Molecular Profiling for Predictors of Radiosensitivity in Patients with Breast or Head-and-Neck Cancer

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
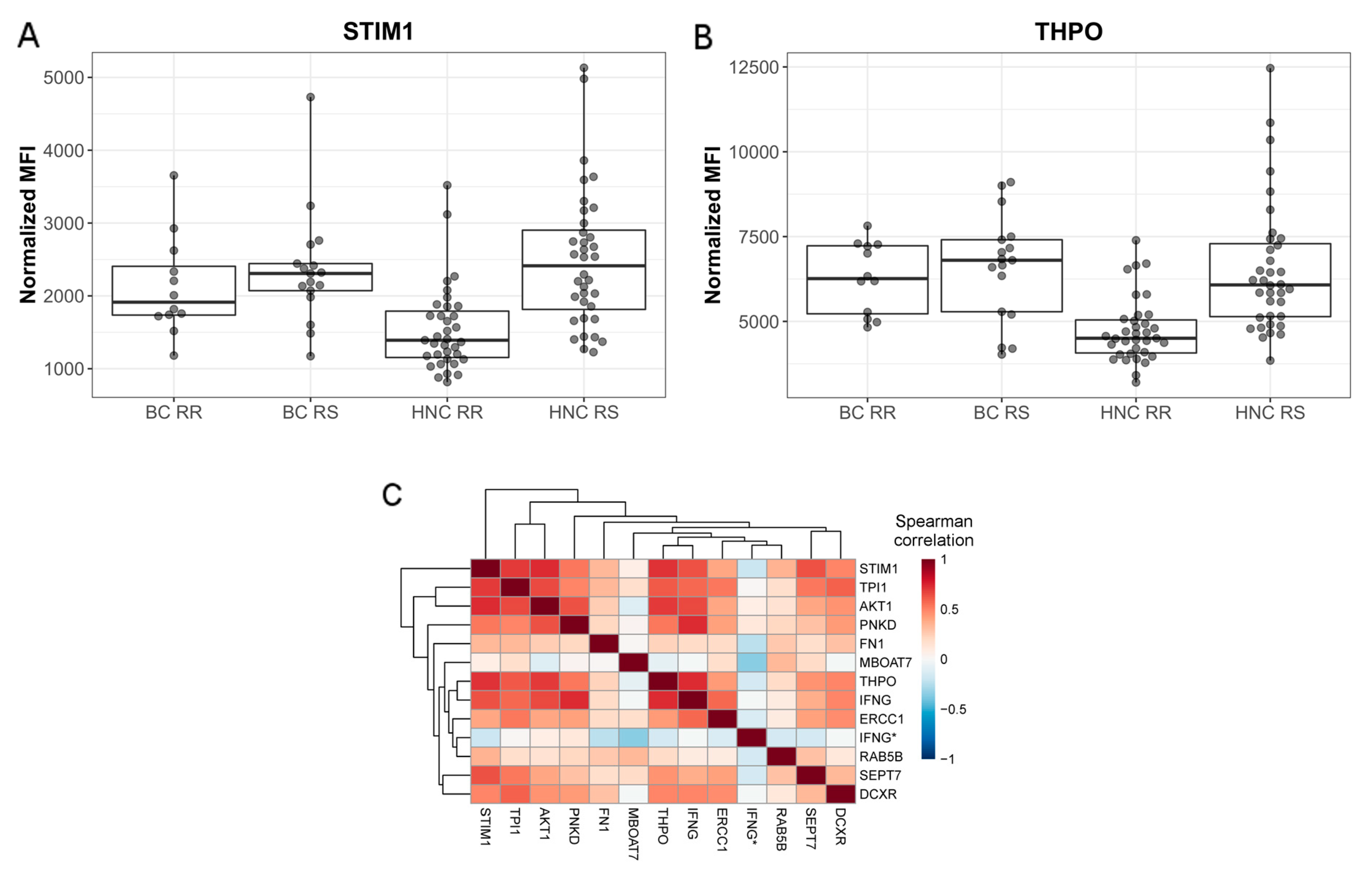
2.1. Profiling Patients with Normal or Adverse Healthy Tissue Response to Radiation
2.2. Patient Genotype and its Association with Radiosensitivity
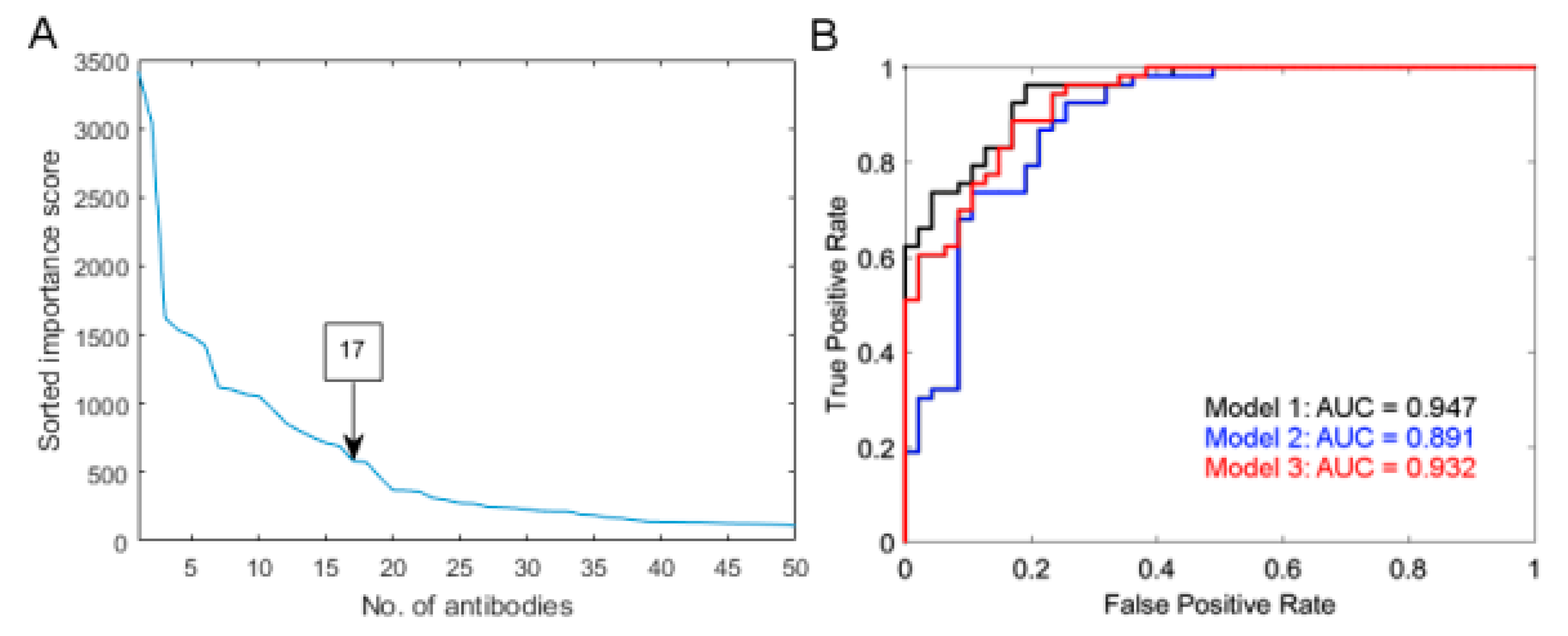
2.3. Multiple Protein Model Predictive of Radiosensitivity
3. Discussion
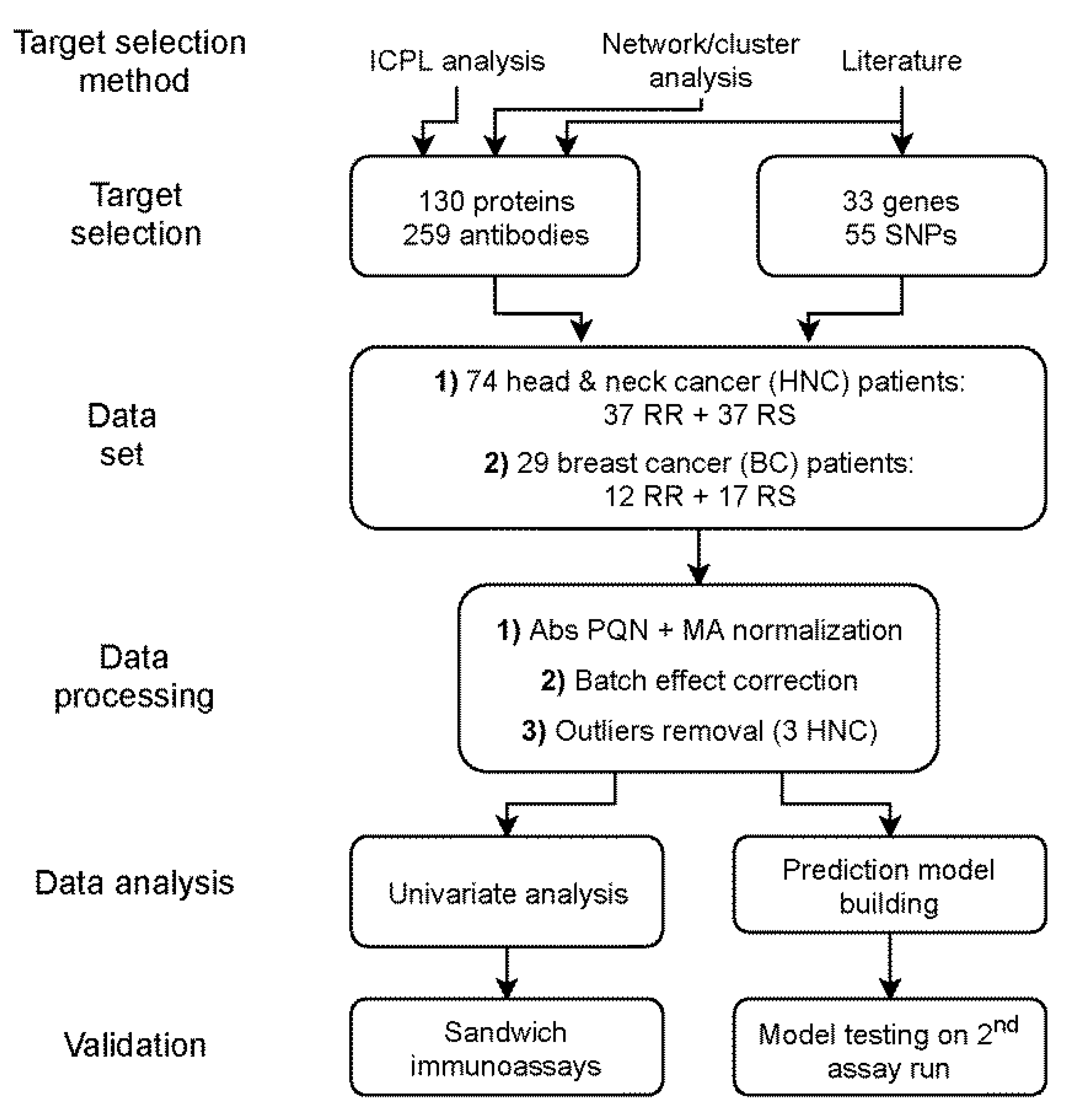
4. Materials and Methods
4.1. Study Population
4.1.1. Head-and-Neck Cancer Study Set
4.1.2. Breast Cancer Study Set
4.2. Antibody Bead Array Assay
4.2.1. Antibody Selection and Bead Coupling
4.2.2. Sample Randomization and Bead Array Processing
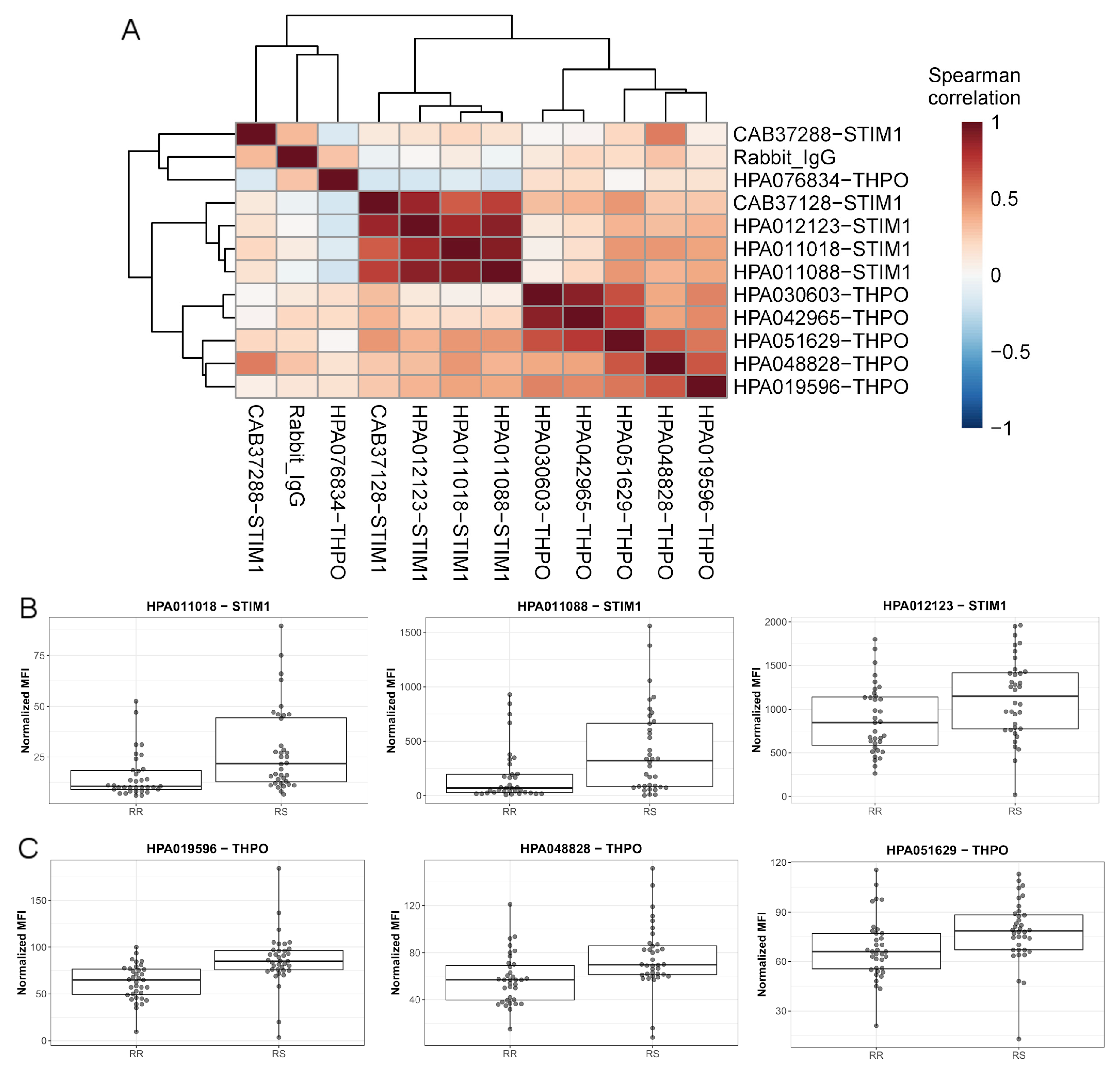
4.2.3. Antibody Validation
4.3. Single Nucleotide Polymorphism (SNP) Assay
4.4. Data Processing and Analysis
4.4.1. Data Pre-Processing
4.4.2. Univariate Comparisons
4.4.3. Prediction Model Building
4.5. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Fundings
Acknowledgments
Conflicts of Interest
References
- Ferlazzo, M.L.; Bourguignon, M.; Foray, N. Functional Assays for Individual Radiosensitivity: A Critical Review. Semin. Radiat. Oncol. 2017, 27, 310–315. [Google Scholar] [CrossRef]
- Fukunaga, H.; Yokoya, A.; Taki, Y.; Butterworth, K.T.; Prise, K.M. Precision Radiotherapy and Radiation Risk Assessment: How Do We Overcome Radiogenomic Diversity? Tohoku J. Exp. Med. 2019, 247, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Haghdoost, S.; Svoboda, P.; Naslund, I.; Harms-Ringdahl, M.; Tilikides, A.; Skog, S. Can 8-oxo-dG be used as a predictor for individual radiosensitivity? Int. J. Radiat. Oncol. Biol. Phys. 2001, 50, 405–410. [Google Scholar] [CrossRef]
- West, K.; Schneider, M.; Wright, C.; Beldham-Collins, R.; Coburn, N.; Tiver, K.; Gebski, V.; Stuart, K.E. Radiation-induced oesophagitis in breast cancer: Factors influencing onset and severity for patients receiving supraclavicular nodal irradiation. J. Med. Imaging Radiat. Oncol. 2019, 64, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, D.; Brehwens, K.; Halle, M.; Marczyk, M.; Sollazzo, A.; Polanska, J.; Munck-Wikland, E.; Wojcik, A.; Haghdoost, S. Influence of genetic background and stress response on risk of mandibular osteoradionecrosis after radiotherapy of head and neck cancer. Head Neck 2014, 38, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Skiold, S.; Azimzadeh, O.; Merl-Pham, J.; Naslund, I.; Wersall, P.; Lidbrink, E.; Tapio, S.; Harms-Ringdahl, M.; Haghdoost, S. Unique proteomic signature for radiation sensitive patients; a comparative study between normo-sensitive and radiation sensitive breast cancer patients. Mutat. Res. 2015, 776, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Bodgi, L.; Duclos, M.; Canet, A.; Ferlazzo, M.L.; Devic, C.; Granzotto, A.; Deneuve, S.; Vogin, G.; Foray, N. Fast