
Challenging, Accurate and Feasible: CAF-1 as a Tumour Proliferation Marker of Diagnostic and Prognostic Value


Abstract
:Simple Summary
Abstract
1. Introduction
Proliferation Markers as Prognostic Tools in Cancer
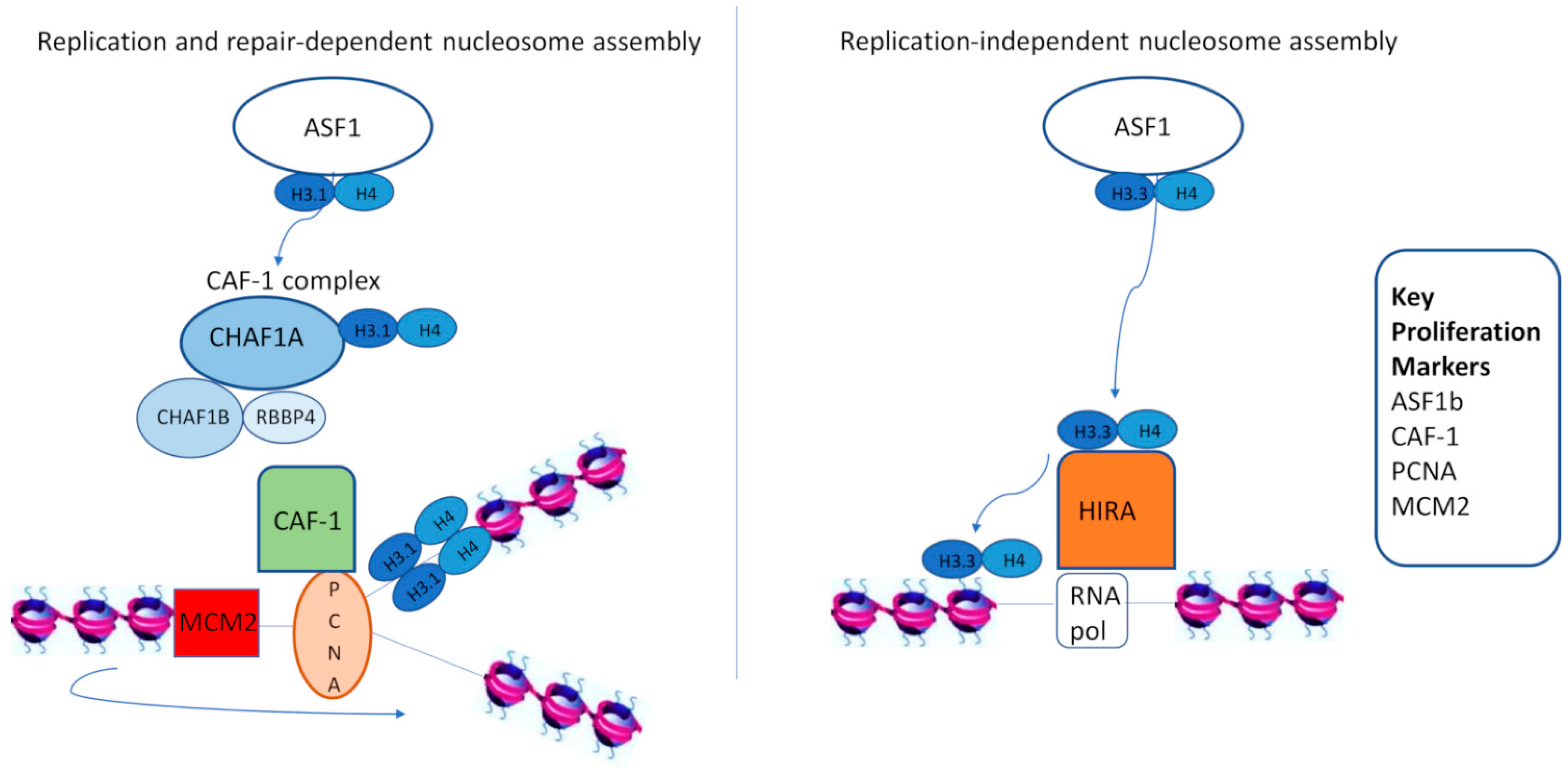
2. CAF-1 as a Proliferation Marker
3. CAF-1 Expression in Tumours: Correlation with Clinicopathologic Data and Survival
CAF-1 Expression in Hematologic Malignancies
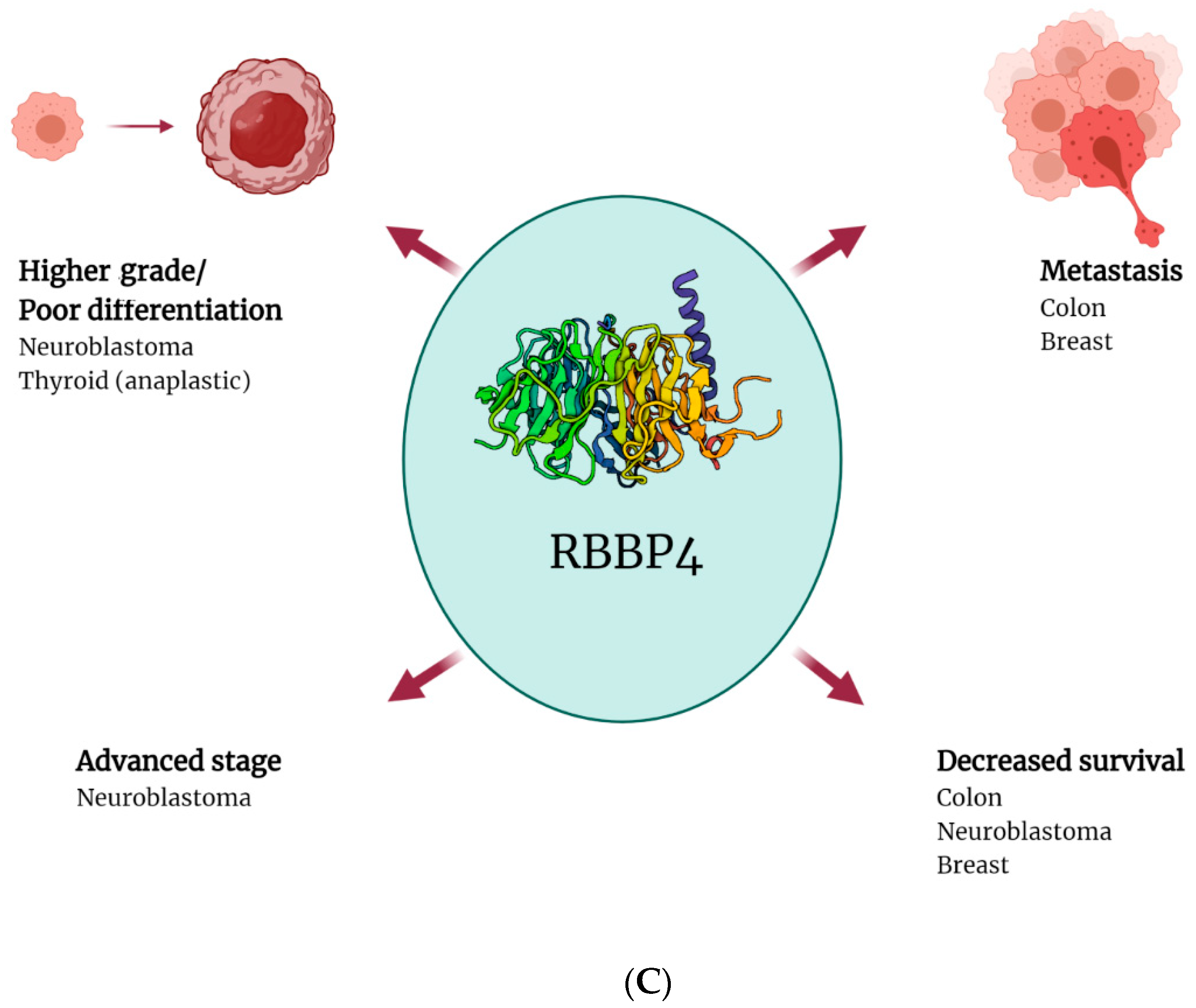
4. CAF-1 Expression in Solid Tumours
4.1. Gynaecologic Cancers
4.2. Lung Cancer
4.3. Gastrointestinal Cancers
4.4. Urogenital Cancer
4.5. Nervous System Tumours
4.6. Breast Cancer
4.7. Skin Cancer
4.8. Head and Neck Cancers
5. Mechanistic Insights-Targeting CAF-1
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wild, C.; Weiderpass, E.; Stewart, B. World Cancer Report: Cancer Research for Cancer Prevention; IARC Press: Lyon, France, 2020; pp. 23–33. [Google Scholar]
- Whitfield, M.L.; George, L.K.; Grant, G.D.; Perou, C.M. Common markers of proliferation. Nat. Rev. Cancer. 2006, 6, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Pezzilli, R.; Partelli, S.; Cannizzaro, R.; Pagano, N.; Crippa, S.; Pagnanelli, M.; Falconi, M. Ki-67 prognostic and therapeutic decision driven marker for pancreatic neuroendocrine neoplasms (PNENs): A systematic review. Adv. Med. Sci. 2016, 61, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Sobecki, M.; Mrouj, K.; Camasses, A.; Parisis, N.; Nicolas, E.; Llères, D.; Gerbe, F.; Prieto, S.; Krasinska, L.; David, A. The cell proliferation antigen Ki-67 organises heterochromatin. elife 2016, 5, e13722. [Google Scholar] [CrossRef]
- Sobecki, M.; Mrouj, K.; Colinge, J.; Gerbe, F.; Jay, P.; Krasinska, L.; Dulic, V.; Fisher, D. Cell-cycle regulation accounts for variability in Ki-67 expression levels. Cancer Res. 2017, 77, 2722–2734. [Google Scholar] [CrossRef] [Green Version]
- Rindi, G.; Klimstra, D.S.; Abedi-Ardekani, B.; Asa, S.L.; Bosman, F.T.; Brambilla, E.; Busam, K.J.; Krijger, R.R.D.; Dietel, M.; El-Naggar, A.K.; et al. A common classification framework for neuroendocrine neoplasms: An International Agency for Research on Cancer (IARC) and World Health Organization (WHO) expert consensus proposal. Mod. Pathol. 2018, 31, 1770–1786. [Google Scholar] [CrossRef]
- Luo, Z.W.; Zhu, M.G.; Zhang, Z.Q.; Ye, F.J.; Huang, W.H.; Luo, X.Z. Increased expression of Ki-67 is a poor prognostic marker for colorectal cancer patients: A meta analysis. BMC Cancer 2019, 19, 123. [Google Scholar] [CrossRef] [Green Version]
- Smith, I.; Robertson, J.; Kilburn, L.; Wilcox, M.; Evans, A.; Holcombe, C.; Horgan, K.; Kirwan, C.; Mallon, E.; Sibbering, M.; et al. Long-term outcome and prognostic value of Ki67 after perioperative endocrine therapy in postmenopausal women with hormone-sensitive early breast cancer (POETIC): An open-label, multicentre, parallel-group, randomised, phase 3 trial. Lancet Oncol. 2020, 21, 1443–1454. [Google Scholar] [CrossRef]
- Walts, A.E.; Mirocha, J.M.; Marchevsky, A.M. Challenges in Ki-67 assessments in pulmonary large-cell neuroendocrine carcinomas. Histopathology 2020, 78, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.O.; Leung, S.C.; Rimm, D.L.; Dodson, A.; Acs, B.; Badve, S.; Denkert, C.; Ellis, M.J.; Fineberg, S.; Flowers, M. Assessment of Ki67 in breast cancer: Updated recommendations from the International Ki67 in Breast Cancer Working Group. J. Nat. Cancer Inst. 2020. [Google Scholar] [CrossRef]
