
Comprehensive Profiling of Hypoxia-Related miRNAs Identifies miR-23a-3p Overexpression as a Marker of Platinum Resistance and Poor Prognosis in High-Grade Serous Ovarian Cancer

 , , , , ,
, , , , ,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
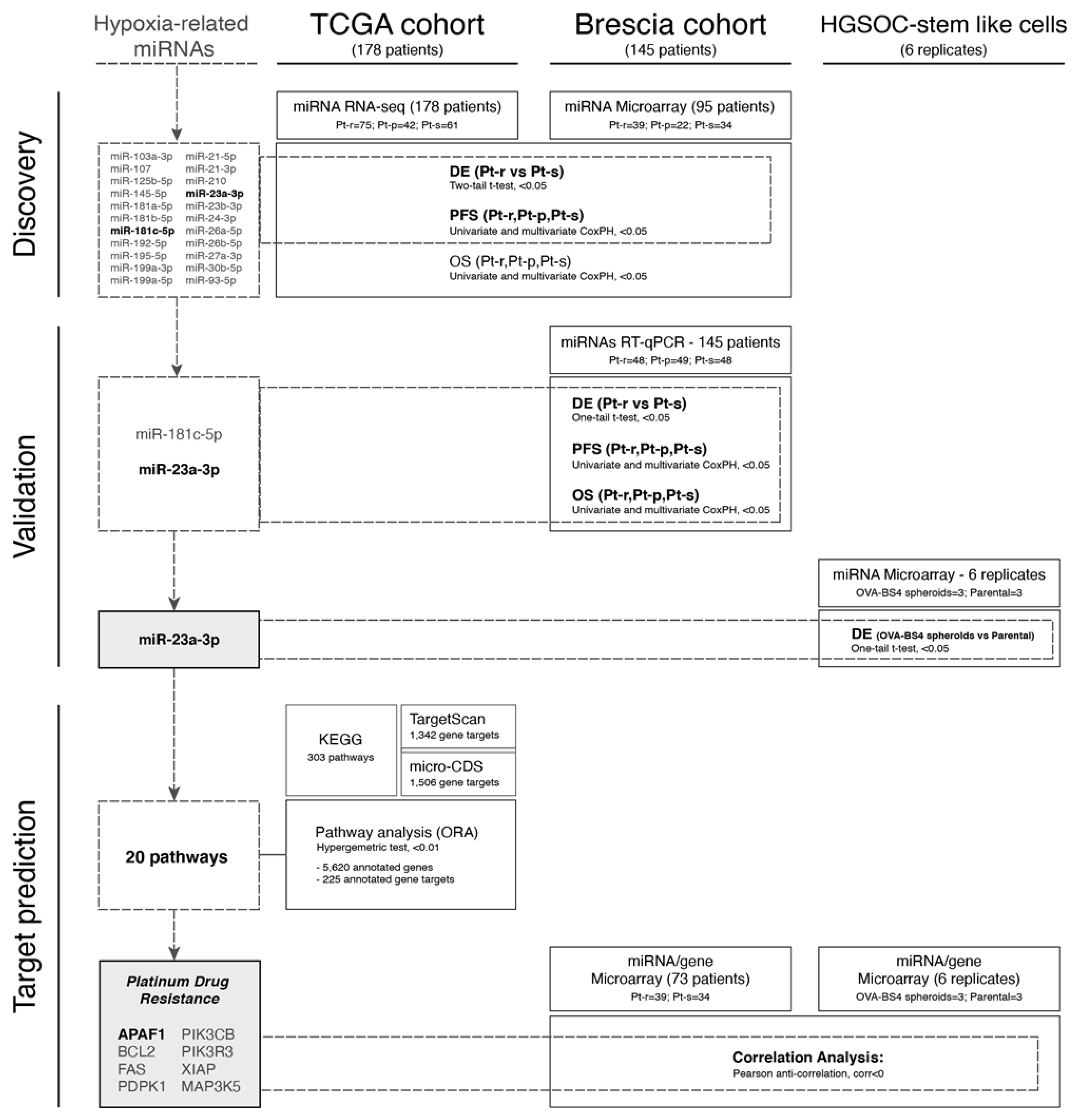
2.1. Selection of Candidate Hypoxia-Related miRNAs
2.2. Patient Sample Cohorts
2.3. Tissue Sample Collection and RNA Extraction
2.4. miRNA and Gene Expression Profiles
2.5. Validation by RT-qPCR
2.6. miRNA Normalization Strategy
2.7. Cell Line Transfection and Apoptotic Cell Death Detection
2.8. Statistical Analysis
2.8.1. Pre-Processing and Differential Expression Analysis
2.8.2. RT-qPCR Data and Equivalence Analysis
2.8.3. Survival Analysis
2.8.4. Over-Representation Analysis
3. Results
3.1. Patient Cohort Description
3.2. Selection of Hypoxia-Related miRNAs
3.3. Evaluation of HRMs in HGSOC Datasets
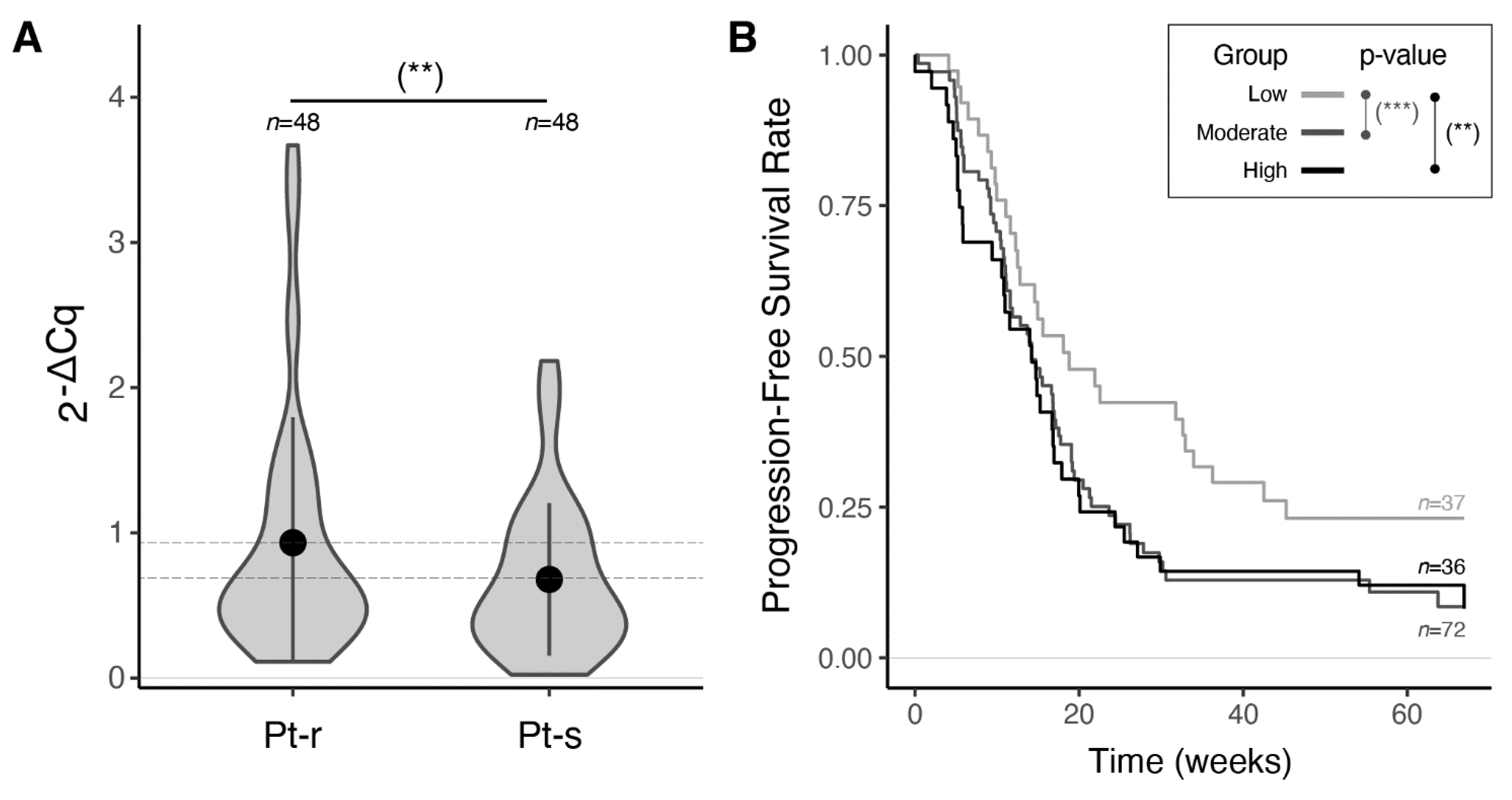
3.4. Validation of miR-23a-3p and miR-181c-5p Expression by RT-qPCR
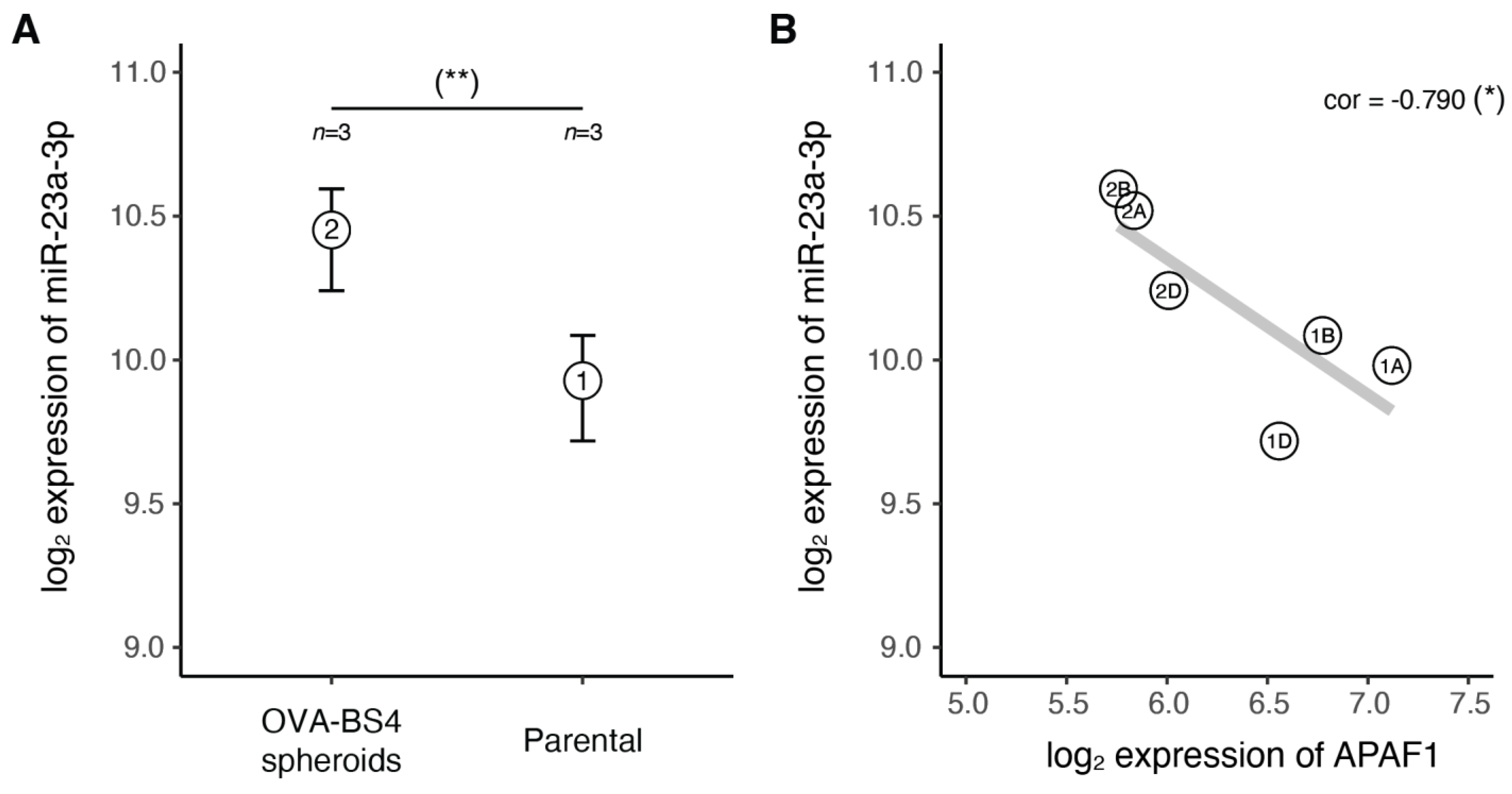
3.5. miR-23a-3p Expression in Ovarian Carcinoma Stem-Like Cells
3.6. In Silico miR-23a-3p Target Prediction and Comparative Pathway Analysis
3.7. The miR-23a-3p/APAF1 Axis and Carboplatin Sensitivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, D.D.; Böhm, S.; Ahmed, A.A.; Aspuria, P.-J.; Bast, R.C.; Beral, V.; Berek, J.S.; Birrer, M.J.; Blagden, S.; Bookman, M.A.; et al. Rethinking ovarian cancer II: Reducing mortality from high-grade serous ovarian cancer. Nat. Rev. Cancer 2015, 15, 668–679. [Google Scholar] [CrossRef]
- Eales, K.L.; Hollinshead, K.E.; Tennant, D.A. Hypoxia and metabolic adaptation of cancer cells. Oncogenesis 2016, 5, e190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Raposo, C.; Mendiola, M.; Barriuso, J.; Casado, E.; Hardisson, D.; Redondo, A. Angiogenesis and ovarian cancer. Clin. Transl. Oncol. 2009, 11, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Paolicchi, E.; Gemignani, F.; Krstic-Demonacos, M.; Dedhar, S.; Mutti, L.; Landi, S. Targeting hypoxic response for cancer therapy. Oncotarget 2016, 7, 13464–13478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.J.; Armaiz Pena, G.N.; Pradeep, S.; Cho, M.S.; Coleman, R.L.; Sood, A.K. Anti-vascular therapies in ovarian cancer: Moving beyond anti-VEGF approaches. Cancer Metastasis Rev. 2015, 34, 19–40. [Google Scholar] [CrossRef] [Green Version]
