Current Status and Perspectives of Patient-Derived Models for Ewing’s Sarcoma

Division of Rare Cancer Research, National Cancer Center Research Institute, 5-1-1 Tsukiji, Chuo-ku, Tokyo 104-0045, Japan
Cancers 2020, 12(9), 2520; https://doi.org/10.3390/cancers12092520
Submission received: 18 August 2020
/
Revised: 1 September 2020
/
Accepted: 1 September 2020
/
Published: 4 September 2020
(This article belongs to the Special Issue Ewing Sarcoma)
Abstract
:Simple Summary
A paucity of patient-derived cancer models hinders the development of novel therapeutic strategies in Ewing’s sarcoma. This review aimed to demonstrate the utility and possibility of popular patient-derived cancer models by overviewing the reported studies and to encourage the researchers to develop more models for Ewing’s sarcoma.
Abstract
Patient-derived cancer models, including cell lines, organoids, and xenografts, are indispensable tools in cancer research. These models, which recapitulate molecular features of original tumors, allow studies on the biological significance of cancer-associated genes, antitumor effects of novel agents, and molecular mechanisms underlying clinical behaviors of tumors. Moreover, the predictive utility of patient-derived cancer models is expected to facilitate drug development and precision medicine. Ewing’s sarcoma is a highly aggressive mesenchymal tumor with a high metastasis rate. Previous studies demonstrated the utility of cell lines and xenografts in Ewing’s sarcoma research and clinical studies. However, the number of Ewing’s sarcoma models available from public biobanks is limited; this creates an obstacle for research on Ewing’s sarcoma. Novel Ewing’s sarcoma models are needed to establish their utility, further our understanding of the molecular mechanisms, and help develop effective therapeutic strategies. In this review, the current status of patient-derived cancer models is overviewed, and future prospects of model development are discussed from the perspective of Ewing’s sarcoma research. It should be of interest to researchers and clinicians who work on patient-derived cancer models.
{kind=link}
1. Introduction
Ewing’s Sarcoma and Patient-Derived Cancer Models
Ewing’s sarcoma is a highly aggressive mesenchymal tumor with a high metastasis rate in the lungs. Ewing’s sarcoma is the second most common bone tumor in children and adolescents after osteosarcoma, accounting for 3% of all childhood malignancies [1,2]. The genetic predispositions for Ewing’s sarcoma have not yet been established [3], and no preventive measures have been developed for Ewing’s sarcoma [4]. Ewing’s sarcoma is characterized by the presence of chromosome translocation, which causes the chimeric fusions; the t(11;22) chromosomal rearrangement causes the fusion of the EWSR1 gene on chromosome 22 with the FLI-1 gene on chromosome 11. The amino terminus of EWSR1 fuses to the DNA-binding domain of FLI1, and a chimeric transcription factor EWSRI-FLI-1 is generated [5]. The EWSR1-FLI-1 fusion is observed in 85% of Ewing’s sarcoma cases [5,6]. EWSR1-FLI1 plays a crucial role in the carcinogenesis and progression of Ewing’s sarcoma, regulating the expression of multiple genes [7,8]. Comprehensive gene and protein analyses have been conducted to investigate the molecular backgrounds of etiology and progression of Ewing’s sarcoma [9,10,11,12].
The treatment of Ewing’s sarcoma consists of a multimodal approach such as a combination of chemotherapy, surgery, and/or radiotherapy. Treatments for patients with localized Ewing’s sarcoma have significantly progressed over the past 30 years [13,14,15,16,17,18]. However, only 55% of patients receive appropriate chemotherapy [4], and there is much scope for improvement in treatment for patients with recurrence and resistance. Recently, novel therapeutic approaches for Ewing’s sarcoma have been developed based on the understanding of the molecular mechanisms of disease progression. For example, the therapeutic utility of a specific antibody [19], a small-molecule inhibitor [20], and their combination [21] targeting the insulin growth factor system has been examined in pre-clinical studies and clinical trials [22]. Numerous therapeutic agents, including monoclonal antibodies and small compounds, targeting the IGF1R pathway have been developed, and the results of clinical trials were well summarized in the previous review article [22].
Patient-derived cancer models have been critical tools for basic and pre-clinical research since the dawn of cancer research. The patient-derived cancer models, which faithfully capture the characters of original tumors, enable the functional studies for genomic aberrations, the evaluation of the antitumor effects of novel agents, and the validation of hypotheses of molecular mechanisms of diseases. Recent progress of genomics technologies generated a massive amount of clinically annotated genomics data [23,24], identifying mutation-defined subgroups, which could be associated with specific treatments [25,26,27,28]. The clarification of biological significance of genomic findings becomes more important. The clinical significance of genetic mutations depends on the cancer types [29,30,31], being influenced by the lineage and differentiation of tumor precursor cells [32]. Thus, we need cell-type-specific patient-derived cancer models to understand the genetic aberrations.
