherin Expression Inhibits Myogenesis and Induces Myoblast Transformation via Rac1 GTPase. Cancer Res. 2008, 68, 6559–6568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, A.W.; Bartlett, P.F.; Lackmann, M. Therapeutic Targeting of EPH Receptors and Their Ligands. Nat. Rev. Drug Discov. 2013, 13, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Klein, R. Eph/Ephrin Signalling during Development. Development 2012, 139, 4105–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B. Cancer Cells Exploit the Eph-Ephrin System to Promote Invasion and Metastasis: Tales of Unwitting Partners. Sci. Signal. 2011, 4, pe28. [Google Scholar] [CrossRef] [Green Version]
- Giaginis, C.; Tsoukalas, N.; Bournakis, E.; Alexandrou, P.; Kavantzas, N.; Patsouris, E.; Theocharis, S. Ephrin (Eph) receptor A1, A4, A5 and A7 Expression in Human Non-Small Cell Lung Carcinoma: Associations with Clinicopathological Parameters, Tumor Proliferative Capacity and Patients’ Survival. BMC Clin. Pathol. 2014, 14, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Jin, M.; Sun, Y.; Jiang, A.; Wu, Y.; Li, C.; Yan, H.; Jin, H. Knockdown of Ephrin Receptor A7 Suppresses the Proliferation and Metastasis of A549 Human Lung Cancer Cells. Mol. Med. Rep. 2016, 13, 3190–3196. [Google Scholar] [CrossRef] [Green Version]
- Di, W.; Weinan, X.; Xin, L.; Zhiwei, Y.; Xinyue, G.; Jinxue, T.; Mingqi, L. Long Noncoding RNA SNHG14 Facilitates Colorectal Cancer Metastasis through Targeting EZH2-Regulated EPHA7. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrych, A.; Pęksa, R.; Kunc, M.; Krawczyk, M.; Izycka-Swieszewska, E.; Biernat, W.; Bień, E. The PD-L1/PD-1 Axis Expression on Tumor-Infiltrating Immune Cells and Tumor Cells in Pediatric Rhabdomyosarcoma. Pathol. Res. Pract. 2019, 215, 152700. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, G.; Bergamaschi, L.; Ferrari, A.; Renne, S.L.; Collini, P.; Gardelli, C.; Barisella, M.; Centonze, G.; Chiaravalli, S.; Paolino, C.; et al. PD-L1 Assessment in Pediatric Rhabdomyosarcoma: A Pilot Study. BMC Cancer 2018, 18, 652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flem-Karlsen, K.; Fodstad, O.; Tan, M.; Nunes-Xavier, C.E. B7-H3 in Cancer—Beyond Immune Regulation. Trends Cancer 2018, 4, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Quan, Y.; Che, F.; Wang, L. B7-H3 in Tumors: Friend or Foe for Tumor Immunity? Cancer Chemother. Pharmacol. 2018, 81, 245–253. [Google Scholar] [CrossRef]
- Mao, Y.; Chen, L.; Wang, F.; Zhu, D.; Ge, X.; Hua, D.; Sun, J. Cancer Cell-Expressed B7-H3 Regulates the Differentiation of Tumor-Associated Macrophages in Human Colorectal Carcinoma. Oncol. Lett. 2017, 14, 6177–6183. [Google Scholar] [CrossRef] [Green Version]