and Binary Assay for Predicting Radiosensitivity Based on the Theory of ATM Nucleo-Shuttling: Development, Validation, and Performance. Int. J. Radiat. Oncol. Biol. Phys. 2018, 100, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Azria, D.; Riou, O.; Castan, F.; Nguyen, T.D.; Peignaux, K.; Lemanski, C.; Lagrange, J.L.; Kirova, Y.; Lartigau, E.; Belkacemi, Y.; et al. Radiation-induced CD8 T-lymphocyte Apoptosis as a Predictor of Breast Fibrosis After Radiotherapy: Results of the Prospective Multicenter French Trial. EBioMedicine 2015, 2, 1965–1973. [Google Scholar] [CrossRef] [Green Version]
- Popanda, O.; Marquardt, J.U.; Chang-Claude, J.; Schmezer, P. Genetic variation in normal tissue toxicity induced by ionizing radiation. Mutat. Res. 2008, 667, 58–69. [Google Scholar] [CrossRef]
- Vogin, G.; Bastogne, T.; Bodgi, L.; Gillet-Daubin, J.; Canet, A.; Pereira, S.; Foray, N. The Phosphorylated ATM Immunofluorescence Assay: A High-performance Radiosensitivity Assay to Predict Postradiation Therapy Overreactions. Int. J. Radiat. Oncol. Biol. Phys. 2018, 101, 690–693. [Google Scholar] [CrossRef] [Green Version]
- Talbot, C.J.; Veldwijk, M.R.; Azria, D.; Batini, C.; Bierbaum, M.; Brengues, M.; Chang-Claude, J.; Johnson, K.; Keller, A.; Smith, S.; et al. Multi-centre technical evaluation of the radiation-induced lymphocyte apoptosis assay as a predictive test for radiotherapy toxicity. Clin. Transl. Radiat. Oncol. 2019, 18, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Averbeck, D.; Candeias, S.; Chandna, S.; Foray, N.; Friedl, A.A.; Haghdoost, S.; Jeggo, P.A.; Lumniczky, K.; Paris, F.; Quintens, R.; et al. Establishing mechanisms affecting the individual response to ionizing radiation. Int. J. Radiat. Biol. 2020, 96, 297–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ruyck, K.; Van Eijkeren, M.; Claes, K.; Morthier, R.; De Paepe, A.; Vral, A.; De Ridder, L.; Thierens, H. Radiation-induced damage to normal tissues after radiotherapy in patients treated for gynecologic tumors: Association with single nucleotide polymorphisms in XRCC1, XRCC3, and OGG1 genes and in vitro chromosomal radiosensitivity in lymphocytes. Int. J. Radiat. Oncol. Biol. Phys. 2005, 62, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Kerns, S.L.; Dorling, L.; Fachal, L.; Bentzen, S.; Pharoah, P.D.; Barnes, D.R.; Gomez-Caamano, A.; Carballo, A.M.; Dearnaley, D.P.; Peleteiro, P.; et al. Meta-analysis of Genome Wide Association Studies Identifies Genetic Markers of Late Toxicity Following Radiotherapy for Prostate Cancer. EBioMedicine 2016, 10, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Russell, N.S.; Begg, A.C. Editorial radiotherapy and oncology 2002: Predictive assays for normal tissue damage. Radiother. Oncol. 2002, 64, 125–129. [Google Scholar] [CrossRef]