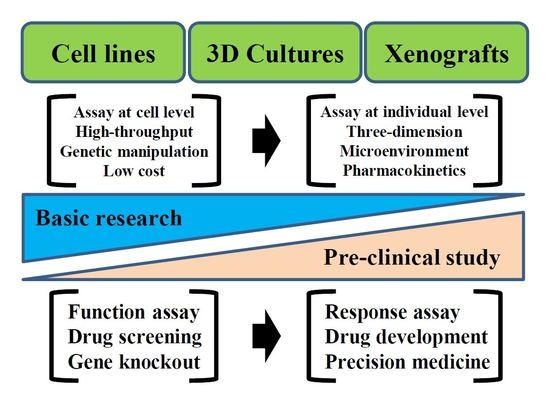
The major patient-derived cancer models are grouped into three types, namely cell lines, organoids, and xenografts, and they, individually, have unique features. Firstly, patient-derived cell lines enable high-throughput drug screening and genome-wide loss-of-function viability experiments [33,34,35,36,37]. Large-scale drug sensitivity screening using cell lines revealed the links between drug sensitivity and genetic alterations, leading to novel indications for anticancer agents [38,39] and discovery of candidate predictive biomarkers [40]. In addition, drug screening and gene function studies can be done at a low cost using cell lines. Numerous cell lines are deposited in public cell banks and are widely available for research use. Beside such advantages, monolayer cell lines lose their three-dimensional architecture and pivotal microenvironment components, such as stroma, vascular structure, and inflammatory cells. In addition, alterations in the genetic background, which affect drug responses, continuously occur during the propagation of cells because of deficient mechanisms of genome stabilization [41]. Moreover, the success rate of cell line establishment varies among cancer types, and tumor cells that are highly selected under survival pressure in tissue culture conditions are subjected to subsequent in vitro studies. Secondly, patient-derived organoids retain the three-dimensional structures of original tumor tissues; hence, they may represent the phenotypic features of original tumors better than the conventional tissue culture monolayer cell lines [42,43,44,45,46]. Furthermore, patient-derived organoids contain cancer stem cells or progenitor cells that are surrounded by microenvironment components. Moreover, the high take rate makes organoids more applicable for clinical practice than other cancer models. Organoids are expected to faithfully recapitulate the characters of original tumors, and have been used in studies of colorectal [47], gastric [48], pancreatic [49], breast [50], bladder [51], prostate [52], and kidney [53] cancers and glioblastoma [54]. However, organoids lack immune components, and organoids of only a limited type of cancers are available from public biobanks. Thirdly, patient-derived xenografts (PDXs) using tumor tissues have several unique advantages over other models. They retain microenvironmental components, such as stromal cells, immune cells, and vascular structure [55,56]. Moreover, PDXs contain the heterogeneous components of the original tumor tissue, suggesting their potentials for predictive modalities [57,58,59,60,61]. In addition to tumor tissues, circulating tumor cells (CTCs) are also promising sources of PDXs. CTCs were reported in patients with Ewing’s sarcoma, and their clinical utilities have been investigated using PDXs [62,63]. The biological behaviors and the metastatic potentials of CTCs can be evaluated by PDXs, and the anti-metastasis effects of drugs will be assessed using PDXs with CTCs. PDXs have been used for several decades in oncology, with the aim to develop clinical applications [64]. Besides their wide use, recent studies have indicated that the genomic contents of PDXs change during passaging in a way that does not resemble the progression of the original tumor; genomic evolution due to genetic instability is generally an issue in cancer models [65,66]. Moreover, human stromal components are gradually replaced with those of the experimental animals during passaging [67,68,69,70,71]. Because the genomic profile and tumor microenvironment affect the response to treatments, such alterations can reduce the utility of PDXs. These models have both pros and cons and complement the use of other patient-derived cancer models.
The purpose of this review article is to provide an overview of the current status of patient-derived models of Ewing’s sarcoma. The necessity of additional models for further insights into the etiology and progression of Ewing’s sarcoma is discussed. This review should be of interest to a wide range of researchers and clinicians who use patient-derived cancer models in their work.
2. Materials and Methods
The previous studies for Ewing’s sarcoma, where patient-derived cancer models were implemented, were investigated by searching the PubMed database (https://pubmed.ncbi.nlm.nih.gov/) using the keywords of Ewing’s sarcoma and the individual model names. After reviewing the search results, the papers with which the author was impressed in terms of novelty and sound results, and the papers demonstrating the possibilities of patient-derived cancer models, were selected. The process of paper selection depended on the author’s experience in the field of sarcoma research and patient-derived cancer models.
3. Results: Technical Variation of Patient-Derived Cancer Models Used for Research on Ewing’s Sarcoma
3.1. Cell Lines
Experiments using patient-derived cells provided novel insights into the molecular mechanisms of the etiology and progression of Ewing’s sarcoma. Several illustrative examples of the utility of cell lines in functional studies of the transcription factor EWSR1-FLI1 have been demonstrated. For example, Boulay et al. [72] reported that in in vitro tissue culture cells, EWSR1-FLI1 interacts with the Brahma homologue-related gene 1/ Brahma homologue-associated factor (BAF) chromatin remodeling complex and activates target gene transcription. Erkizan et al. [73] reported the interaction of EWSR1-FLI1 with multiple proteins in cell lines using mass spectrometry and demonstrated that RNA-helicase A could be a therapeutic target for Ewing’s sarcoma. Genome-wide gene knockdown assays using RNAi and cell lines revealed the function of EWSR1-FLI1. He et al. [74] reported that RNA interference (RNAi) screening targeting 6781 genes resulted in the identification of leucine-rich repeats and tryptophan-aspartic acid-repeat domain containing 1 (LRWD1) as a regulator of EWS-FLI1-driven cell viability. LRWD1-regulated transcriptional activity of EWSR1-FLI1 is associated with poor prognosis of patients with Ewing’s sarcoma. Another example of the utility of cell lines is the screening of oncology drugs. By screening a drug library, olaparib was identified to have remarkable in vitro activity against Ewing’s sarcoma [40,75], and these results were reproduced by other researchers [76,77,78]. The efficacy of olaparib was evaluated in phase II clinical trials [79]. Although the objective responses were not observed, the utility of patient-derived cancer models was proven in this case. Patient-derived cancer cells have also contributed to the study of the molecular mechanisms underlying disease progression. Ewing’s sarcoma tumor tissues include multiple types of tumor cells with different expression levels of EWSR1-FLI1. Franzetti et al. [80] followed up on these interesting observations by investigating the heterogeneous expression level of EWSR1-FLI1 using single cell reverse transcription polymerase chain reaction (RT-PCR) and imaging analysis, and demonstrated that EWSR1-FLI1 expression promoted lung metastasis [80]. Additional examples of the utility of cell lines in the study of Ewing’s sarcoma are too numerous to discuss here. Although some aspects of cellular physiology and heterogeneity may be inadequately retained in the cell lines, none of these studies could be performed if patient-derived cancer cells were not available for Ewing’s sarcoma.