- Acs, B.; Pelekanou, V.; Bai, Y.; Martinez-Morilla, S.; Toki, M.; Leung, S.C.; Nielsen, T.O.; Rimm, D.L. Ki67 reproducibility using digital image analysis: An inter-platform and inter-operator study. Lab. Investig. 2019, 99, 107–117. [Google Scholar] [CrossRef]
- Rimm, D.L.; Leung, S.C.; McShane, L.M.; Bai, Y.; Bane, A.L.; Bartlett, J.M.; Bayani, J.; Chang, M.C.; Dean, M.; Denkert, C. An international multicenter study to evaluate reproducibility of automated scoring for assessment of Ki67 in breast cancer. Mod. Pathol. 2019, 32, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Aung, T.N.; Acs, B.; Warrell, J.; Bai, Y.; Gaule, P.; Martinez-Morilla, S.; Vathiotis, I.; Shafi, S.; Moutafi, M.; Gerstein, M. A new tool for technical standardization of the Ki67 immunohistochemical assay. Mod. Pathol. 2021, 1–10. [Google Scholar] [CrossRef]
- Rodel, F.; Zhou, S.; Gyorffy, B.; Raab, M.; Sanhaji, M.; Mandal, R.; Martin, D.; Becker, S.; Strebhardt, K. The prognostic relevance of the proliferation markers Ki-67 and Plk1 in early-stage ovarian cancer patients with serous, low-grade carcinoma based on mRNA and protein expression. Front. Oncol. 2020, 10, 558932. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.; Min, M.; Yang, C.; Tian, C.; Gookin, S.; Carter, D.; Spencer, S.L. Ki67 is a graded rather than a binary marker of proliferation versus quiescence. Cell Rep. 2018, 24, 1105–1112.e5. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, S.E.; Skopelitou, A.S.; Margeli, A.P.; Pavlaki, K.J.; Kittas, C. Proliferating cell nuclear antigen (PCNA) expression in regenerating rat liver after partial hepatectomy. Dig. Dis. Sci. 1994, 39, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Jurikova, M.; Danihel, L.; Polak, S.; Varga, I. Ki67, PCNA, and MCM proteins: Markers of proliferation in the diagnosis of breast cancer. Acta Histochem. 2016, 118, 544–552. [Google Scholar] [CrossRef]
- Stoeber, K.; Tlsty, T.D.; Happerfield, L.; Thomas, G.A.; Romanov, S.; Bobrow, L.; Williams, E.D.; Williams, G.H. DNA replication licensing and human cell proliferation. J. Cell Sci. 2001, 114, 2027–2041. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wang, G.; Shi, Y.; Xu, H.; Zheng, Y.; Chen, Y. MCMs in cancer: Prognostic potential and mechanisms. Anal. Cell Pathol. 2020, 2020, 3750294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, H.; Zhang, J.; Cheng, A.S.L.; Yu, J.; To, K.F.; Kang, W. MCM family in gastrointestinal cancer and other malignancies: From functional characterization to clinical implication. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188415. [Google Scholar] [CrossRef] [PubMed]
- Giaginis, C.; Georgiadou, M.; Dimakopoulou, K.; Tsourouflis, G.; Gatzidou, E.; Kouraklis, G.; Theocharis, S. Clinical significance of MCM-2 and MCM-5 expression in colon cancer: Association with clinicopathological parameters and tumor proliferative capacity. Dig. Dis. Sci. 2009, 54, 282–291. [Google Scholar] [CrossRef]