- Sennino, B.; McDonald, D.M. Controlling escape from angiogenesis inhibitors. Nat. Rev. Cancer 2012, 12, 699–709. [Google Scholar] [CrossRef]
- Aghajanian, C.; Blank, S.V.; Goff, B.A.; Judson, P.L.; Teneriello, M.G.; Husain, A.; Sovak, M.A.; Yi, J.; Nycum, L.R. OCEANS: A Randomized, Double-Blind, Placebo-Controlled Phase III Trial of Chemotherapy With or Without Bevacizumab in Patients With Platinum-Sensitive Recurrent Epithelial Ovarian, Primary Peritoneal, or Fallopian Tube Cancer. J. Clin. Oncol. 2012, 30, 2039–2045. [Google Scholar] [CrossRef] [Green Version]
- Kulshreshtha, R.; Ferracin, M.; Wojcik, S.E.; Garzon, R.; Alder, H.; Agosto-Perez, F.J.; Davuluri, R.; Liu, C.-G.; Croce, C.M.; Negrini, M.; et al. A MicroRNA Signature of Hypoxia. Mol. Cell. Biol. 2007, 27, 1859–1867. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Giannakakis, A.; Sandaltzopoulos, R.; Greshock, J.; Liang, S.; Huang, J.; Hasegawa, K.; Li, C.; O’Brien-Jenkins, A.; Katsaros, D.; Weber, B.L.; et al. miR-210 links hypoxia with cell cycle regulation and is deleted in human epithelial ovarian cancer. Cancer Biol. Ther. 2008, 7, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polytarchou, C.; Iliopoulos, D.; Hatziapostolou, M.; Kottakis, F.; Maroulakou, I.; Struhl, K.; Tsichlis, P.N. Akt2 Regulates All Akt Isoforms and Promotes Resistance to Hypoxia through Induction of miR-21 upon Oxygen Deprivation. Cancer Res. 2011, 71, 4720–4731. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Liu, L.-Z.; Qian, X.; Chen, Q.; Jiang, Y.; Li, D.; Lai, L.; Jiang, B.-H. MiR-145 directly targets p70S6K1 in cancer cells to inhibit tumor growth and angiogenesis. Nucleic Acids Res. 2011, 40, 761–774. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jing, Y.; Li, W.; Qian, X.; Xu, Q.; Li, F.-S.; Liu, L.-Z.; Jiang, B.-H.; Jiang, Y. Roles and Mechanism of miR-199a and miR-125b in Tumor Angiogenesis. PLoS ONE 2013, 8, e56647. [Google Scholar]
- Yeh, Y.-M.; Chuang, C.-M.; Chao, K.-C.; Wang, L.-H. MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis by targeting SOX4 and HIF-1α. Int. J. Cancer 2013, 133, 867–878. [Google Scholar] [CrossRef]
- Joshi, H.P.; Subramanian, I.V.; Schnettler, E.K.; Ghosh, G.; Rupaimoole, R.; Evans, C.; Saluja, M.; Jing, Y.; Cristina, I.; Roy, S.; et al. Dynamin 2 along with microRNA-199a reciprocally regulate hypoxia-inducible factors and ovarian cancer metastasis. Proc. Natl. Acad. Sci. USA 2014, 111, 5331–5336. [Google Scholar] [CrossRef] [Green Version]