- Kang, F.-B.; Wang, L.; Li, N.; Zhang, Y.-G.; Sun, D.-X. Hepatocellular Carcinomas Promote Tumor-Associated Macrophage M2-Polarization via Increased B7-H3 Expression. Oncol. Rep. 2014, 33, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Yim, J.; Koh, J.; Kim, S.; Song, S.G.; Ahn, H.K.; Kim, A.Y.; Jeon, Y.K.; Chung, D.H. Effects of B7-H3 Expression on Tumour-Infiltrating Immune Cells and Clinicopathological Characteristics in Non–Small-Cell Lung Cancer. Eur. J. Cancer 2020, 133, 74–85. [Google Scholar] [CrossRef]
- Zhang, G.; Huang, H.; Zhu, Y.; Yu, G.; Gao, X.; Xu, Y.; Liu, C.; Hou, J.; Zhang, X. A Novel Subset of B7-H3+CD14+HLA-DR−/Lowmyeloid-Derived Suppressor Cells are Associated with Progression of Human NSCLC. OncoImmunology 2015, 4, e977164. [Google Scholar] [CrossRef] [Green Version]
- Baumann, T.; Dunkel, A.; Schmid, C.; Schmitt, S.; Hiltensperger, M.; Lohr, K.; Laketa, V.; Donakonda, S.; Ahting, U.; Lorenz-Depiereux, B.; et al. Regulatory Myeloid Cells Paralyze T Cells through Cell–Cell Transfer of the Metabolite Methylglyoxal. Nat. Immunol. 2020, 21, 555–566. [Google Scholar] [CrossRef]
- Si, Y.; Merz, S.F.; Jansen, P.; Wang, B.; Bruderek, K.; Altenhoff, P.; Mattheis, S.; Lang, S.; Gunzer, M.; Klode, J.; et al. Multidimensional Imaging Provides Evidence for Down-Regulation of T Cell Effector Function by MDSC in Human Cancer Tissue. Sci. Immunol. 2019, 4, eaaw9159. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wei, W.; Zhao, Q. B7-H3, a Checkpoint Molecule, as a Target for Cancer Immunotherapy. Int. J. Biol. Sci. 2020, 16, 1767–1773. [Google Scholar] [CrossRef] [Green Version]
- Majzner, R.G.; Theruvath, J.L.; Nellan, A.; Heitzeneder, S.; Cui, Y.; Mount, C.W.; Rietberg, S.P.; Linde, M.H.; Xu, P.; Rota, C.; et al. CAR T Cells Targeting B7-H3, a Pan-Cancer Antigen, Demonstrate Potent Preclinical Activity Against Pediatric Solid Tumors and Brain Tumors. Clin. Cancer Res. 2019, 25, 2560–2574. [Google Scholar] [CrossRef] [PubMed]
- Theruvath, J.; Sotillo, E.; Mount, C.W.; Graef, C.M.; Delaidelli, A.; Heitzeneder, S.; Labanieh, L.; Dhingra, S.; Leruste, A.; Majzner, R.G.; et al. Locoregionally Administered B7-H3-Targeted CAR T Cells for Treatment of Atypical Teratoid/Rhabdoid Tumors. Nat. Med. 2020, 26, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Scribner, J.A.; Brown, J.G.; Son, T.; Chiechi, M.; Li, P.; Sharma, S.; Li, H.; De Costa, A.; Li, Y.; Chen, Y.; et al. Preclinical Development of MGC018, a Duocarmycin-Based Antibody–Drug Conjugate Targeting B7-H3 for Solid Cancer. Mol. Cancer Ther. 2020, 19, 2235–2244. [Google Scholar] [CrossRef]