- Giaginis, C.; Vgenopoulou, S.; Vielh, P.; Theocharis, S. MCM proteins as diagnostic and prognostic tumor markers in the clinical setting. Histol. Histopathol. 2010, 25, 351–370. [Google Scholar] [CrossRef]
- Corpet, A.; Koning, L.D.; Toedling, J.; Savignoni, A.; Berger, F.; Lemaître, C.; O’Sullivan, R.J.; Karlseder, J.; Barillot, E.; Asselain, B. Asf1b, the necessary Asf1 isoform for proliferation, is predictive of outcome in breast cancer. EMBO J. 2011, 30, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Corpet, A.; Gurard-Levin, Z.A.; Juan, D.; Ochsenbein, F.; Rico, D.; Valencia, A.; Almouzni, G. Subfunctionalization via adaptive evolution influenced by genomic context: The case of histone chaperones ASF1a and ASF1b. Mol. Biol. Evol. 2013, 30, 1853–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koning, L.D.; Savignoni, A.; Boumendil, C.; Rehman, H.; Asselain, B.; Sastre-Garau, X.; Almouzni, G. Heterochromatin protein 1α: A hallmark of cell proliferation relevant to clinical oncology. EMBO Mol. Med. 2009, 1, 178–191. [Google Scholar] [CrossRef]
- Saha, A.K.; Contreras-Galindo, R.; Niknafs, Y.S.; Iyer, M.; Qin, T.; Padmanabhan, K.; Siddiqui, J.; Palande, M.; Wang, C.; Qian, B. The role of the histone H3 variant CENPA in prostate cancer. J. Biol. Chem. 2020, 295, 8537–8549. [Google Scholar] [CrossRef]
- Oca, R.M.D.; Gurard-Levin, Z.A.; Berger, F.; Rehman, H.; Martel, E.; Corpet, A.; Koning, L.D.; Vassias, I.; Wilson, L.O.W.; Meseure, D.; et al. The histone chaperone HJURP is a new independent prognostic marker for luminal A breast carcinoma. Mol. Oncol. 2015, 9, 657–674. [Google Scholar] [CrossRef]
- Polo, S.E.; Theocharis, S.E.; Klijanienko, J.; Savignoni, A.; Asselain, B.; Vielh, P.; Almouzni, G. Chromatin assembly factor-1, a marker of clinical value to distinguish quiescent from proliferating cells. Cancer Res. 2004, 64, 2371–2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polo, S.E.; Theocharis, S.E.; Grandin, L.; Gambotti, L.; Antoni, G.; Savignoni, A.; Asselain, B.; Patsouris, E.; Almouzni, G. Clinical significance and prognostic value of chromatin assembly factor-1 overexpression in human solid tumours. Histopathology 2010, 57, 716–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray-Gallet, D.; Almouzni, G. The Histone H3 Family and Its Deposition Pathways. In Histone Mutations and Cancer; Springer: Amsterdam, The Netherlands, 2021; pp. 17–42. [Google Scholar]
- Smith, S.; Stillman, B. Purification and characterization of CAF-I, a human cell factor required for chromatin assembly during DNA replication in vitro. Cell 1989, 58, 15–25. [Google Scholar] [CrossRef]
- Kaufman, P.D.; Kobayashi, R.; Kessler, N.; Stillman, B. The p150 and p60 subunits of chromatin assembly factor I: A molecular link between newly synthesized histones and DNA replication. Cell 1995, 81, 1105–1114. [Google Scholar] [CrossRef] [Green Version]
- Verreault, A.; Kaufman, P.D.; Kobayashi, R.; Stillman, B. Nucleosome assembly by a complex of CAF-1 and acetylated histones H3/H4. Cell 1996, 87, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Gaillard, P.H.L.; Martini, E.M.; Kaufman, P.D.; Stillman, B.; Moustacchi, E.; Almouzni, G. Chromatin assembly coupled to DNA repair: A new role for chromatin assembly factor I. Cell 1996, 86, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Cheloufi, S.; Elling, U.; Hopfgartner, B.; Jung, Y.L.; Murn, J.; Ninova, M.; Hubmann, M.; Badeaux, A.I.; Ang, C.E.; Tenen, D.; et al. The histone chaperone CAF-1 safeguards somatic cell identity. Nature 2015, 528, 218–224. [Google Scholar] [CrossRef]