- Skiöld, S.; Becker, S.; Auer, G.; Hellman, U.; Näslund, I.; Harms-Ringdahl, M.; Haghdoost, S. Low doses of γ–radiation induce consistent protein expression changes in human leukocytes. Int. J. Low Radiat. 2011, 8, 374–387. [Google Scholar] [CrossRef]
- Skiold, S.; Naslund, I.; Brehwens, K.; Andersson, A.; Wersall, P.; Lidbrink, E.; Harms-Ringdahl, M.; Wojcik, A.; Haghdoost, S. Radiation-induced stress response in peripheral blood of breast cancer patients differs between patients with severe acute skin reactions and patients with no side effects to radiotherapy. Mutat. Res. 2013, 756, 152–157. [Google Scholar] [CrossRef]
- Loseva, O.; Shubbar, E.; Haghdoost, S.; Evers, B.; Helleday, T.; Harms-Ringdahl, M. Chronic Low Dose Rate Ionizing Radiation Exposure Induces Premature Senescence in Human Fibroblasts that Correlates with Up Regulation of Proteins Involved in Protection against Oxidative Stress. Proteomes 2014, 2, 341–362. [Google Scholar] [CrossRef] [Green Version]
- Yentrapalli, R.; Azimzadeh, O.; Barjaktarovic, Z.; Sarioglu, H.; Wojcik, A.; Harms-Ringdahl, M.; Atkinson, M.J.; Haghdoost, S.; Tapio, S. Quantitative proteomic analysis reveals induction of premature senescence in human umbilical vein endothelial cells exposed to chronic low-dose rate gamma radiation. Proteomics 2013, 13, 1096–1107. [Google Scholar] [CrossRef]
- Yentrapalli, R.; Azimzadeh, O.; Sriharshan, A.; Malinowsky, K.; Merl, J.; Wojcik, A.; Harms-Ringdahl, M.; Atkinson, M.J.; Becker, K.F.; Haghdoost, S.; et al. The PI3K/Akt/mTOR pathway is implicated in the premature senescence of primary human endothelial cells exposed to chronic radiation. PLoS ONE 2013, 8, e70024. [Google Scholar] [CrossRef] [Green Version]
- Bystrom, S.; Ayoglu, B.; Haggmark, A.; Mitsios, N.; Hong, M.G.; Drobin, K.; Forsstrom, B.; Fredolini, C.; Khademi, M.; Amor, S.; et al. Affinity proteomic profiling of plasma, cerebrospinal fluid, and brain tissue within multiple sclerosis. J. Proteome Res. 2014, 13, 4607–4619. [Google Scholar] [CrossRef] [PubMed]
- Haussler, R.S.; Bendes, A.; Iglesias, M.; Sanchez-Rivera, L.; Dodig-Crnkovic, T.; Bystrom, S.; Fredolini, C.; Birgersson, E.; Dale, M.; Edfors, F.; et al. Systematic Development of Sandwich Immunoassays for the Plasma Secretome. Proteomics 2019, 19, e1900008. [Google Scholar] [CrossRef] [Green Version]
- Alsner, J.; Andreassen, C.N.; Overgaard, J. Genetic markers for prediction of normal tissue toxicity after radiotherapy. Semin. Radiat. Oncol. 2008, 18, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, C.N.; Alsner, J.; Overgaard, M.; Sorensen, F.B.; Overgaard, J. Risk of radiation-induced subcutaneous fibrosis in relation to single nucleotide polymorphisms in TGFB1, SOD2, XRCC1, XRCC3, APEX and ATM—A study based on DNA from formalin fixed paraffin embedded tissue samples. Int. J. Radiat. Biol. 2006, 82, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Langsenlehner, T.; Renner, W.; Gerger, A.; Hofmann, G.; Thurner, E.M.; Kapp, K.S.; Langsenlehner, U. Impact of VEGF gene polymorphisms and haplotypes on radiation-induced late toxicity in prostate cancer patients. Strahlenther. Onkol. 2011, 187, 784–791. [Google Scholar] [CrossRef]