- Kinose, Y.; Sawada, K.; Nakamura, K.; Sawada, I.; Toda, A.; Nakatsuka, E.; Hashimoto, K.; Mabuchi, S.; Takahashi, K.; Kurachi, H.; et al. The hypoxia-related microRNA miR-199a-3p displays tumor suppressor functions in ovarian carcinoma. Oncotarget 2015, 6, 11342–11356. [Google Scholar] [CrossRef] [Green Version]
- Rupaimoole, R.; Ivan, C.; Yang, D.; Gharpure, K.M.; Wu, S.Y.; Pecot, C.V.; Previs, R.A.; Nagaraja, A.; Armaiz-Pena, G.N.; McGuire, M.; et al. Hypoxia-upregulated microRNA-630 targets Dicer, leading to increased tumor progression. Oncogene 2016, 35, 4312–4320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfisterer, J.; Ledermann, J.A. Management of Platinum-Sensitive Recurrent Ovarian Cancer. Semin. Oncol. 2006, 33, 12–16. [Google Scholar] [CrossRef]
- Todeschini, P.; Salviato, E.; Paracchini, L.; Ferracin, M.; Petrillo, M.; Zanotti, L.; Tognon, G.; Gambino, A.; Calura, E.; Caratti, G.; et al. Circulating miRNA landscape identifies miR-1246 as promising diagnostic biomarker in high-grade serous ovarian carcinoma: A validation across two independent cohorts. Cancer Lett. 2017, 388, 320–327. [Google Scholar] [CrossRef]
- Benvenuto, G.; Todeschini, P.; Paracchini, L.; Calura, E.; Fruscio, R.; Romani, C.; Beltrame, L.; Martini, P.; Ravaggi, A.; Ceppi, L.; et al. Expression profiles of PRKG1, SDF2L1 and PPP1R12A are predictive and prognostic factors for therapy response and survival in high-grade serous ovarian cancer. Int. J. Cancer 2020, 147, 565–574. [Google Scholar] [CrossRef]
- Bignotti, E.; Calza, S.; Tassi, R.A.; Zanotti, L.; Bandiera, E.; Sartori, E.; Odicino, F.E.; Ravaggi, A.; Todeschini, P.; Romani, C. Identification of stably expressed reference small non-coding RNA s for micro RNA quantification in high-grade serous ovarian carcinoma tissues. J. Cell. Mol. Med. 2016, 20, 2341–2348. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Silva, A.M.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Zanotti, L.; Romani, C.; Tassone, L.; Todeschini, P.; Tassi, R.A.; Bandiera, E.; Damia, G.; Ricci, F.; Ardighieri, L.; Calza, S.; et al. MAL gene overexpression as a marker of high-grade serous ovarian carcinoma stem-like cells that predicts chemoresistance and poor prognosis. BMC Cancer 2017, 17, 366. [Google Scholar] [CrossRef] [Green Version]
- Hauck, W.W.; Anderson, S. A new statistical procedure for testing equivalence in two-group comparative bioavailability trials. J. Pharmacokinet. Biopharm. 1984, 12, 83–91. [Google Scholar] [CrossRef]