- Adam, S.; Polo, S.E.; Almouzni, G. How to restore chromatin structure and function in response to DNA damage—Let the chaperones play. FEBS J. 2014, 281, 2315–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volk, A.; Crispino, J.D. The role of the chromatin assembly complex (CAF-1) and its p60 subunit (CHAF1b) in homeostasis and disease. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Petryk, N.; Dalby, M.; Wenger, A.; Stromme, C.B.; Strandsby, A.; Andersson, R.; Groth, A. MCM2 promotes symmetric inheritance of modified histones during DNA replication. Science 2018, 361, 1389–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinge, S.; Bliss-Moreau, M.; Kirsammer, G.; Diebold, L.; Chlon, T.; Gurbuxani, S.; Crispino, J.D. Increased dosage of the chromosome 21 ortholog Dyrk1a promotes megakaryoblastic leukemia in a murine model of Down syndrome. J. Clin. Investig. 2012, 122, 948–962. [Google Scholar] [CrossRef] [Green Version]
- Volk, A.; Liang, K.; Suraneni, P.; Li, X.; Zhao, J.; Bulic, M.; Marshall, S.; Pulakanti, K.; Malinge, S.; Taub, J.; et al. A CHAF1B-dependent molecular switch in hematopoiesis and leukemia pathogenesis. Cancer Cell 2018, 34, 707–723.e7. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, X.; Zhang, Z. CHAF1B overexpression: A brake for the differentiation of leukemia cells. Cancer Cell 2018, 34, 693–694. [Google Scholar] [CrossRef] [Green Version]
- Casas, S.; Ollila, J.; Aventín, A.; Vihinen, M.; Sierra, J.; Knuutila, S. Changes in apoptosis-related pathways in acute myelocytic leukemia. Cancer Genet. Cytogenet. 2003, 146, 89–101. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, C.; Trudeau, S.J.; Narita, Y.; Zhao, B.; Teng, M.; Guo, R.; Gewurz, B.E. Histone loaders CAF1 and HIRA restrict Epstein-Barr virus B-Cell lytic reactivation. mBio 2020, 11. [Google Scholar] [CrossRef]
- Mascolo, M.; Travaglino, A.; Varricchio, S.; Russo, D.; Sabattini, E.; Agostinelli, C.; Bertuzzi, C.; Baldo, A.; Pileri, A.; Picardi, M.; et al. Role of chromatin assembly factor-1/p60 and poly [ADP-ribose] polymerase 1 in mycosis fungoides. Virchows Arch. 2020, 478, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Long, Q.; Chen, M.; Liu, X.; Zhou, H. CAF-1/p150 promotes cell proliferation, migration, invasion and predicts a poor prognosis in patients with cervical cancer. Oncol. Lett. 2020, 20, 2338–2346. [Google Scholar] [CrossRef]
- Kong, L.; Yu, X.P.; Bai, X.H.; Zhang, W.F.; Zhang, Y.; Zhao, W.M.; Jia, J.H.; Tang, W.; Zhou, Y.B.; Liu, C.J. RbAp48 is a critical mediator controlling the transforming activity of human papillomavirus type 16 in cervical cancer. J. Biol. Chem. 2007, 282, 26381–26391. [Google Scholar] [CrossRef] [Green Version]
- Xia, D.; Yang, X.; Liu, W.; Shen, F.; Pan, J.; Lin, Y.; Du, N.; Sun, Y.; Xi, X. Over-expression of CHAF1A in epithelial ovarian cancer can promote cell proliferation and inhibit cell apoptosis. Biochem. Biophys. Res. Commun. 2017, 486, 191–197. [Google Scholar] [CrossRef]