- Andreassen, C.N.; Alsner, J. Genetic variants and normal tissue toxicity after radiotherapy: A systematic review. Radiother. Oncol. 2009, 92, 299–309. [Google Scholar] [CrossRef]
- Zheng, Z.L.; Hwang, Y.H.; Kim, S.K.; Kim, S.; Son, M.J.; Ro, H.; Sung, S.A.; Lee, H.H.; Chung, W.K.; Joo, K.W.; et al. Genetic polymorphisms of hypoxia-inducible factor-1 alpha and cardiovascular disease in hemodialysis patients. Nephron Clin. Pract. 2009, 113, c104–c111. [Google Scholar] [CrossRef]
- Yin, M.; Liao, Z.; Huang, Y.J.; Liu, Z.; Yuan, X.; Gomez, D.; Wang, L.E.; Wei, Q. Polymorphisms of homologous recombination genes and clinical outcomes of non-small cell lung cancer patients treated with definitive radiotherapy. PLoS ONE 2011, 6, e20055. [Google Scholar] [CrossRef] [Green Version]
- Raabe, A.; Derda, K.; Reuther, S.; Szymczak, S.; Borgmann, K.; Hoeller, U.; Ziegler, A.; Petersen, C.; Dikomey, E. Association of single nucleotide polymorphisms in the genes ATM, GSTP1, SOD2, TGFB1, XPD and XRCC1 with risk of severe erythema after breast conserving radiotherapy. Radiat. Oncol. 2012, 7, 65. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Cho, J.W.; Lee, S.; Yun, A.; Kim, H.; Bae, D.; Yang, S.; Kim, C.Y.; Lee, M.; Kim, E.; et al. TRRUST v2: An expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018, 46, D380–D386. [Google Scholar] [CrossRef]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhsng, C.Z.; Wala, J.; Mermel, C.H.; et al. Pan-cancer patterns of somatic copy number alteration. Nat. Genet. 2013, 45, 1134–1140. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Leiserson, M.D.; Vandin, F.; Wu, H.T.; Dobson, J.R.; Eldridge, J.V.; Thomas, J.L.; Papoutsaki, A.; Kim, Y.; Niu, B.; McLellan, M.; et al. Pan-cancer network analysis identifies combinations of rare somatic mutations across pathways and protein complexes. Nat. Genet. 2015, 47, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Thariat, J.; Valable, S.; Laurent, C.; Haghdoost, S.; Peres, E.A.; Bernaudin, M.; Sichel, F.; Lesueur, P.; Cesaire, M.; Petit, E.; et al. Hadrontherapy Interactions in Molecular and Cellular Biology. Int. J. Mol. Sci. 2019, 21, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcellos-Hoff, M.H.; Park, C.; Wright, E.G. Radiation and the microenvironment—Tumorigenesis and therapy. Nat. Rev. Cancer 2005, 5, 867–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, Z.N.; Mi, J.; Zhang, Y.; Delong, M.; Jackson, I.L.; Fleckenstein, K.; Salahuddin, F.K.; Zhang, X.; Clary, B.; Anscher, M.S.; et al. Hypoxia inducible factor 1alpha signaling in fractionated radiation-induced lung injury: Role of oxidative stress and tissue hypoxia. Radiat. Res. 2010, 173, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Rofstad, E.K.; Gaustad, J.V.; Egeland, T.A.; Mathiesen, B.; Galappathi, K. Tumors exposed to acute cyclic hypoxic stress show enhanced angiogenesis, perfusion and metastatic dissemination. Int. J. Cancer 2010, 127, 1535–1546. [Google Scholar] [CrossRef]