- Wellek, S. Testing Statistical Hypotheses of Equivalence and Noninferiority; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-microT web server v5.0: Service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Backes, C.; Keller, A.; Kuentzer, J.; Kneissl, B.; Comtesse, N.; Elnakady, Y.A.; Muller, R.; Meese, E.; Lenhof, H.-P. GeneTrail--advanced gene set enrichment analysis. Nucleic Acids Res. 2007, 35, W186–W192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales, G.; Calura, E.; Cavalieri, D.; Romualdi, C. Graphite—A Bioconductor package to convert pathway topology to gene network. BMC Bioinform. 2012, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA–gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [Green Version]
- Burger, R.A.; Brady, M.F.; Bookman, M.A.; Fleming, G.F.; Monk, B.J.; Huang, H.; Mannel, R.S.; Homesley, H.D.; Fowler, J.; Greer, B.E.; et al. Incorporation of Bevacizumab in the Primary Treatment of Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Tewari, K.S.; Burger, R.A.; Enserro, D.; Norquist, B.M.; Swisher, E.M.; Brady, M.F.; Bookman, M.A.; Fleming, G.F.; Huang, H.; Homesley, H.D.; et al. Final Overall Survival of a Randomized Trial of Bevacizumab for Primary Treatment of Ovarian Cancer. J. Clin. Oncol. 2019, 37, 2317–2328. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, F.L.; Wang, C.W.; Roslani, A.C.; Law, C.W. The Involvement of miR-23a/APAF1 Regulation Axis in Colorectal Cancer. Int. J. Mol. Sci. 2014, 15, 11713–11729. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Liu, M. Correlation analysis on the expression levels of microRNA-23a and microRNA-23b and the incidence and prognosis of ovarian cancer. Oncol. Lett. 2018, 16, 262–266. [Google Scholar] [CrossRef]
- Vaksman, O.; Stavnes, H.T.; Kaern, J.; Trope, C.G.; Davidson, B.; Reich, R. miRNA profiling along tumour progression in ovarian carcinoma. J. Cell. Mol. Med. 2011, 15, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Jin, A.-H.; Wei, Z.-L. Molecular mechanism of increased sensitivity of cisplatin to ovarian cancer by inhibition of microRNA-23a expression. Int. J. Clin. Exp. Med. 2015, 8, 13329–13334. [Google Scholar] [PubMed]