- Liu, T.; Wei, J.; Jiang, C.; Wang, C.; Zhang, X.; Du, Y.; Li, J.; Zhao, H. CHAF1A, the largest subunit of the chromatin assembly factor 1 complex, regulates the growth of H1299 human non-small cell lung cancer cells by inducing G0/G1 cell cycle arrest. Exp. Ther. Med. 2017, 14, 4681–4686. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Duan, Y.; Liu, T.; Li, L.; Li, W.; Li, J. Over-expression of chromatin assembly factor 1 subunit A (CHAF1A) facilitates cell proliferation in non-small cell lung cancer (NSCLC). Biotechnol. Biotechnol. Equip. 2019, 33, 894–901. [Google Scholar] [CrossRef]
- Cai, Y.; Dong, Z.Y.; Wang, J.Y. MiR-520b inhibited metastasis and proliferation of non-small cell lung cancer by targeting CHAF1A. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7742–7749. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, T.; Li, S.; Huang, M.; Li, X.; Zhao, H.; Li, J. CHAF1B promotes proliferation and reduces apoptosis in 95-D lung cancer cells and predicts a poor prognosis in non-small cell lung cancer. Oncol. Rep. 2019, 41, 2518–2528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Cui, F.; Yu, F.; Peng, X.; Jiang, T.; Chen, D.; Lu, S.; Tang, H.; Peng, Z. Up-regulation of CHAF1A, a poor prognostic factor, facilitates cell proliferation of colon cancer. Biochem. Biophys. Res. Commun. 2014, 449, 208–215. [Google Scholar] [CrossRef]
- Li, Y.D.; Lv, Z.; Xie, H.Y.; Zheng, S.S. Retinoblastoma binding protein 4 up-regulation is correlated with hepatic metastasis and poor prognosis in colon cancer patients. Hepatobiliary Pancreat. Dis. Int. 2019, 18, 446–451. [Google Scholar] [CrossRef]
- Li, Y.D.; Lv, Z.; Zhu, W.F. RBBP4 promotes colon cancer malignant progression. World J. Gastroenterol. 2020, 26, 5328–5342. [Google Scholar] [CrossRef]
- Zheng, L.; Liang, X.; Li, S.; Li, T.; Shang, W.; Ma, L.; Jia, X.; Shao, W.; Sun, P.; Chen, C.; et al. CHAF1A interacts with TCF4 to promote gastric carcinogenesis via upregulation of c-MYC and CCND1 expression. EBioMedicine 2018, 38, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Li, X.; Shen, B.; Chen, X.; Shu, Y. Histone chaperone CHAF1A impacts the outcome of fluoropyrimidines-based adjuvant therapy in gastric cancer by regulating the expression of thymidylate synthetase. Gene 2019, 716, 144034. [Google Scholar] [CrossRef]
- Xu, M.; Jia, Y.; Liu, Z.; Ding, L.; Tian, R.; Gu, H.; Wang, Y.; Zhang, H.; Tu, K.; Liu, Q. Chromatin assembly factor 1, subunit A (P150) facilitates cell proliferation in human hepatocellular carcinoma. OncoTargets Ther. 2016, 9, 4023–4035. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Xia, S.L.; Liao, C.; Li, Y.L.; Wang, Y.F.; Li, T.P.; Zhao, M.J. Genes encoding Pir51, Beclin 1, RbAp48 and aldolase b are up or down-regulated in human primary hepatocellular carcinoma. World J. Gastroenterol. 2004, 10, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascolo, M.; Ilardi, G.; Merolla, F.; Russo, D.; Vecchione, M.L.; Rosa, G.D.; Staibano, S. Tissue microarray-based evaluation of Chromatin Assembly Factor-1 (CAF-1)/p60 as tumour prognostic marker. Int. J. Mol. Sci. 2012, 13, 11044–11062. [Google Scholar] [CrossRef] [PubMed]