Despite their substantial utility, cell lines for Ewing’s sarcoma are rarely available from public cell banks. According to a search of the world’s largest cell line database, Cellosaurus [81], there are only nine Ewing’s sarcoma cell lines deposited in cell banks. This is partly due to the number of patients with Ewing’s sarcoma being substantially small; the overall incidence of Ewing’s sarcoma averages 2.93 cases per million annually [14]. The number of cell lines needed to examine the antitumor effects of agents to generate enough data to promote further studies is unclear. In phase II clinical trials, where the efficacy of drugs is evaluated before the following phase III study, several dozens of patients need to be enrolled [82]. Although the absolute number of patients required for clinical trials with statistical power can be influenced by many factors, such as the expected efficacy of the drugs, prior probability of a favorable response, heterogeneity of the patient population, and the endpoint of the clinical trial, it is also desirable to include an equivalent number of cell lines from different patients in the preclinical study. In addition to the number of cell lines, cell lines with improved quality are needed. Most of the existing cell lines have inherent problems; generally, they have been passaged for many years, or even decades, and they are not adequately annotated with clinical and pathological data [83]. In addition, the discrepancy in results obtained from cell lines and those from clinical trials may be attributable to the instability of the genomic background of the cell lines and accumulation of mutations during multiple passaging [41]. Thus, after long-term culture, the cells should not be used for drug response assays. Recent studies indicated that short-term cultured cells present with features that are more similar to those of original tumors than long-term cultured cells, and thus are more useful for drug screening in both two- and three-dimensional culture conditions [37,44].
3.2. Organoids
Previous studies have indicated the potential utility of organoids for basic and clinical studies [42]. A literature search of PubMed using the keywords “sarcoma” and “organoid” revealed no reports on the study of sarcomas using organoids. This may be owing to the difficulty in establishing organoids using sarcoma tumor tissues, for unknown reasons. Alternatively, it may be simply because of the difficulty in obtaining adequate tumor tissues for experiments. If mesenchymal tumors are generally refractory to organoid formation, the investigation of mechanisms underlying such unique characteristics may lead to elucidation of novel biological aspects of organoids, or development of methodologies for establishing organoids.
3.3. PDXs
Previous studies employed and characterized PDXs of Ewing’s sarcoma. Nanni et al. [84] studied bone malignancies, including Ewing’s sarcoma. They implanted tumor tissues from 29 patients with Ewing’s sarcoma into mice and obtained PDXs in 7 of the 29 cases (24%). Gene expression analysis revealed that the expression level of all examined genes was not significantly different between PDX and tumor tissues. Patient-derived orthotopic xenografts (PDOXs) were also used in the study of Ewing’s sarcoma. The majority of subcutaneously implanted tumor tissues do not metastasize in immunocompromised mice; in contrast, the orthotopically implanted tumor tissues lead to metastasis [85]. Thus, PDOXs may better mimic the propensities of original tumors and may be more suitable to cancer research than the conventional PDXs. Miyake et al. [86] used PDOXs to demonstrate that treatment with regorafenib was effective against doxorubicin-resistant Ewing’s sarcoma. Miyake et al. [86] also demonstrated that the combination of temozolomide and irinote can inhibit the growth of a doxorubicin-resistant PDOX of recurrent Ewing’s sarcoma [87]. The antitumor effects of palbociclib, a CDK4/6 inhibitor, and listinib, an inhibitor of insulin receptor and insulin growth factor receptors, were examined using PDOXs of Ewing’s sarcoma with Fusion involved in t(12;16) in malignant liposarcoma-ETS-related gene fusion and cyclin-dependent kinase inhibitor 2A/B loss [88]. The antitumor effects of unique antitumor agents were also examined in PDOXs of Ewing’s sarcoma. Methionine addiction is a common feature of cancer, and methionine restriction causes cell cycle arrest. Thus, methionine is a therapeutic target in methionine-dependent cancers. Recombinant methionase (rMETase) can restrict the metabolism of methionine, and the antitumor effects of rMETase were reported in PDOXs of Ewing’s sarcoma [89]. Salmonella sp., a facultative anaerobe, preferentially replicates and colonizes within tumors when injected from a distal site, and this leads to tumor regression [90]. Salmonella typhimurium A1-R is a strain with high tumor colonization and antitumor efficacy [91]. Miyake et al. [92] demonstrated that combination treatments with rMETase and Salmonella typhimurium A1-R had significantly more prominent antitumor effects in PDOXs of Ewing’s sarcoma, compared with monotherapy with each agen. These reports demonstrated that PDXs and PDOXs can facilitate drug development and serve as tools of pre-clinical study.
The feasibility of using PDXs for precision medicine was suggested in a previous study. Stebbing et al. [93] reported how they used PDXs for precision medicine in Ewing’s sarcoma. In their study, the tumor tissues from three patients with Ewing’s sarcoma were examined, and data were obtained from two of the three. The effects of chemotherapy were compared between the PDXs and the tumors from which they established PDX in one case. A combination of docetaxel, gemcitabine, and bevacizumab proved effective in the PDXs. Based on the results of this experiment, the patient was treated with the same combination of anticancer drugs. A partial response was observed after two cycles of treatment, and after an additional five treatment cycles, the beneficial effects continued for more than six months [93]. These observations suggested how we can use PDXs for drug development and precision cancer medicine. Despite these promising observations and expectations for clinical applications, according to a search of the PDX database PDXFinder (https://www.pdxfinder.org/), there are only five PDXs of Ewing’s sarcoma that are currently available from public biobanks. Clearly, more PDXs are needed for preclinical studies.
4. Conclusions
Future Perspectives of Patient-Derived Ewing’s Sarcoma Models
This review presented several examples of the considerable possibilities for using patient-derived cancer models. The examples demonstrated in this review showed that adequate models can facilitate research considerably. Although the patient-derived cancer models have versatile utilities, each model has its own pros and cons, and we should use different models depending on the situation. Because of the limited number of patients, such models are seriously difficult to obtain in Ewing’s sarcoma. In other words, the delay in effective treatments in patients with Ewing’s sarcoma is partially attributable to this paucity of adequate cancer models. Indeed, since this disease was defined about one century ago, only a limited number and variation of models were developed. Tumor tissues for Ewing’s sarcoma research are limited owing to the exceptionally low incidence and prevalence of this type of cancer, and the researchers who can access such models have significant advantages over other researchers who cannot, and at the same time, they have a responsibility to use them in a proper way. The establishment of cancer models should not be performed by researchers in a competitive way; rather, cancer models should be shared as public resources and made available to subsequent generations.
Funding
This research was supported by the Japan Agency for Medical Research and Development grant 20ck0106537h0001, “Study to Overcome the Limits of Cancer Genome-Based Medicine Using Patient-Derived ‘Rare Cancer’ Model”.
Conflicts of Interest
The author declares no conflict of interest. The sponsors had no role in the design, execution, interpretation, or writing of the study.