- Yang, Z.; Wang, X.-L.; Bai, R.; Liu, W.-Y.; Li, X.; Liu, M.; Tang, H. miR-23a promotes IKKα expression but suppresses ST7L expression to contribute to the malignancy of epithelial ovarian cancer cells. Br. J. Cancer 2016, 115, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Prahm, K.P.; Høgdall, C.; Karlsen, M.A.; Christensen, I.J.; Novotny, G.W.; Høgdall, E. Identification and validation of potential prognostic and predictive miRNAs of epithelial ovarian cancer. PLoS ONE 2018, 13, e0207319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, R.-J.; Bai, X.-X.; Liu, W. MicroRNA-23a depletion promotes apoptosis of ovarian cancer stem cell and inhibits cell migration by targeting DLG2. Cancer Biol. Ther. 2019, 20, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.L.A.; Co, N.-N.; Tsuruga, T.; Yeung, T.-L.; Kwan, S.-Y.; Leung, C.S.; Li, Y.; Lu, E.S.; Kwan, K.; Wong, K.-K.; et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat. Commun. 2016, 7, 11150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eoh, K.J.; Lee, S.H.; Kim, H.J.; Lee, J.-Y.; Kim, S.; Kim, S.W.; Kim, Y.T.; Nam, E.J. MicroRNA-630 inhibitor sensitizes chemoresistant ovarian cancer to chemotherapy by enhancing apoptosis. Biochem. Biophys. Res. Commun. 2018, 497, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tan, H.-Y.; Feng, Y.-G.; Zhang, C.; Chen, F.; Feng, Y. microRNA-23a in Human Cancer: Its Roles, Mechanisms and Therapeutic Relevance. Cancers 2018, 11, 7. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Clinical Annotations | Brescia Cohort | TCGA Cohort |
|---|---|---|
| No. of Patients | ||
| Total No. of patients | 145 | 178 |
| Age | ||
| Median (range) years | 62 (36–85) | 60 (35–88) |
| Histotype | ||
| Serous | 145 (100%) | 178 (100%) |
| FIGO Classification | ||
| III | 112 (77%) | 159 (89%) |
| IV | 33 (23%) | 19 (11%) |
| Residual Tumor (RT) | ||
| RT = 0 | 41 (28%) | 44 (25%) |
| RT > 0 | 104 (72%) | 134 (75%) |
| Treatment | ||
| Carboplatin + Paclitaxel | 123 (85%) | 169 (95%) |
| Carboplatin + Paclitaxel + Bevacizumab | 22 (15%) | 9 (5%) |
| Platinum Status | ||
| Carboplatin + Paclitaxel | ||
| Sensitive | 48 (33%) | 60 (34%) |
| Partially Sensitive | 23 (16%) | 41 (23%) |
| Resistant | 48 (33%) | 68 (38%) |
| NA | 4 (3%) | - |
| Carboplatin + Paclitaxel + Bevacizumab | ||
| Sensitive | 7 (5%) | 1 (0.5%) |
| Partially Sensitive | 6 (4%) | 1 (0.5%) |
| Resistant | 9 (6%) | 7 (4%) |
| Median follow-up, years (range) | 3.6 (0–15) | 2.62 (0–12.7) |
| Median PFS, months (range) | 23.7 (1.7–172.6) | 15.3 (0.8–111.7) |
| Median OS, months (range) | 43.7 (1.2–177.3) | 33.3 (0.8–152.9) |
| miRNA Name | Brescia Cohort—Microarray 73 Samples (39 Pt-r, 34 Pt-s) | TCGA Cohort—RNA-seq 136 Samples (75 Pt-r, 61 Pt-s) | ||||
|---|---|---|---|---|---|---|