- Staibano, S.; Mascolo, M.; Mancini, F.P.; Kisslinger, A.; Salvatore, G.; Benedetto, M.D.; Chieffi, P.; Altieri, V.; Prezioso, D.; Ilardi, G.; et al. Overexpression of chromatin assembly factor-1 (CAF-1) p60 is predictive of adverse behaviour of prostatic cancer. Histopathology 2009, 54, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Du, B.; Jiang, H.; Gao, J. Over-expression of CHAF1A promotes cell proliferation and apoptosis resistance in glioblastoma cells via AKT/FOXO3a/Bim pathway. Biochem. Biophys. Res. Commun. 2016, 469, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Tayrac, M.D.; Saikali, S.; Aubry, M.; Bellaud, P.; Boniface, R.; Quillien, V.; Mosser, J. Prognostic significance of EDN/RB, HJURP, p60/CAF-1 and PDLI4, four new markers in high-grade gliomas. PLoS ONE 2013, 8, e73332. [Google Scholar] [CrossRef] [Green Version]
- Tayrac, M.D.; Aubry, M.; Saikali, S.; Etcheverry, A.; Surbled, C.; Guenot, F.; Galibert, M.-D.; Hamlat, A.; Lesimple, T.; Quillien, V.; et al. A 4-Gene signature associated with clinical outcome in high-grade gliomas. Clin. Cancer Res. 2011, 17, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, E.; Preter, K.D.; Capasso, M.; Johansson, P.; Man, T.-K.; Chen, Z.; Stowers, P.; Tonini, G.P.; Speleman, F.; Shohet, J.M. A p53 drug response signature identifies prognostic genes in high-risk neuroblastoma. PLoS ONE 2013, 8, e79843. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, E.; Preter, K.D.; Capasso, M.; Chen, Z.; Hsu, D.M.; Tonini, G.P.; Lefever, S.; Hicks, J.; Versteeg, R.; Pession, A.; et al. Histone chaperone CHAF1A inhibits differentiation and promotes aggressive neuroblastoma. Cancer Res. 2014, 74, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Song, H.; Mei, H.; Fang, E.; Wang, X.; Yang, F.; Li, H.; Chen, Y.; Huang, K.; Zheng, L.; et al. Armadillo repeat containing 12 promotes neuroblastoma progression through interaction with retinoblastoma binding protein 4. Nat. Commun. 2018, 9, 2829. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Cheng, K.; Wang, X.; Li, X.; Yu, Y.; Hua, Y.; Yang, Z. Expression of HDAC1 and RBBP4 correlate with clinicopathologic characteristics and prognosis in breast cancer. Int. J. Clin. Exp. Pathol. 2020, 13, 563–572. [Google Scholar] [PubMed]
- Mascolo, M.; Vecchione, M.L.; Ilardi, G.; Scalvenzi, M.; Molea, G.; Benedetto, M.D.; Nugnes, L.; Siano, M.; Rosa, G.D.; Staibano, S. Overexpression of chromatin assembly factor-1/p60 helps to predict the prognosis of melanoma patients. BMC Cancer 2010, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Staibano, S.; Pepe, S.; Muzio, L.L.; Somma, P.; Mascolo, M.; Argenziano, G.; Scalvenzi, M.; Salvatore, G.; Fabbrocini, G.; Molea, G.; et al. Poly(adenosine diphosphate-ribose) polymerase 1 expression in malignant melanomas from photoexposed areas of the head and neck region. Hum. Pathol. 2005, 36, 724–731. [Google Scholar] [CrossRef]