- Liu, Y.; Kudo, K.; Abe, Y.; Aoki, M.; Hu, D.L.; Kijima, H.; Nakane, A. Hypoxia expression in radiation-induced late rectal injury. J. Radiat. Res. 2008, 49, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kudo, K.; Abe, Y.; Hu, D.L.; Kijima, H.; Nakane, A.; Ono, K. Inhibition of transforming growth factor-beta, hypoxia-inducible factor-1alpha and vascular endothelial growth factor reduced late rectal injury induced by irradiation. J. Radiat. Res. 2009, 50, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.A.; Benjamin, L.; Zeng, H.; Dvorak, A.M.; Dvorak, H.F. Vascular permeability, vascular hyperpermeability and angiogenesis. Angiogenesis 2008, 11, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Jaal, J.; Dorr, W. Radiation induced late damage to the barrier function of small blood vessels in mouse bladder. J. Urol. 2006, 176, 2696–2700. [Google Scholar] [CrossRef] [PubMed]
- Minchenko, A.; Salceda, S.; Bauer, T.; Caro, J. Hypoxia regulatory elements of the human vascular endothelial growth factor gene. Cell Mol. Biol. Res. 1994, 40, 35–39. [Google Scholar] [PubMed]
- Anscher, M.S.; Vujaskovic, Z. Mechanisms and potential targets for prevention and treatment of normal tissue injury after radiation therapy. Semin. Oncol. 2005, 32, S86–S91. [Google Scholar] [CrossRef] [PubMed]
- Lindegren, A.; Schultz, I.; Sinha, I.; Cheung, L.; Khan, A.A.; Tekle, M.; Wickman, M.; Halle, M. Autologous fat transplantation alters gene expression patterns related to inflammation and hypoxia in the irradiated human breast. Br. J. Surg. 2019, 106, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halle, M.; Gabrielsen, A.; Paulsson-Berne, G.; Gahm, C.; Agardh, H.E.; Farnebo, F.; Tornvall, P. Sustained inflammation due to nuclear factor-kappa B activation in irradiated human arteries. J. Am. Coll. Cardiol. 2010, 55, 1227–1236. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.B.; Xu, L.; Childress, P.J.; Maupin, K.A.; Mohamad, S.F.; Chitteti, B.R.; Himes, E.; Olivos, D.J., III; Cheng, Y.H.; Conway, S.J.; et al. Megakaryocyte and Osteoblast Interactions Modulate Bone Mass and Hematopoiesis. Stem Cells Dev. 2018, 27, 671–682. [Google Scholar] [CrossRef]
- Robinson, L.J.; Blair, H.C.; Barnett, J.B.; Soboloff, J. The roles of Orai and Stim in bone health and disease. Cell Calcium 2019, 81, 51–58. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Hirouchi, T.; Yokoyama, K.; Nishiyama, A.; Murakami, S.; Kashiwakura, I. The thrombopoietin mimetic romiplostim leads to the complete rescue of mice exposed to lethal ionizing radiation. Sci. Rep. 2018, 8, 10659. [Google Scholar] [CrossRef]