| Log 2(FC) | AveExpr | Adj. p-Value | Log 2(FC) | Mean | Adj. p-Value | |
| hsa-miR-103a-3p | 0.125 | 9.956 | 0.358 | 0.174 | 44316 | 0.324 |
| hsa-miR-107 | 0.058 | 9.317 | 0.682 | 0.131 | 152.4 | 0.323 |
| hsa-miR-125b-5p | 0.493 | 10.508 | 0.040 ** | −0.195 | 10786.2 | 0.327 |
| hsa-miR-145-5p | 0.602 | 7.031 | 0.032 ** | −0.169 | 3572.4 | 0.314 |
| hsa-miR-181a-5p | 0.403 | 7.724 | 0.034 ** | −0.019 | 11434.2 | 0.915 |
| hsa-miR-181b-5p | 0.174 | 5.776 | 0.309 | 0.208 | 1374.8 | 0.228 |
| hsa-miR-181c-5p | 0.547 | 4.896 | 0.036 ** | 0.650 | 223.2 | 0.004 *** |
| hsa-miR-192-5p | −0.082 | 4.712 | 0.740 | 0.241 | 223.4 | 0.230 |
| hsa-miR-195-5p | −0.173 | 7.756 | 0.475 | 0.187 | 6.7 | 0.351 |
| hsa-miR-199a-3p | 0.684 | 9.412 | 0.027 ** | −0.064 | 618.8 | 0.740 |
| hsa-miR-199a-5p | 0.856 | 7.786 | 0.012 ** | −0.308 | 770.8 | 0.161 |
| hsa-miR-21-5p | 0.065 | 13.243 | 0.759 | 0.221 | 11189.6 | 0.268 |
| hsa-miR-21-3p | 0.091 | 7.215 | 0.703 | −0.182 | 2141.6 | 0.305 |
| hsa-miR-210 | 0.023 | 8.001 | 0.910 | −0.035 | 1574.9 | 0.866 |
| hsa-miR-23a-3p | 0.363 | 10.310 | 0.007 *** | 0.286 | 6830.0 | 0.033 ** |
| hsa-miR-23b-3p | −0.177 | 9.554 | 0.389 | 0.183 | 4070.9 | 0.266 |
| hsa-miR-24-3p | −0.008 | 10.460 | 0.945 | 0.252 | 3371.5 | 0.058 * |
| hsa-miR-26a-5p | 0.188 | 10.206 | 0.138 | −0.081 | 1902.6 | 0.572 |
| hsa-miR-26b-5p | 0.106 | 9.162 | 0.480 | −0.015 | 265.4 | 0.917 |
| hsa-miR-27a-3p | 0.440 | 10.151 | 0.002 *** | 0.193 | 1397.1 | 0.208 |
| hsa-miR-30b-5p | −0.313 | 9.202 | 0.089 * | 0.140 | 186.4 | 0.429 |
| hsa-miR-93-5p | −0.238 | 8.468 | 0.109 | 0.051 | 16511.6 | 0.750 |
| miRNA Name | Univariate | Multivariate (1) | ||||
|---|---|---|---|---|---|---|
| Hazard | SE | p-Value | Hazard | SE | p-Value | |
| Brescia Cohort—95 samples | ||||||
| hsa-mir-23a-3p | 1.819 | 0.250 | 0.017 ** | 2.003 | 0.257 | 0.007 *** |
| hsa-mir-181c-5p | 1.349 | 0.111 | 0.007 ** | 1.268 | 0.113 | 0.037 ** |
| TCGA Cohort—178 samples | ||||||
| hsa-mir-23a-3p | 1.072 | 0.112 | 0.539 | 1.076 | 0.113 | 0.517 |
| hsa-mir-181c-5p | 1.026 | 0.070 | 0.715 | 1.036 | 0.072 | 0.627 |
| miRNA Name | Mean (sd) Cq | Mean (sd) −ΔCq (1) | Cq | −ΔCq (1) | ||||
|---|---|---|---|---|---|---|---|---|
| Pt-r | Pt-s | Pt-r | Pt-s | Stat (95%CI) | p-Value | Stat (95%CI) | p-Value | |
| hsa-miR-23a-3p | 24.82 (1.3) | 25.39 (1.3) | −0.56 (1.0) | −1.04 (1.1) | −2.140 (−Inf, −0.128) | 0.017 ** | 1.910 (0.063, Inf) | 0.030 ** |