References
- Grier, H.E. The Ewing Family of Tumors. Pediatr. Clin. N. Am. 1997, 44, 991–1004. [Google Scholar] [CrossRef]
- Rodriguez-Galindo, C.; Liu, T.; Krasin, M.J.; Wu, J.; Billups, C.A.; Daw, N.C.; Spunt, S.L.; Rao, B.N.; Santana, V.M.; Navid, F. Analysis of prognostic factors in Ewing sarcoma family of tumors. Cancer 2007, 110, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.L.; Lessnick, S.L.; Jones, K.B.; Gouw, L.G.; Cummings, J.E.; Albright, L.C.; Schiffman, J.D. Is There a Predisposition Gene for Ewing’s Sarcoma? J. Oncol. 2010, 2010, 397632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamuth, N.J.; Womer, R.B. Ewing’s sarcoma. Lancet Oncol. 2010, 11, 184–192. [Google Scholar] [CrossRef]
- Delattre, O.; Zucman-Rossi, J.; Plougastel-Douglas, B.; Desmaze, C.; Melot, T.; Peter, M.; Kovar, H.; Joubert, I.; De Jong, P.; Rouleau, G.; et al. Gene fusion with an ETS DNA-binding domain caused by chromosome translocation in human tumours. Nature 1992, 359, 162–165. [Google Scholar] [CrossRef]
- Dubois, S.G.; Grier, H.E.; Lessnick, S.L. Ewing’s Sarcoma. In Oncology of Infancy and Childhood, 1st ed.; Saunders: Philadelphia, PA, USA, 2009; pp. 829–869. [Google Scholar]
- Üren, A.; Toretsky, J.A. Ewing’s sarcoma oncoprotein EWS–FLI1: The perfect target without a therapeutic agent. Futur. Oncol. 2005, 1, 521–528. [Google Scholar] [CrossRef]
- Staege, M.S.; Hutter, C.; Neumann, I.; Foja, S.; Hattenhorst, U.E.; Hansen, G.; Afar, D.; Burdach, S.E.G. DNA Microarrays Reveal Relationship of Ewing Family Tumors to Both Endothelial and Fetal Neural Crest-Derived Cells and Define Novel Targets. Cancer Res. 2004, 64, 8213–8221. [Google Scholar] [CrossRef] [Green Version]
- Sheffield, N.C.; Pierron, G.; Klughammer, J.; Datlinger, P.; Schönegger, A.; Schuster, M.; Hadler, J.; Surdez, D.; Guillemot, D.; Lapouble, E.; et al. DNA methylation heterogeneity defines a disease spectrum in Ewing sarcoma. Nat. Med. 2017, 23, 386–395. [Google Scholar] [CrossRef]
- Brohl, A.S.; Solomon, D.A.; Chang, W.; Wang, J.; Song, Y.; Sindiri, S.; Patidar, R.; Hurd, L.; Chen, L.; Shern, J.F.; et al. The Genomic Landscape of the Ewing Sarcoma Family of Tumors Reveals Recurrent STAG2 Mutation. PLoS Genet. 2014, 10, e1004475. [Google Scholar] [CrossRef] [Green Version]
- Crompton, B.D.; Stewart, C.; Taylor-Weiner, A.; Alexe, G.; Kurek, K.C.; Calicchio, M.L.; Kiezun, A.; Carter, S.L.; Shukla, S.A.; Mehta, S.S.; et al. The Genomic Landscape of Pediatric Ewing Sarcoma. Cancer Discov. 2014, 4, 1326–1341. [Google Scholar] [CrossRef] [Green Version]
- Tirode, F.; Surdez, D.; Ma, X.; Parker, M.D.; Le Deley, M.-C.; Bahrami, A.; Zhang, Z.; Lapouble, E.; Grossetête-Lalami, S.; Rusch, M.; et al. Genomic landscape of Ewing sarcoma defines an aggressive subtype with co-association of STAG2 and TP53 mutations. Cancer Discov. 2014, 4, 1342–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damron, T.; Ward, W.G.; Stewart, A. Osteosarcoma, Chondrosarcoma, and Ewing Sarcoma. Clin. Orthop. Relat. Res. 2007, 459, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Esiashvili, N.; Goodman, M.; Marcus, R.B. Changes in Incidence and Survival of Ewing Sarcoma Patients Over the Past 3 Decades. J. Pediatr. Hematol. 2008, 30, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Cotterill, S.; Ahrens, S.; Paulussen, M.; Jürgens, H.; Voûte, P.; Gadner, H.; Craft, A. Prognostic Factors in Ewing’s Tumor of Bone: Analysis of 975 Patients From the European Intergroup Cooperative Ewing’s Sarcoma Study Group. J. Clin. Oncol. 2000, 18, 3108–3114. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, S.; Hoffmann, C.; Jabar, S.; Braun-Munzinger, G.; Paulussen, M.; Dunst, J.; Rübe, C.; Winkelmann, W.; Heinecke, A.; Göbel, U.; et al. Evaluation of prognostic factors in a tumor volume-adapted treatment strategy for localized Ewing sarcoma of bone: The CESS 86 experience. Med Pediatr. Oncol. 1999, 32, 186–195. [Google Scholar] [CrossRef]
- Gaspar, N.; Hawkins, D.S.; Dirksen, U.; Lewis, I.J.; Ferrari, S.; Le Deley, M.-C.; Kovar, H.; Grimer, R.J.; Whelan, J.; Claude, L.; et al. Ewing Sarcoma: Current Management and Future Approaches Through Collaboration. J. Clin. Oncol. 2015, 33, 3036–3046. [Google Scholar] [CrossRef]
- Werier, J.; Yao, X.; Caudrelier, J.-M.; Di Primio, G.; Ghert, M.; Gupta, A.A.; Kandel, R.; Verma, S. A systematic review of optimal treatment strategies for localized Ewing’s sarcoma of bone after neo-adjuvant chemotherapy. Surg. Oncol. 2016, 25, 16–23. [Google Scholar] [CrossRef]
- Olmos, D.; Postel-Vinay, S.; Molife, L.R.; Okuno, S.H.; Schuetze, S.M.; Paccagnella, M.L.; Batzel, G.N.; Yin, D.; Pritchard-Jones, K.; Judson, I.; et al. Safety, pharmacokinetics, and preliminary activity of the anti-IGF-1R antibody figitumumab (CP-751,871) in patients with sarcoma and Ewing’s sarcoma: A phase 1 expansion cohort study. Lancet Oncol. 2010, 11, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Manara, M.C.; Landuzzi, L.; Nanni, P.; Nicoletti, G.; Zambelli, D.; Lollini, P.L.; Nanni, C.; Hofmann, F.; Garcia-Echeverria, C.; Picci, P.; et al. Preclinical In vivo Study of New Insulin-Like Growth Factor-I Receptor-Specific Inhibitor in Ewing’s Sarcoma. Clin. Cancer Res. 2007, 13, 1322–1330. [Google Scholar] [CrossRef] [Green Version]