- Russo, D.; Travaglino, A.; Varricchio, S.; Merolla, F.; Ilardi, G.; Raffone, A.; Scalvenzi, M.; Costa, C.; Fabbrocini, G.; Staibano, S.; et al. BRIT-1 expression and its relationship with PARP-1 and CAF-1/p60 in cutaneous melanoma. J. Eur. Acad. Dermatol. Venereol. 2020, 35, e254–e257. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, F.; Paolillo, M.; Chiappetta, G.; Crescenzi, E.; Arena, S.; Scaloni, A.; Monaco, M.; Vascotto, C.; Tell, G.; Formisano, S.; et al. RbAp48 is a target of nuclear factor-kappaB activity in thyroid cancer. J. Clin. Endocrinol. Metab. 2007, 92, 1458–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staibano, S.; Mascolo, M.; Rocco, A.; Muzio, L.L.; Ilardi, G.; Siano, M.; Pannone, G.; Vecchione, M.L.; Nugnes, L.; Califano, L.; et al. The proliferation marker Chromatin Assembly Factor-1 is of clinical value in predicting the biological behaviour of salivary gland tumours. Oncol. Rep. 2010, 25. [Google Scholar] [CrossRef] [Green Version]
- Staibano, S.; Mignogna, C.; Muzio, L.L.; Mascolo, M.; Salvatore, G.; Benedetto, M.D.; Califano, L.; Rubini, C.; Rosa, G.D. Chromatin assembly factor-1 (CAF-1)-mediated regulation of cell proliferation and DNA repair: A link with the biological behaviour of squamous cell carcinoma of the tongue? Histopathology 2007, 50, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Mascolo, M.; Ilardi, G.; Romano, M.F.; Celetti, A.; Siano, M.; Romano, S.; Luise, C.; Merolla, F.; Rocco, A.; Vecchione, M.L.; et al. Overexpression of chromatin assembly factor-1 p60, poly(ADP-ribose) polymerase 1 and nestin predicts metastasizing behaviour of oral cancer. Histopathology 2012, 61, 1089–1105. [Google Scholar] [CrossRef] [Green Version]
- Morra, F.; Merolla, F.; Picardi, I.; Russo, D.; Ilardi, G.; Varricchio, S.; Liotti, F.; Pacelli, R.; Palazzo, L.; Mascolo, M.; et al. CAF-1 subunits levels suggest combined treatments with PARP-inhibitors and ionizing radiation in advanced HNSCC. Cancers 2019, 11, 1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.P.; Ilter, D.; Low, V.; Rosenzweig, A.; Shen, Z.-J.; Schild, T.; Rivas, M.A.; Er, E.E.; Mcnally, D.R.; Mutvei, A.P.; et al. Dynamic incorporation of histone H3 variants into chromatin is essential for acquisition of aggressive traits and metastatic colonization. Cancer Cell 2019, 36, 402–417.e13. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tumour | CAF-1 Subunit | Clinicopathologic Parameters | Survival | Reference |
|---|---|---|---|---|
| AML | CHAF1B mRNA | - | ↓ OS | [39,40] |
| RBBP4 mRNA | - | - | [42] | |
| MF | CHAF1B | - | ↓ OS | [44] |
| Cervical cancer | CHAF1A | Advanced stage Local recurrence Distant metastasis | ↓ OS | [45] |
| CHAF1B | Histological grade | ↓ OS | [29] | |
| RBBP4 | - | - | [46] | |
| Endometrial cancer | CHAF1B | Histologic grade Advanced stage | ↓ OS | [29] |
| Ovarian cancer | CHAF1A | Advanced stage Lymph node metastasis | ↓ OS | [47] |
| NSCLC | CHAF1A (mRNA) CHAF1A (protein) | - Local recurrence Metastasis | - ↓ OS and DFS | [48] [49] |
| CHAF1B | Male gender Smokers Squamous subtype Increased tumour size Advanced stage | ↓ OS and DFS | [51] | |
| Colon cancer | CHAF1A | Advanced stage Tumour invasion Poor differentiation | ↓ OS and DFS | [52] |
| CHAF1B | No correlation | - | [29] | |
| RBBP4 | Vascular invasion Lymph node spread Hepatic metastasis | ↓ OS | [53] | |
| Gastric cancer | CHAF1A | - | ↓ OS | [55] |
| CHAF1A | Resistance to 5-FU chemotherapy (↓ OS and DFS) | [56] | ||
| CHAF1B | No correlation | - | [29] | |
| Hepatocellular carcinoma | CHAF1A | Increased tumour size Multifocal tumours Advanced stage Advanced Edmondson–Steiner grade | ↓ OS and DFS | [57] |