- Love, I.M.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Gene Names | Gene | Proteomic RMS | Transcriptomic RMS | Proteomic Normal | Transcriptomic Normal | Composite Sum | Composite Rank |
|---|---|---|---|---|---|---|---|---|
| P55283 | CDH4 | CDH4 | 1 | 12 | 1 | 1 | 15 | 1 |
| P52803 | EFNA5 EPLG7 LERK7 | EFNA5 | 1 | 9 | 2 | 4 | 16 | 2 |
| Q92823 | NRCAM KIAA0343 | NRCAM | 4 | 1 | 13 | 6 | 24 | 3 |
| O60245 | PCDH7 BHPCDH | PCDH7 | 9 | 8 | 5 | 5 | 27 | 4 |
| Q15375 | EPHA7 EHK3 HEK11 | EPHA7 | 18 | 3 | 7 | 2 | 30 | 5 |
| Q15223 | NECTIN1 HVEC PRR1 PVRL1 | NECTIN1 | 1 | 2 | 19 | 14 | 36 | 6 |
| O14936 | CASK LIN2 | CASK | 5 | 21 | 12 | 3 | 41 | 7 |
| O60462 | NRP2 VEGF165R2 | NRP2 | 7 | 22 | 3 | 9 | 41 | 7 |
| P54826 | GAS1 | GAS1 | 8 | 14 | 6 | 17 | 45 | 9 |
| Q06787 | FMR1 | FMR1 | 6 | 15 | 16 | 11 | 48 | 10 |
| Q16658 | FSCN1 FAN1 HSN SNL | FSCN1 | 12 | 5 | 14 | 19 | 50 | 11 |
| Q6ZRP7 | QSOX2 QSCN6L1 SOXN | QSOX2 | 20 | 11 | 10 | 10 | 51 | 12 |
| Q9C0H2 | TTYH3 KIAA1691 | TTYH3 | 10 | 7 | 18 | 16 | 51 | 12 |
| O15020 | SPTBN2 KIAA0302 SCA5 | SPTBN2 | 14 | 4 | 20 | 15 | 53 | 14 |
| P32004 | L1CAM CAML1 MIC5 | L1CAM | 15 | 10 | 15 | 13 | 53 | 14 |
| Q5ZPR3 | CD276 B7H3 PSEC0249 UNQ309/PRO352 | CD276 | 23 | 13 | 8 | 12 | 56 | 16 |
| O75051 | PLXNA2 KIAA0463 OCT PLXN2 UNQ209/PRO235 | PLXNA2 | 24 | 16 | 9 | 8 | 57 | 17 |
| P09429 | HMGB1 HMG1 | HMGB1 | 11 | 24 | 4 | 22 | 61 | 18 |
| Q8WXX5 | DNAJC9 | DNAJC9 | 17 | 18 | 22 | 7 | 64 | 19 |
| P49006 | MARCKSL1 MLP MRP | MARCKSL1 | 16 | 6 | 24 | 24 | 70 | 20 |
| Q9H910 | JPT2 C16orf34 HN1L L11 | HN1L | 13 | 19 | 23 | 18 | 73 | 21 |
| P34741 | SDC2 HSPG1 | SDC2 | 22 | 20 | 11 | 21 | 74 | 22 |
| Q15691 | MAPRE1 | MAPRE1 | 19 | 17 | 21 | 23 | 80 | 23 |
| Q14160 | SCRIB CRIB1 KIAA0147 LAP4 SCRB1 VARTUL | SCRIB | 21 | 23 | 17 | 20 | 81 | 24 |
| Entry | Gene Names | Gene | Surfaceomic RMS | Transcriptomic RMS | Proteomic Normal | Transcriptomic Normal | Composite Sum | Composite Rank |
|---|---|---|---|---|---|---|---|---|
| P52803 | EFNA5 EPLG7 LERK7 | EFNA5 | 1 | 10 | 1 | 3 | 15 | 1 |
| Q92823 | NRCAM KIAA0343 | NRCAM | 8 | 4 | 12 | 7 | 31 | 2 |
| Q15223 | NECTIN1 HVEC PRR1 PVRL1 | NECTIN1 | 1 | 1 | 19 | 13 | 34 | 3 |
| O60245 | PCDH7 BHPCDH | PCDH7 | 15 | 11 | 5 | 4 | 35 | 4 |
| Q9BY67 | CADM1 IGSF4 IGSF4A NECL2 SYNCAM TSLC1 | CADM1 | 1 | 19 | 9 | 6 | 35 | 4 |
| P27701 | CD82 KAI1 SAR2 ST6 TSPAN27 | CD82 | 1 | 14 | 2 | 20 | 37 | 6 |