- Benini, S.; Manara, M.C.; Baldini, N.; Cerisano, V.; Serra, M.; Mercuri, M.; Lollini, P.L.; Nanni, P.; Picci, P.; Scotlandi, K. Inhibition of insulin-like growth factor I receptor increases the antitumor activity of doxorubicin and vincristine against Ewing’s sarcoma cells. Clin. Cancer Res. 2001, 7, 1790–1797. [Google Scholar]
- Mancarella, C.; Scotlandi, K. 40 YEARS OF IGF1: IGF system in sarcomas: A crucial pathway with many unknowns to exploit for therapy. J. Mol. Endocrinol. 2018, 61, T45–T60. [Google Scholar] [CrossRef] [PubMed]
- AACR Project GENIE Consortium. The AACR Project GENIE Consortium AACR Project GENIE: Powering Precision Medicine through an International Consortium. Cancer Discov. 2017, 7, 818–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsello, S.M.; Bittker, J.A.; Liu, Z.; Gould, J.; McCarren, P.; Hirschman, J.E.; Johnston, S.E.; Vrcic, A.; Wong, B.; Khan, M.; et al. The Drug Repurposing Hub: A next-generation drug library and information resource. Nat. Med. 2017, 23, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Hudson, T.J.; Anderson, W.; Artez, A.; Barker, A.D.; Bell, C.; Bernabe, R.R.; Bhan, M.K.; Calvo, F.; Eerola, I.; Gerhard, D.S.; et al. The International Cancer Genome Consortium International network of cancer genome projects. Nature 2010, 464, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Research Network. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Levine, D.A. The Cancer Genome Atlas Research Network: Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature 2013, 499, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Tsimberidou, A.; Wen, S.; Hong, D.S.; Wheler, J.J.; Falchook, G.S.; Fu, S.; Piha-Paul, S.; Naing, A.; Janku, F.; Aldape, K.; et al. Personalized medicine for patients with advanced cancer in the phase I program at MD Anderson: Validation and landmark analyses. Clin. Cancer Res. 2014, 20, 4827–4836. [Google Scholar] [CrossRef] [Green Version]
- André, F.; Bachelot, T.; Commo, F.; Campone, M.; Arnedos, M.; Dieras, V.; Lacroix-Triki, M.; Lacroix, L.; Cohen, P.; Gentien, D.; et al. Comparative genomic hybridisation array and DNA sequencing to direct treatment of metastatic breast cancer: A multicentre, prospective trial (SAFIR01/UNICANCER). Lancet Oncol. 2014, 15, 267–274. [Google Scholar] [CrossRef]
- Vansteenkiste, J.; Canon, J.-L.; De Braud, F.; Grossi, F.; De Pas, T.; Gray, J.E.; Su, W.-C.; Felip, E.; Yoshioka, H.; Gridelli, C.; et al. Safety and Efficacy of Buparlisib (BKM120) in Patients with PI3K Pathway-Activated Non-Small Cell Lung Cancer: Results from the Phase II BASALT-1 Study. J. Thorac. Oncol. 2015, 10, 1319–1327. [Google Scholar] [CrossRef] [Green Version]
- Garraway, L.A.; Sellers, W.R. Lineage dependency and lineage-survival oncogenes in human cancer. Nat. Rev. Cancer 2006, 6, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.V.; Haber, D.A.; Settleman, J. Cell line-based platforms to evaluate the therapeutic efficacy of candidate anticancer agents. Nat. Rev. Cancer 2010, 10, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Goodspeed, A.; Heiser, L.M.; Gray, J.W.; Costello, J.C. Tumor-Derived Cell Lines as Molecular Models of Cancer Pharmacogenomics. Mol. Cancer Res. 2015, 14, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564–576.e16. [Google Scholar] [CrossRef] [Green Version]
- McMillan, E.A.; Ryu, M.-J.; Diep, C.H.; Mendiratta, S.; Clemenceau, J.R.; Vaden, R.M.; Kim, J.-H.; Motoyaji, T.; Covington, K.R.; Peyton, M.; et al. Chemistry-First Approach for Nomination of Personalized Treatment in Lung Cancer. Cell 2018, 173, 864–878.e29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-K.; Liu, Z.; Sa, J.K.; Shin, S.; Wang, J.; Bordyuh, M.; Cho, H.J.; Elliott, O.; Chu, T.; Choi, S.W.; et al. Pharmacogenomic landscape of patient-derived tumor cells informs precision oncology therapy. Nat. Genet. 2018, 50, 1399–1411. [Google Scholar] [CrossRef]
- Iorio, F.; Knijnenburg, T.A.; Vis, D.J.; Bignell, G.R.; Menden, M.P.; Schubert, M.; Aben, N.; Gonçalves, E.; Barthorpe, S.; Lightfoot, H.; et al. A Landscape of Pharmacogenomic Interactions in Cancer. Cell 2016, 166, 740–754. [Google Scholar] [CrossRef] [Green Version]
- Holbeck, S.L.; Collins, J.M.; Doroshow, J.H. Analysis of Food and Drug Administration-approved anticancer agents in the NCI60 panel of human tumor cell lines. Mol. Cancer Ther. 2010, 9, 1451–1460. [Google Scholar] [CrossRef] [Green Version]
- Garnett, M.J.; Edelman, E.J.; Heidorn, S.J.; Greenman, C.D.; Dastur, A.; Lau, K.W.; Greninger, P.; Thompson, I.R.; Luo, X.; Soares, J.; et al. Systematic identification of genomic markers of drug sensitivity in cancer cells. Nature 2012, 483, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef]