| RBBP4 (mRNA) | - | - | [58] | |
| Prostate cancer | CHAF1B | Higher Gleason score Advanced stage | ↓ OS | [59,60] |
| Renal cancer | CHAF1B | Age Histological grade Advanced stage | ↓ OS | [29] |
| Glioblastoma | CHAF1A | Increased tumour size Advanced grade | ↓ OS | [61] |
| CHAF1B | Advanced grade | ↓ OS | [62] | |
| Neuroblastoma | CHAF1A | Advanced stage | ↓ OS and DFS | [65] |
| RBBP4 | Poor differentiation High mitotic index Advanced staging | ↓ OS | [66] | |
| Breast cancer | CHAF1A | High mitotic index Poor histological grade | ↓ DFS | [23,27] |
| CHAF1B | Increased tumour size Poor histological grade High mitotic index | ↓ OS and DFS [23] | [23,28,29] [23,27] | |
| RBBP4 | Lymph node metastasis | ↓ OS | [67] | |
| Melanoma | CHAF1B | Breslow depth Metastasis | ↓ OS and DFS | [68,70] |
| Skin SCC | CHAF1B | Recurrence Metastasis | ↓ OS | [59] |
| Thyroid cancer | RBBP4 | Poor (anaplastic) morphology | - | [71] |
| Salivary gland tumours | CHAF1B | Malignant potential of benign tumours Recurrence Metastasis | - | [59,72] |
| SCC of the tongue | CHAF1A | Advanced stage Recurrence Metastasis | ↓ OS | [73] |
| SCC of the oral cavity | CHAF1B | Advanced stage Recurrence Metastasis | ↓ OS | [73,74,75] |
| SCC of the larynx | CHAF1B | Advanced stage Recurrence Metastasis | ↓ OS | [59] |
| Tumour | CAF-1 Subunit | Marker | Significance | Reference |
|---|---|---|---|---|
| Breast | CHAF1B | Ki-67 | r = 0.97; p < 10−4 | [29] |
| Breast (smear) | CHAF1B | Ki-67 | r = 0.94; p < 10−4 | [28] |
| Breast (smear) | CHAF1B | PCNA | r = 0.95; p = 10−4 | [28] |
| CHAF1A | ASF1B | r = 0.6; p < 10−10 | [23] | |
| CHAF1B | ASF1B | r = 0.7; p < 10−9 | [23] | |
| CHAF1A | Ki-67 | r = 0.69; p < 10−3 | [27] | |
| CHAF1B | Ki-67 | r = 0.65; p < 10−3 | [27] | |
| Endometrial | CHAF1B | Ki-67 | r = 0.98; p < 10−4 | [29] |
| Cervical | CHAF1B | Ki-67 | r = 0.80; p < 10−4 | [29] |
| Prostate | CHAF1B | Ki-67 | r = 0.95; p < 10−4 | [29] |
| Colon | CHAF1B | Ki-67 | r = 0.97; p < 10−4 | [29] |
| Gastric | CHAF1B | Ki-67 | r = 0.89; p < 10−4 | [29] |
| CHAF1B | MCM2 | r = 0.64; p < 10−4 | [28] | |
| CHAF1B | MCM5 | r = 0.49; p < 10−4 | [28] | |
| Pancreatic | CHAF1B | Ki-67 | r = 0.99; p < 10−4 | [29] |
| Renal | CHAF1B | Ki-67 | r = 0.96; p < 10−4 | [29] |
| Thyroid | CHAF1B | Ki-67 | r = 0.86; p < 10−4 | [29] |
| NSCLC | CHAF1B | Ki-67 | r = 0.86; p < 10−4 | [51] |
| SCC (tongue) | CHAF1A | PCNA | Inverse correlation | [73] |
| CHAF1B | PCNA | Correlation | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sykaras, A.G.; Pergaris, A.; Theocharis, S. Challenging, Accurate and Feasible: CAF-1 as a Tumour Proliferation Marker of Diagnostic and Prognostic Value. Cancers 2021, 13, 2575. https://doi.org/10.3390/cancers13112575
Sykaras AG, Pergaris A, Theocharis S. Challenging, Accurate and Feasible: CAF-1 as a Tumour Proliferation Marker of Diagnostic and Prognostic Value. Cancers. 2021; 13(11):2575. https://doi.org/10.3390/cancers13112575
Chicago/Turabian StyleSykaras, Alexandros G., Alexandros Pergaris, and Stamatios Theocharis. 2021. "Challenging, Accurate and Feasible: CAF-1 as a Tumour Proliferation Marker of Diagnostic and Prognostic Value" Cancers 13, no. 11: 2575. https://doi.org/10.3390/cancers13112575