- De Laval, B.; Pawlikowska, P.; Petit-Cocault, L.; Bilhou-Nabera, C.; Aubin-Houzelstein, G.; Souyri, M.; Pouzoulet, F.; Gaudry, M.; Porteu, F. Thrombopoietin-increased DNA-PK-dependent DNA repair limits hematopoietic stem and progenitor cell mutagenesis in response to DNA damage. Cell Stem Cell 2013, 12, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Woodbine, L.; Gennery, A.R.; Jeggo, P.A. Reprint of “The clinical impact of deficiency in DNA non-homologous end-joining”. DNA Repair 2014, 17, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Hooper, R.; Samakai, E.; Kedra, J.; Soboloff, J. Multifaceted roles of STIM proteins. Pflugers Arch. 2013, 465, 1383–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancarella, S.; Wang, Y.; Deng, X.; Landesberg, G.; Scalia, R.; Panettieri, R.A.; Mallilankaraman, K.; Tang, X.D.; Madesh, M.; Gill, D.L. Hypoxia-induced acidosis uncouples the STIM-Orai calcium signaling complex. J. Biol. Chem. 2011, 286, 44788–44798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollak, C.E.; van Weely, S.; van Oers, M.H.; Aerts, J.M. Marked elevation of plasma chitotriosidase activity. A novel hallmark of Gaucher disease. J. Clin. Investig. 1994, 93, 1288–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, S.; Ishii, H.; Kazama, M. Heparin-like glycosaminoglycan is a receptor for antithrombin III-dependent but not for thrombin-dependent prostacyclin production in human endothelial cells. Thromb. Res. 1990, 59, 895–904. [Google Scholar] [CrossRef]
- Levy, J.H.; Sniecinski, R.M.; Welsby, I.J.; Levi, M. Antithrombin: Anti-inflammatory properties and clinical applications. Thromb. Haemost. 2016, 115, 712–728. [Google Scholar] [PubMed]
- Rao, F.V.; Houston, D.R.; Boot, R.G.; Aerts, J.M.; Sakuda, S.; van Aalten, D.M. Crystal structures of allosamidin derivatives in complex with human macrophage chitinase. J. Biol. Chem. 2003, 278, 20110–20116. [Google Scholar] [CrossRef] [Green Version]
- Wajner, A.; Michelin, K.; Burin, M.G.; Pires, R.F.; Pereira, M.L.; Giugliani, R.; Coelho, J.C. Biochemical characterization of chitotriosidase enzyme: Comparison between normal individuals and patients with Gaucher and with Niemann-Pick diseases. Clin. Biochem. 2004, 37, 893–897. [Google Scholar] [CrossRef]
- Bargagli, E.; Bennett, D.; Maggiorelli, C.; Di Sipio, P.; Margollicci, M.; Bianchi, N.; Rottoli, P. Human chitotriosidase: A sensitive biomarker of sarcoidosis. J. Clin. Immunol. 2013, 33, 264–270. [Google Scholar] [CrossRef]