- Tiriac, H.; Belleau, P.; Engle, D.D.; Plenker, D.; Deschênes, A.; Somerville, T.D.D.; Froeling, F.E.M.; Burkhart, R.A.; Denroche, R.E.; Jang, G.H.; et al. Organoid Profiling Identifies Common Responders to Chemotherapy in Pancreatic Cancer. Cancer Discov. 2018, 8, 1112–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, S.J.; Decker, B.; Roberts, E.; Horowitz, N.S.; Muto, M.G.; Worley, M.J.; Feltmate, C.M.; Nucci, M.R.; Swisher, E.M.; Nguyen, H.; et al. Prediction of DNA Repair Inhibitor Response in Short-Term Patient-Derived Ovarian Cancer Organoids. Cancer Discov. 2018, 8, 1404–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, J.T.; Li, X.; Zhu, J.; Giangarra, V.; Grzeskowiak, C.L.; Ju, J.; Liu, I.H.; Chiou, S.-H.; Salahudeen, A.A.; Smith, A.R.; et al. Organoid Modeling of the Tumor Immune Microenvironment. Cell 2018, 175, 1972–1988.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuveson, D.A.; Clevers, H. Cancer modeling meets human organoid technology. Science 2019, 364, 952–955. [Google Scholar] [CrossRef]
- Van De Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; Van Houdt, W.; Van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.H.N.; Siu, H.C.; Law, S.; Ho, S.L.; Yue, S.S.; Tsui, W.Y.; Chan, D.; Chan, A.S.; Ma, S.K.Y.; Lam, K.O.; et al. A Comprehensive Human Gastric Cancer Organoid Biobank Captures Tumor Subtype Heterogeneity and Enables Therapeutic Screening. Cell Stem Cell 2018, 23, 882–897.e11. [Google Scholar] [CrossRef] [Green Version]
- Seino, T.; Kawasaki, S.; Shimokawa, M.; Tamagawa, H.; Toshimitsu, K.; Fujii, M.; Ohta, Y.; Matano, M.; Nanki, K.; Kawasaki, K.; et al. Human Pancreatic Tumor Organoids Reveal Loss of Stem Cell Niche Factor Dependence during Disease Progression. Cell Stem Cell 2018, 22, 454–467.e6. [Google Scholar] [CrossRef] [Green Version]
- Sachs, N.; De Ligt, J.; Kopper, O.; Gogola, E.; Bounova, G.; Weeber, F.; Balgobind, A.V.; Wind, K.; Gracanin, A.; Begthel, H.; et al. A Living Biobank of Breast Cancer Organoids Captures Disease Heterogeneity. Cell 2018, 172, 373–386.e10. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Hu, W.; Matulay, J.T.; Silva, M.V.; Owczarek, T.B.; Kim, K.; Chua, C.-W.; Barlow, L.J.; Kandoth, C.; Williams, A.B.; et al. Tumor Evolution and Drug Response in Patient-Derived Organoid Models of Bladder Cancer. Cell 2018, 173, 515–528.e17. [Google Scholar] [CrossRef] [Green Version]
- Beshiri, M.L.; Tice, C.M.; Tran, C.; Nguyen, H.M.; Sowalsky, A.G.; Agarwal, S.; Jansson, K.H.; Yang, Q.; McGowen, K.M.; Yin, J.J.; et al. A PDX/Organoid Biobank of Advanced Prostate Cancers Captures Genomic and Phenotypic Heterogeneity for Disease Modeling and Therapeutic Screening. Clin. Cancer Res. 2018, 24, 4332–4345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calandrini, C.; Schutgens, F.; Oka, R.; Margaritis, T.; Candelli, T.; Mathijsen, L.; Ammerlaan, C.; Van Ineveld, R.L.; Derakhshan, S.; De Haan, S.; et al. An organoid biobank for childhood kidney cancers that captures disease and tissue heterogeneity. Nat. Commun. 2020, 11, 1310. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Salinas, R.D.; Zhang, D.Y.; Nguyen, P.T.; Schnoll, J.G.; Wong, S.Z.H.; Thokala, R.; Sheikh, S.; Saxena, D.; Prokop, S.; et al. A Patient-Derived Glioblastoma Organoid Model and Biobank Recapitulates Inter- and Intra-tumoral Heterogeneity. Cell 2020, 180, 188–204.e22. [Google Scholar] [CrossRef] [PubMed]
- Calles, A.; Rubio-Viqueira, B.; Hidalgo, M. Primary Human Non-Small Cell Lung and Pancreatic Tumorgraft Models-Utility and Applications in Drug Discovery and Tumor Biology. Curr. Protoc. Pharmacol. 2013, 61, 14.26.1–14.26.21. [Google Scholar] [CrossRef]
- Tentler, J.J.; Tan, A.C.; Weekes, C.D.; Jimeno, A.; Leong, S.; Pitts, T.M.; Arcaroli, J.J.; Messersmith, W.A.; Eckhardt, S.G. Patient-derived tumor xenografts as models for oncology drug development. Nat. Rev. Clin. Oncol. 2012, 9, 338–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, M.; Amant, F.; Biankin, A.V.; Budinská, E.; Byrne, A.; Caldas, C.; Clarke, R.; De Jong, S.; Jonkers, J.; Mælandsmo, G.M.; et al. Patient-Derived Xenograft Models: An Emerging Platform for Translational Cancer Research. Cancer Discov. 2014, 4, 998–1013. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Korn, J.M.; Ferretti, S.; Monahan, J.E.; Wang, Y.; Singh, M.; Zhang, C.; Schnell, C.; Yang, G.; Zhang, Y.; et al. High-throughput screening using patient-derived tumor xenografts to predict clinical trial drug response. Nat. Med. 2015, 21, 1318–1325. [Google Scholar] [CrossRef]
- Townsend, E.C.; Murakami, M.A.; Christodoulou, A.; Christie, A.L.; Köster, J.; DeSouza, T.A.; Morgan, E.A.; Kallgren, S.P.; Liu, H.; Wu, S.-C.; et al. The Public Repository of Xenografts (ProXe) enables discovery and randomized phase II-like trials in mice. Cancer Cell 2016, 29, 574–586. [Google Scholar] [CrossRef]