| O14910 | LIN7A MALS1 VELI1 | LIN7A | 1 | 17 | 22 | 1 | 41 | 7 |
| P22455 | FGFR4 JTK2 TKF | FGFR4 | 13 | 3 | 20 | 9 | 45 | 8 |
| O14936 | CASK LIN2 | CASK | 7 | 26 | 11 | 2 | 46 | 9 |
| Q8NBJ4 | GOLM1 C9orf155 GOLPH2 PSEC0242 | GOLM1 | 10 | 13 | 3 | 22 | 48 | 10 |
| P54826 | GAS1 | GAS1 | 12 | 16 | 6 | 18 | 52 | 11 |
| Q06787 | FMR1 | FMR1 | 9 | 18 | 15 | 12 | 54 | 12 |
| Q9C0H2 | TTYH3 KIAA1691 | TTYH3 | 16 | 7 | 18 | 17 | 58 | 13 |
| O15020 | SPTBN2 KIAA0302 SCA5 | SPTBN2 | 18 | 2 | 25 | 14 | 59 | 14 |
| P18065 | IGFBP2 BP2 IBP2 | IGFBP2 | 1 | 5 | 24 | 29 | 59 | 14 |
| P36639 | NUDT1 MTH1 | NUDT1 | 25 | 12 | 23 | 5 | 65 | 16 |
| Q16658 | FSCN1 FAN1 HSN SNL | FSCN1 | 24 | 8 | 14 | 21 | 67 | 17 |
| Q6ZRP7 | QSOX2 QSCN6L1 SOXN | QSOX2 | 26 | 24 | 7 | 10 | 67 | 17 |
| P42574 | CASP3 CPP32 | CASP3 | 14 | 30 | 13 | 11 | 68 | 19 |
| O96019 | ACTL6A BAF53 BAF53A INO80K | ACTL6A | 30 | 23 | 8 | 16 | 77 | 20 |
| P07093 | SERPINE2 PI7 PN1 | SERPINE2 | 27 | 15 | 21 | 15 | 78 | 21 |
| P09429 | HMGB1 HMG1 | HMGB1 | 20 | 28 | 4 | 26 | 78 | 21 |
| P62166 | NCS1 FLUP FREQ | NCS1 | 17 | 9 | 28 | 24 | 78 | 21 |
| Q8WXX5 | DNAJC9 | DNAJC9 | 21 | 22 | 27 | 8 | 78 | 21 |
| P34741 | SDC2 HSPG1 | SDC2 | 28 | 21 | 10 | 25 | 84 | 25 |
| Q14160 | SCRIB CRIB1 KIAA0147 LAP4 SCRB1 VARTUL | SCRIB | 19 | 25 | 17 | 23 | 84 | 26 |
| P05067 | APP A4 AD1 | APP | 11 | 29 | 16 | 30 | 86 | 27 |
| P49006 | MARCKSL1 MLP MRP | MARCKSL1 | 23 | 6 | 30 | 28 | 87 | 28 |
| Q9H910 | JPT2 C16orf34 HN1L L11 | HN1L | 22 | 27 | 29 | 19 | 97 | 28 |
| Q15691 | MAPRE1 | MAPRE1 | 29 | 20 | 26 | 27 | 102 | 30 |
| Entry | Gene Names | Gene | Surfaceomic RMS | Transcriptomic RMS | Proteomic Normal | Transcriptomic Normal | Composite Sum | Composite Rank |
|---|---|---|---|---|---|---|---|---|
| P55283 | CDH4 | CDH4 | 1 | 17 | 1 | 1 | 20 | 1 |
| Q58EX2 | SDK2 KIAA1514 | SDK2 | 1 | 6 | 11 | 4 | 22 | 2 |
| Q96KG7 | MEGF10 KIAA1780 | MEGF10 | 15 | 9 | 1 | 1 | 26 | 3 |
| O00762 | UBE2C UBCH10 | UBE2C | 23 | 2 | 1 | 1 | 27 | 4 |
| P52803 | EFNA5 EPLG7 LERK7 | EFNA5 | 1 | 23 | 4 | 10 | 38 | 5 |
| P49407 | ARRB1 ARR1 | ARRB1 | 1 | 13 | 13 | 18 | 45 | 6 |
| P78310 | CXADR CAR | CXADR | 1 | 26 | 6 | 13 | 46 | 7 |
| Q15375 | EPHA7 EHK3 HEK11 | EPHA7 | 22 | 3 | 17 | 6 | 48 | 8 |
| Q92823 | NRCAM KIAA0343 | NRCAM | 8 | 1 | 26 | 14 | 49 | 9 |
| O60245 | PCDH7 BHPCDH | PCDH7 | 13 | 12 | 15 | 11 | 51 | 10 |
| P24394 | IL4R IL4RA 582J2.1 | IL4R | 1 | 21 | 5 | 31 | 58 | 11 |
| Q15223 | NECTIN1 HVEC PRR1 PVRL1 | NECTIN1 | 1 | 4 | 35 | 21 | 61 | 12 |
| P23468 | PTPRD | PTPRD | 18 | 5 | 34 | 7 | 64 | 13 |
| O60462 | NRP2 VEGF165R2 | NRP2 | 11 | 30 | 9 | 15 | 65 | 14 |