| hsa-miR-181c-5p | 30.61 (2.3) | 31.09 (2.5) | −6.35 (2.1) | −6.73 (2.6) | −0.965 (−Inf, 0.345) | 0.169 | 0.812 (0.404, Inf) | 0.209 |
| miRNA Name | Univariate-ΔCq (1) | Multivariate (2)-ΔCq (1) | ||||
|---|---|---|---|---|---|---|
| Hazard (95% CI) | SE | p-Value | Hazard (95% CI) | SE | p-Value | |
| Overall Survival (OS) | ||||||
| hsa-miR-23a-3p | 1.149 (0.977–1.352) | 0.083 | 0.092 * | 1.195 (1.027,1.390) | 0.077 | 0.021 ** |
| hsa-miR-181c-5p | 1.026 (0.940–1.121) | 0.045 | 0.561 | 1.025 (0.938,1.119) | 0.045 | 0.587 |
| Progression-Free Survival (PFS) | ||||||
| hsa-miR-23a-3p | 1.178 (1.016–1.366) | 0.075 | 0.030 ** | 1.244 (1.071,1.446) | 0.077 | 0.004 *** |
| hsa-miR-181c-5p | 1.034 (0.957–1.118) | 0.040 | 0.399 | 1.040 (0.960,1.127) | 0.041 | 0.346 |
| Pathway | No. Genes | No. Targets | −log10 (adj. p-Value) (1) | Target | |
|---|---|---|---|---|---|
| 1 | Renal cell carcinoma | 56 | 13 | 15.32 | ARNT; ARNT2; CREBBP; CRK; EGLN2; GAB1; MET; PIK3CB; PIK3R3; PAK3; PAK6; TGFA; RAP1A |
| 2 | Platinum drug resistance | 39 | 8 | 10 | APAF1; BCL2; FAS; PDPK1; PIK3CB; PIK3R3; XIAP; MAP3K5 |
| 3 | Hedgehog signalling pathway | 47 | 8 | 8.21 | CSNK1G1; CSNK1G3; CUL3; HHIP; SMURF2; SPOPL; BCL2; GSK3B |
| 4 | EGFR tyrosine kinase inhibitor resistance | 79 | 11 | 7.41 | BCL2; FGF2; GAB1; IL6R; JAK1; MET; PIK3CB; PIK3R3; PTEN; TGFA; GSK3B |
| 5 | ErbB signalling pathway | 85 | 11 | 6.59 | CRK; ERBB4; GAB1; PAK3; PAK6; PIK3CB; PIK3R3; TGFA; GSK3B; CBLB; STAT5B |
| 6 | Bacterial invasion of epithelial cells | 53 | 7 | 5.57 | CRK; GAB1; MET; PIK3CB; PIK3R3; WASL; DNM3 |
| 7 | Non-small cell lung cancer | 65 | 8 | 5.34 | EML4; PDPK1; PIK3CB; PIK3R3; RXRG; TGFA; STAT5B; STK4 |
| 8 | Glycosphingolipid biosynthesis—lacto and neolacto series | 27 | 4 | 4.72 | FUT4; FUT9; GCNT2; ST8SIA1 |
| 9 | p53 signalling pathway | 71 | 8 | 4.61 | APAF1; BCL2; CCNG1; FAS; PTEN; RCHY1; SESN2; SESN3 |
| 10 | mTOR signalling pathway | 142 | 15 | 4.51 | ATP6V1B2; ATP6V1C1; ATP6V1E1; FNIP2; FZD4; FZD5; GSK3B; LRP5; PDPK1; PIK3CB; PIK3R3; PTEN; SEH1L; SESN2; CHUK |
| 11 | Prostate cancer | 85 | 9 | 4.25 | CHUK; CREBBP; GSK3B; PDPK1; PIK3CB; PIK3R3; PTEN; TGFA; BCL2 |
| 12 | Aldosterone-regulated sodium reabsorption | 30 | 4 | 4.2 | NEDD4L; PDPK1; PIK3CB; PIK3R3 |
| 13 | Measles | 107 | 11 | 4.14 | CBLB; CHUK; JAK1; PIK3CB; PIK3R3; RCHY1; TNFAIP3; GSK3B; FAS; IL12B; STAT5B |
| 14 | Fc gamma R-mediated phagocytosis | 91 | 9 | 3.66 | ASAP1; CRK; PIK3CB; PIK3R3; PRKCE; WASL; CFL2; MARCKS; MARCKSL1 |