- Bruna, A.; Rueda, O.M.; Greenwood, W.; Batra, A.S.; Callari, M.; Batra, R.N.; Pogrebniak, K.; Sandoval, J.L.; Cassidy, J.W.; Vidakovic, A.T.; et al. A Biobank of Breast Cancer Explants with Preserved Intra-tumor Heterogeneity to Screen Anticancer Compounds. Cell 2016, 167, 260–274.e22. [Google Scholar] [CrossRef] [Green Version]
- Brait, M.; Izumchenko, E.; Kagohara, L.T.; Long, S.; Wysocki, P.T.; Faherty, B.; Fertig, E.; Khor, T.O.; Bruckheimer, E.; Baia, G.; et al. Comparative mutational landscape analysis of patient-derived tumour xenografts. Br. J. Cancer 2017, 116, 515–523. [Google Scholar] [CrossRef]
- Benini, S.; Gamberi, G.; Cocchi, S.; Garbetta, J.; Alberti, L.; Righi, A.; Gambarotti, M.; Picci, P.; Ferrari, S. Detection of circulating tumor cells in liquid biopsy from Ewing sarcoma patients. Cancer Manag. Res. 2018, 10, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Zhu, P.; Mccarty, G.; Meyer, C.F.; Pratilas, C.A.; Levin, A.; Morris, C.D.; Albert, C.M.; Jackson, K.W.; Tang, C.-M.; et al. Size-based detection of sarcoma circulating tumor cells and cell clusters. Oncotarget 2017, 8, 78965–78977. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, H.H.; Neumann, H.A.; Henß, H.; Koch, H.; Kaiser, D.; Arnold, H. Development of Three Human Small Cell Lung Cancer Models in Nude Mice. Adv. Struct. Saf. Stud. 1985, 97, 77–86. [Google Scholar] [CrossRef]
- Ben-David, U.; Ha, G.; Tseng, Y.-Y.; Greenwald, N.F.; Oh, C.; Shih, J.; McFarland, J.M.; Wong, B.; Boehm, J.S.; Beroukhim, R.; et al. Patient-derived xenografts undergo mouse-specific tumor evolution. Nat. Genet. 2017, 49, 1567–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-David, U.; Beroukhim, R.; Golub, T.R. Genomic evolution of cancer models: Perils and opportunities. Nat. Rev. Cancer 2018, 19, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Derose, Y.S.; Wang, G.; Lin, Y.-C.; Bernard, P.S.; Buys, S.S.; Ebbert, M.T.W.; Factor, R.; Matsen, C.B.; Milash, B.A.; Nelson, E.; et al. Tumor grafts derived from women with breast cancer authentically reflect tumor pathology, growth, metastasis and disease outcomes. Nat. Med. 2011, 17, 1514–1520. [Google Scholar] [CrossRef]
- Cassidy, J.W.; Caldas, C.; Bruna, A. Maintaining Tumor Heterogeneity in Patient-Derived Tumor Xenografts. Cancer Res. 2015, 75, 2963–2968. [Google Scholar] [CrossRef] [Green Version]
- Hylander, B.L.; Punt, N.; Tang, H.; Hillman, J.; Vaughan, M.; Bshara, W.; Pitoniak, R.; Repasky, E.A. Origin of the vasculature supporting growth of primary patient tumor xenografts. J. Transl. Med. 2013, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Pirollo, K.F.; Tang, W.-H.; Rait, A.; Chang, E.H. Transferrin-Liposome-Mediated Systemic p53 Gene Therapy in Combination with Radiation Results in Regression of Human Head and Neck Cancer Xenografts. Hum. Gene Ther. 1999, 10, 2941–2952. [Google Scholar] [CrossRef]
- Sanz, L.; Cuesta, A.M.; Salas, C.; Corbacho, C.; Bellas, C.; Álvarez-Vallina, L. Differential transplantability of human endothelial cells in colorectal cancer and renal cell carcinoma primary xenografts. Lab. Investig. 2008, 89, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Boulay, G.; Sandoval, G.J.; Riggi, N.; Iyer, S.; Buisson, R.; Naigles, B.; Awad, M.E.; Rengarajan, S.; Volorio, A.; McBride, M.; et al. Cancer-Specific Retargeting of BAF Complexes by a Prion-like Domain. Cell 2017, 171, 163–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkizan, H.V.; Kong, Y.; Merchant, M.; Schlottmann, S.; Barber-Rotenberg, J.S.; Yuan, L.; Abaan, O.D.; Chou, T.-H.; Dakshanamurthy, S.; Brown, M.L.; et al. A small molecule blocking oncogenic protein EWS-FLI1 interaction with RNA helicase A inhibits growth of Ewing’s sarcoma. Nat. Med. 2009, 15, 750–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.; Surdez, D.; Rantala, J.K.; Haapa-Paananen, S.; Ban, J.; Kauer, M.; Tomazou, E.M.; Fey, V.; Alonso, J.; Kovar, H.; et al. High-throughput RNAi screen in Ewing sarcoma cells identifies leucine rich repeats and WD repeat domain containing 1 (LRWD1) as a regulator of EWS-FLI1 driven cell viability. Gene 2017, 596, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Brenner, J.C.; Feng, F.Y.; Han, S.; Patel, S.; Goyal, S.V.; Bou-Maroun, L.M.; Liu, M.; Lonigro, R.; Prensner, J.R.; Tomlins, S.A.; et al. PARP-1 inhibition as a targeted strategy to treat Ewing’s sarcoma. Cancer Res. 2012, 72, 1608–1613. [Google Scholar] [CrossRef] [Green Version]
- Stewart, E.; Goshorn, R.; Bradley, C.; Griffiths, L.M.; Benavente, C.A.; Twarog, N.R.; Miller, G.M.; Caufield, W.V.; Freeman, B.B.; Bahrami, A.; et al. Targeting the DNA repair pathway in Ewing sarcoma. Cell Rep. 2014, 9, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Engert, F.; Schneider, C.; Weiss, L.M.; Probst, M.; Fulda, S. PARP Inhibitors Sensitize Ewing Sarcoma Cells to Temozolomide-Induced Apoptosis via the Mitochondrial Pathway. Mol. Cancer Ther. 2015, 14, 2818–2830. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-J.; Yoon, C.; Schmidt, B.; Park, J.; Zhang, A.Y.; Erkizan, H.V.; Toretsky, J.A.; Kirsch, D.G.; Yoon, S.S. Combining PARP-1 inhibition and radiation in Ewing sarcoma results in lethal DNA damage. Mol. Cancer Ther. 2013, 12, 2591–2600. [Google Scholar] [CrossRef] [Green Version]