| P08581 | MET | MET | 16 | 36 | 8 | 12 | 72 | 15 |
| P13612 | ITGA4 CD49D | ITGA4 | 38 | 22 | 10 | 5 | 75 | 16 |
| O14936 | CASK LIN2 | CASK | 9 | 33 | 25 | 9 | 76 | 17 |
| P32004 | L1CAM CAML1 MIC5 | L1CAM | 17 | 11 | 29 | 20 | 77 | 18 |
| P52292 | KPNA2 RCH1 SRP1 | KPNA2 | 14 | 16 | 27 | 22 | 79 | 19 |
| Q9P258 | RCC2 KIAA1470 TD60 | RCC2 | 26 | 14 | 12 | 27 | 79 | 19 |
| Q06787 | FMR1 | FMR1 | 10 | 25 | 30 | 17 | 82 | 21 |
| P54826 | GAS1 | GAS1 | 21 | 24 | 16 | 24 | 85 | 22 |
| Q9C0H2 | TTYH3 KIAA1691 | TTYH3 | 19 | 10 | 33 | 23 | 85 | 22 |
| Q6ZRP7 | QSOX2 QSCN6L1 SOXN | QSOX2 | 36 | 15 | 19 | 16 | 86 | 24 |
| P55291 | CDH15 CDH14 CDH3 | CDH15 | 37 | 19 | 23 | 8 | 87 | 25 |
| Q92974 | ARHGEF2 KIAA0651 LFP40 | ARHGEF2 | 12 | 20 | 32 | 26 | 90 | 26 |
| Q16658 | FSCN1 FAN1 HSN SNL | FSCN1 | 28 | 7 | 28 | 29 | 92 | 27 |
| Q5ZPR3 | CD276 B7H3 PSEC0249 UNQ309/PRO352 | CD276 | 29 | 27 | 18 | 19 | 93 | 28 |
| P54760 | EPHB4 HTK MYK1 TYRO11 | EPHB4 | 31 | 32 | 7 | 28 | 98 | 29 |
| Q9BX67 | JAM3 UNQ859/PRO1868 | JAM3 | 33 | 18 | 21 | 30 | 102 | 30 |
| P09429 | HMGB1 HMG1 | HMGB1 | 20 | 38 | 14 | 34 | 106 | 31 |
| P49006 | MARCKSL1 MLP MRP | MARCKSL1 | 34 | 8 | 38 | 37 | 117 | 32 |
| P50895 | BCAM LU MSK19 | BCAM | 27 | 31 | 22 | 38 | 118 | 33 |
| P49327 | FASN FAS | FASN | 32 | 34 | 20 | 33 | 119 | 34 |
| Q9H910 | JPT2 C16orf34 HN1L L11 | HN1L | 30 | 29 | 37 | 25 | 121 | 35 |
| Q15691 | MAPRE1 | MAPRE1 | 25 | 28 | 36 | 35 | 124 | 36 |
| P17612 | PRKACA PKACA | PKACA | 24 | 35 | 31 | 36 | 126 | 37 |
| P34741 | SDC2 HSPG1 | SDC2 | 35 | 37 | 24 | 32 | 128 | 38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavoie, R.R.; Gargollo, P.C.; Ahmed, M.E.; Kim, Y.; Baer, E.; Phelps, D.A.; Charlesworth, C.M.; Madden, B.J.; Wang, L.; Houghton, P.J.; et al. Surfaceome Profiling of Rhabdomyosarcoma Reveals B7-H3 as a Mediator of Immune Evasion. Cancers 2021, 13, 4528. https://doi.org/10.3390/cancers13184528
Lavoie RR, Gargollo PC, Ahmed ME, Kim Y, Baer E, Phelps DA, Charlesworth CM, Madden BJ, Wang L, Houghton PJ, et al. Surfaceome Profiling of Rhabdomyosarcoma Reveals B7-H3 as a Mediator of Immune Evasion. Cancers. 2021; 13(18):4528. https://doi.org/10.3390/cancers13184528
Chicago/Turabian StyleLavoie, Roxane R., Patricio C. Gargollo, Mohamed E. Ahmed, Yohan Kim, Emily Baer, Doris A. Phelps, Cristine M. Charlesworth, Benjamin J. Madden, Liguo Wang, Peter J. Houghton, and et al. 2021. "Surfaceome Profiling of Rhabdomyosarcoma Reveals B7-H3 as a Mediator of Immune Evasion" Cancers 13, no. 18: 4528. https://doi.org/10.3390/cancers13184528