| 15 | Adherens junction | 69 | 7 | 3.62 | CSNK2A2; MET; TGFBR2; WASL; YES1; TJP1; NLK |
| 16 | Small cell lung cancer | 92 | 9 | 3.57 | APAF1; BCL2; CHUK; COL4A4; PIK3CB; PIK3R3; PTEN; RXRG; XIAP |
| 17 | Mannose type O-glycan biosynthesis | 23 | 3 | 3.49 | CHST10; FUT4; FUT9 |
| 18 | Non-alcoholic fatty liver disease (NAFLD) | 71 | 7 | 3.42 | FAS; IL6R; MAP3K5; PIK3CB; PIK3R3; GSK3B; CASP7 |
| 19 | Phosphatidylinositol signalling system | 86 | 8 | 3.1 | DGKE; INPP5A; IPMK; PIK3C2A; PIK3CB; PIK3R3; PIP4K2B; PTEN |
| 20 | IL-17 signalling pathway | 14 | 2 | 3.08 | FOSB; TAB3 |
| Symbol | Target Scan | microT−CDS | Tarbase | Pearson Correlation | |
|---|---|---|---|---|---|
| Context Score | miTGscore | Experimentally Validated | Brescia Cohort Microarray | OVA−BS4 Cell Line Microarray | |
| APAF1 | −0.54 | 0.96 | Yes | −0.227 | −0.790 |
| BCL2 | −0.36 | 0.73 | No | 0.250 | 0.893 |
| FAS | −0.51 | 0.84 | No | 0.124 | −0.783 |
| PDPK1 | −0.15 | 0.92 | Yes | −0.159 | −0.002 |
| PIK3CB | −0.17 | 0.91 | Yes | 0.040 | −0.201 |
| PIK3R3 | −0.17 | 0.95 | Yes | −0.126 | 0.927 |
| XIAP | −0.28 | 0.95 | No | 0.052 | −0.907 |
| MAP3K5 | −0.08 | 0.81 | No | −0.038 | 0.816 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todeschini, P.; Salviato, E.; Romani, C.; Raimondi, V.; Ciccarese, F.; Ferrari, F.; Tognon, G.; Marchini, S.; D’Incalci, M.; Zanotti, L.; et al. Comprehensive Profiling of Hypoxia-Related miRNAs Identifies miR-23a-3p Overexpression as a Marker of Platinum Resistance and Poor Prognosis in High-Grade Serous Ovarian Cancer. Cancers 2021, 13, 3358. https://doi.org/10.3390/cancers13133358
Todeschini P, Salviato E, Romani C, Raimondi V, Ciccarese F, Ferrari F, Tognon G, Marchini S, D’Incalci M, Zanotti L, et al. Comprehensive Profiling of Hypoxia-Related miRNAs Identifies miR-23a-3p Overexpression as a Marker of Platinum Resistance and Poor Prognosis in High-Grade Serous Ovarian Cancer. Cancers. 2021; 13(13):3358. https://doi.org/10.3390/cancers13133358
Chicago/Turabian StyleTodeschini, Paola, Elisa Salviato, Chiara Romani, Vittoria Raimondi, Francesco Ciccarese, Federico Ferrari, Germana Tognon, Sergio Marchini, Maurizio D’Incalci, Laura Zanotti, and et al. 2021. "Comprehensive Profiling of Hypoxia-Related miRNAs Identifies miR-23a-3p Overexpression as a Marker of Platinum Resistance and Poor Prognosis in High-Grade Serous Ovarian Cancer" Cancers 13, no. 13: 3358. https://doi.org/10.3390/cancers13133358