- Mattsson, N.; Tabatabaei, S.; Johansson, P.; Hansson, O.; Andreasson, U.; Mansson, J.E.; Johansson, J.O.; Olsson, B.; Wallin, A.; Svensson, J.; et al. Cerebrospinal fluid microglial markers in Alzheimer’s disease: Elevated chitotriosidase activity but lack of diagnostic utility. Neuromolecul. Med. 2011, 13, 151–159. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, F.; Liang, M. SerpinC1/Antithrombin III in kidney-related diseases. Clin. Sci. 2017, 131, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Alper, S.L. Molecular physiology and genetics of Na+-independent SLC4 anion exchangers. J. Exp. Biol. 2009, 212, 1672–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivares-Gonzalez, L.; Martinez-Fernandez de la Camara, C.; Hervas, D.; Millan, J.M.; Rodrigo, R. HIF-1alpha stabilization reduces retinal degeneration in a mouse model of retinitis pigmentosa. FASEB J. 2018, 32, 2438–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penn, J.S.; Li, S.; Naash, M.I. Ambient hypoxia reverses retinal vascular attenuation in a transgenic mouse model of autosomal dominant retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4007–4013. [Google Scholar]
- Fosnot, J.; Fischer, J.P.; Smartt, J.M., Jr.; Low, D.W.; Kovach, S.J., III; Wu, L.C.; Serletti, J.M. Does previous chest wall irradiation increase vascular complications in free autologous breast reconstruction? Plast. Reconstr. Surg. 2011, 127, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Halle, M.; Eriksson, B.O.; Docherty Skogh, A.C.; Sommar, P.; Hammarstedt, L.; Gahm, C. Improved Head and Neck Free Flap Outcome-Effects of a Treatment Protocol Adjustment from Pre- to Postoperative Radiotherapy. Plast. Reconstr. Surg. Glob. Open 2017, 5, e1253. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, H.C.; Kagan, A.R. Osteoradionecrosis of the mandible: Scientific basis for clinical staging. Am. J. Clin. Oncol. 2002, 25, 168–171. [Google Scholar] [CrossRef]
- Drobin, K.; Nilsson, P.; Schwenk, J.M. Highly multiplexed antibody suspension bead arrays for plasma protein profiling. Methods Mol. Biol. 2013, 1023, 137–145. [Google Scholar]
- Kato, B.S.; Nicholson, G.; Neiman, M.; Rantalainen, M.; Holmes, C.C.; Barrett, A.; Uhlen, M.; Nilsson, P.; Spector, T.D.; Schwenk, J.M. Variance decomposition of protein profiles from antibody arrays using a longitudinal twin model. Proteome Sci. 2011, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Dezfouli, M.; Vickovic, S.; Iglesias, M.J.; Nilsson, P.; Schwenk, J.M.; Ahmadian, A. Magnetic bead assisted labeling of antibodies at nanogram scale. Proteomics 2014, 14, 14–18. [Google Scholar] [CrossRef]
- Jurinke, C.; van den Boom, D.; Cantor, C.R.; Koster, H. Automated genotyping using the DNA MassArray technology. Methods Mol. Biol. 2002, 187, 179–192. [Google Scholar]
- Hong, M.-G.; Lee, W.; Nilsson, P.; Pawitan, Y.; Schwenk, J.M. Multi-dimensional normalization in affinity-based proteomics. J. Proteome Res. 2016, 15, 3473–3480. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Team Remote Control. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- The MathWorks, Inc. MATLAB & Statistics Toolbox Release; The MathWorks, Inc.: Natick, MA, USA, 2016. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor | Model 1 | Model 2 | Model 3 |