- Choy, E.; Butrynski, J.E.; Harmon, D.C.; Morgan, J.A.; George, S.; Wagner, A.J.; D’Adamo, D.; Cote, G.M.; Flamand, Y.; Benes, C.H.; et al. Phase II study of olaparib in patients with refractory Ewing sarcoma following failure of standard chemotherapy. BMC Cancer 2014, 14, 813. [Google Scholar] [CrossRef] [Green Version]
- Franzetti, G.-A.; Laud-Duval, K.; Van Der Ent, W.; Brisac, A.; Irondelle, M.; Aubert, S.; Dirksen, U.; Bouvier, C.; De Pinieux, G.; Snaar-Jagalska, E.; et al. Cell-to-cell heterogeneity of EWSR1-FLI1 activity determines proliferation/migration choices in Ewing sarcoma cells. Oncogene 2017, 36, 3505–3514. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A. The Cellosaurus, a Cell-Line Knowledge Resource. J. Biomol. Tech. JBT 2018, 29, 25. [Google Scholar] [CrossRef]
- Rubinstein, L.V.; Crowley, J.; Ivy, P.; Leblanc, M.; Sargent, D.J. Randomized phase II designs. Clin. Cancer Res. 2009, 15, 1883–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilding, J.L.; Bodmer, W.F. Cancer Cell Lines for Drug Discovery and Development. Cancer Res. 2014, 74, 2377–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanni, P.; Landuzzi, L.; Manara, M.C.; Righi, A.; Nicoletti, G.; Cristalli, C.; Pasello, M.; Parra, A.; Carrabotta, M.; Ferracin, M.; et al. Bone sarcoma patient-derived xenografts are faithful and stable preclinical models for molecular and therapeutic investigations. Sci. Rep. 2019, 9, 12174. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.M. Patient-derived orthotopic xenografts: Better mimic of metastasis than subcutaneous xenografts. Nat. Rev. Cancer 2015, 15, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K.; Kiyuna, T.; Kawaguchi, K.; Higuchi, T.; Oshiro, H.; Zhang, Z.; Wangsiricharoen, S.; Razmjooei, S.; Li, Y.; Nelson, S.D.; et al. Regorafenib regressed a doxorubicin-resistant Ewing’s sarcoma in a patient-derived orthotopic xenograft (PDOX) nude mouse model. Cancer Chemother. Pharmacol. 2019, 83, 809–815. [Google Scholar] [CrossRef]
- Miyake, K.; Murakami, T.; Kiyuna, T.; Igarashi, K.; Kawaguchi, K.; Miyake, M.; Li, Y.; Nelson, S.D.; Dry, S.M.; Bouvet, M.; et al. The combination of temozolomide-irinotecan regresses a doxorubicin-resistant patient-derived orthotopic xenograft (PDOX) nude-mouse model of recurrent Ewing’s sarcoma with a FUS-ERG fusion and CDKN2A deletion: Direction for third-line patient therapy. Oncotarget 2017, 8, 103129–103136. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Singh, A.S.; Kiyuna, T.; Dry, S.M.; Li, Y.; James, A.W.; Igarashi, K.; Kawaguchi, K.; Delong, J.C.; Zhang, Y.; et al. Effective molecular targeting of CDK4/6 and IGF-1R in a rare FUS-ERG fusion CDKN2A-deletion doxorubicin-resistant Ewing’s sarcoma patient-derived orthotopic xenograft (PDOX) nude-mouse model. Oncotarget 2016, 7, 47556–47564. [Google Scholar] [CrossRef]
- Murakami, T.; Li, S.; Han, Q.; Tan, Y.; Kiyuna, T.; Igarashi, K.; Kawaguchi, K.; Hwang, H.K.; Miyake, K.; Singh, A.S.; et al. Recombinant methioninase effectively targets a Ewing’s sarcoma in a patient-derived orthotopic xenograft (PDOX) nude-mouse model. Oncotarget 2017, 8, 35630–35638. [Google Scholar] [CrossRef] [Green Version]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Bacteria as tumor-targeting vectors. Lancet Oncol. 2003, 4, 548–556. [Google Scholar] [CrossRef]
- Zhao, M.; Geller, J.; Ma, H.; Yang, M.; Penman, S.; Hoffman, R.M. Monotherapy with a tumor-targeting mutant of Salmonella typhimurium cures orthotopic metastatic mouse models of human prostate cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 10170–10174. [Google Scholar] [CrossRef] [Green Version]
- Miyake, K.; Kiyuna, T.; Li, S.; Han, Q.; Tan, Y.; Zhao, M.; Oshiro, H.; Kawaguchi, K.; Higuchi, T.; Zhang, Z.; et al. Combining Tumor-Selective Bacterial Therapy with Salmonella typhimurium A1-R and Cancer Metabolism Targeting with Oral Recombinant Methioninase Regressed an Ewing’s Sarcoma in a Patient-Derived Orthotopic Xenograft Model. Chemotherapy 2018, 63, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Stebbing, J.; Paz, K.; Schwartz, G.K.; Wexler, L.H.; Maki, R.G.; Pollock, R.E.; Morris, R.; Cohen, R.; Shankar, A.; Blackman, G.; et al. Patient-derived xenografts for individualized care in advanced sarcoma. Cancer 2014, 120, 2006–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kondo, T. Current Status and Perspectives of Patient-Derived Models for Ewing’s Sarcoma. Cancers 2020, 12, 2520. https://doi.org/10.3390/cancers12092520
AMA Style
Kondo T. Current Status and Perspectives of Patient-Derived Models for Ewing’s Sarcoma. Cancers. 2020; 12(9):2520. https://doi.org/10.3390/cancers12092520
Chicago/Turabian StyleKondo, Tadashi. 2020. "Current Status and Perspectives of Patient-Derived Models for Ewing’s Sarcoma" Cancers 12, no. 9: 2520. https://doi.org/10.3390/cancers12092520
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.