|---|---|---|---|
| HPA011088 - STIM1 | 1.14 (0.99;1.31) | 1.13 (1.01;1.26) * | 1.20 (1.07;1.36) * |
| HPA011325 - PDGFB | 4.36 (1.42;13.39) * | 2.37 (1.07;5.27) * | 3.86 (1.50;9.91) * |
| HPA010134 - PNKD | 1.24 (0.61;2.51) | 1.35 (0.86;2.10) | 1.38 (0.77;2.48) |
| HPA030603 - THPO | 1.06 (0.97;1.16) | 1.05 (0.99;1.12) | 1.08 (1.01;1.16) * |
| HPA010115 - CHIT1 | 1.19 (1.03;1.38) * | 1.08 (0.98;1.19) | 1.17 (1.02;1.33) * |
| HPA000909 - RP2 | 2.36 (0.65;8.60) | 1.46 (0.64;3.34) | 2.19 (0.73;6.56) |
| HPA001816 - SERPINC1 | 0.93 (0.85;1.01) | 0.95 (0.89;1.00) | 0.92 (0.86;0.99) * |
| HPA063911 - SLC4A1 | 0.74 (0.45;1.24) | 0.55 (0.36;0.83) * | 0.40 (0.23;0.71) * |
| HPA004156 - AKT1 | 2.56 (0.76;8.63) | - | - |
| HPA035034 – GCA | 0.77 (0.56;1.056) | - | - |
| HPA027066 - FN1 | 1.06 (0.96;1.17) | - | - |
| HPA064755 – FGA | 0.96 (0.89;1.04) | - | - |
| HPA051370 – FGA | 1.12 (0.99;1.27) | - | - |
| HPA027735 - DBNL | 0.29 (0.08;1.09) | - | - |
| HPA041937 - BLVRB | 1.11 (0.84;1.46) | - | - |
| HPA004819 – PGR | 0.36 (0.16;0.81) * | - | - |
| HPA051420 - PPARA | 0.74 (0.50;1.11) | - | - |
| Rs69947 – AA/AC | - | - | 1 |
| Rs69947 – CC | - | - | 0.03 (0.00;0.22) * |
| Phase | Model | BC+HNC | BC only | HNC only | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sens | Spec | WErr | Sens | Spec | WErr | Sens | Spec | WErr | ||
| Training | 1 | 0.96 | 0.81 | 0.11 | 1.00 | 0.67 | 0.17 | 0.94 | 0.86 | 0.10 |
| 2 | 0.92 | 0.74 | 0.17 | 0.82 | 0.33 | 0.42 | 0.97 | 0.89 | 0.07 | |
| 3 | 0.89 | 0.83 | 0.14 | 0.82 | 0.50 | 0.34 | 0.92 | 0.94 | 0.07 | |
| Testing | 1 | 0.89 | 0.83 | 0.14 | 0.94 | 0.67 | 0.20 | 0.86 | 0.89 | 0.13 |
| 2 | 0.87 | 0.70 | 0.21 | 0.82 | 0.25 | 0.46 | 0.89 | 0.86 | 0.13 | |
| 3 | 0.85 | 0.81 | 0.17 | 0.76 | 0.50 | 0.37 | 0.89 | 0.91 | 0.10 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drobin, K.; Marczyk, M.; Halle, M.; Danielsson, D.; Papiez, A.; Sangsuwan, T.; Bendes, A.; Hong, M.-G.; Qundos, U.; Harms-Ringdahl, M.; et al. Molecular Profiling for Predictors of Radiosensitivity in Patients with Breast or Head-and-Neck Cancer. Cancers 2020, 12, 753. https://doi.org/10.3390/cancers12030753
Drobin K, Marczyk M, Halle M, Danielsson D, Papiez A, Sangsuwan T, Bendes A, Hong M-G, Qundos U, Harms-Ringdahl M, et al. Molecular Profiling for Predictors of Radiosensitivity in Patients with Breast or Head-and-Neck Cancer. Cancers. 2020; 12(3):753. https://doi.org/10.3390/cancers12030753
Chicago/Turabian StyleDrobin, Kimi, Michal Marczyk, Martin Halle, Daniel Danielsson, Anna Papiez, Traimate Sangsuwan, Annika Bendes, Mun-Gwan Hong, Ulrika Qundos, Mats Harms-Ringdahl, and et al. 2020. "Molecular Profiling for Predictors of Radiosensitivity in Patients with Breast or Head-and-Neck Cancer" Cancers 12, no. 3: 753. https://doi.org/10.3390/cancers12030753