Carbon-Ion Beam Irradiation Alone or in Combination with Zoledronic acid Effectively Kills Osteosarcoma Cells


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
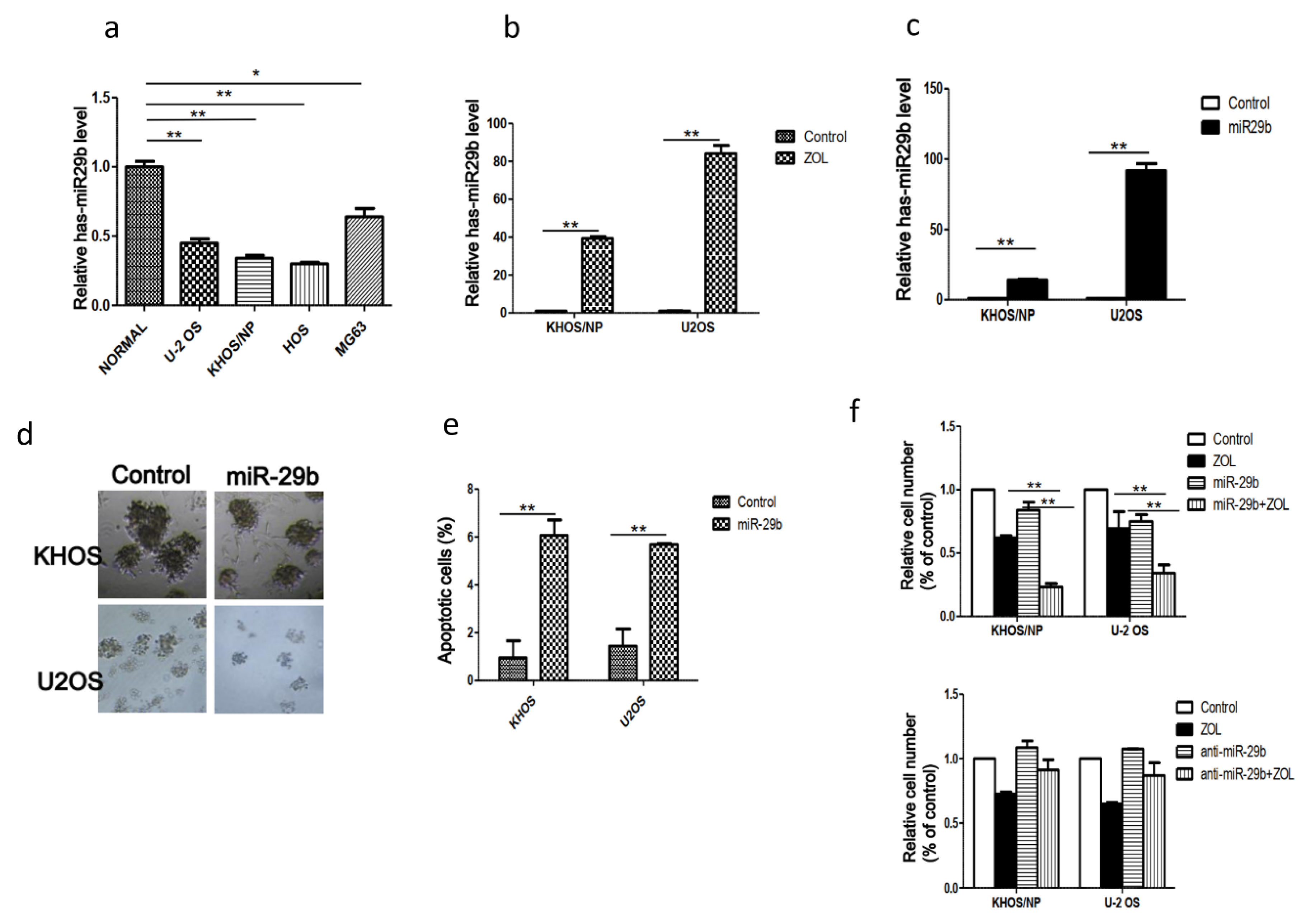
2.1. Expression of MiR-29b and Its Role in OSA Cell Proliferation and Apoptosis
2.2. Apoptosis Induction and Cell Cycle Aberration after Treatment with Carbon-Ion Beam Irradiation Alone or in Combination with ZOL in OSA Cells
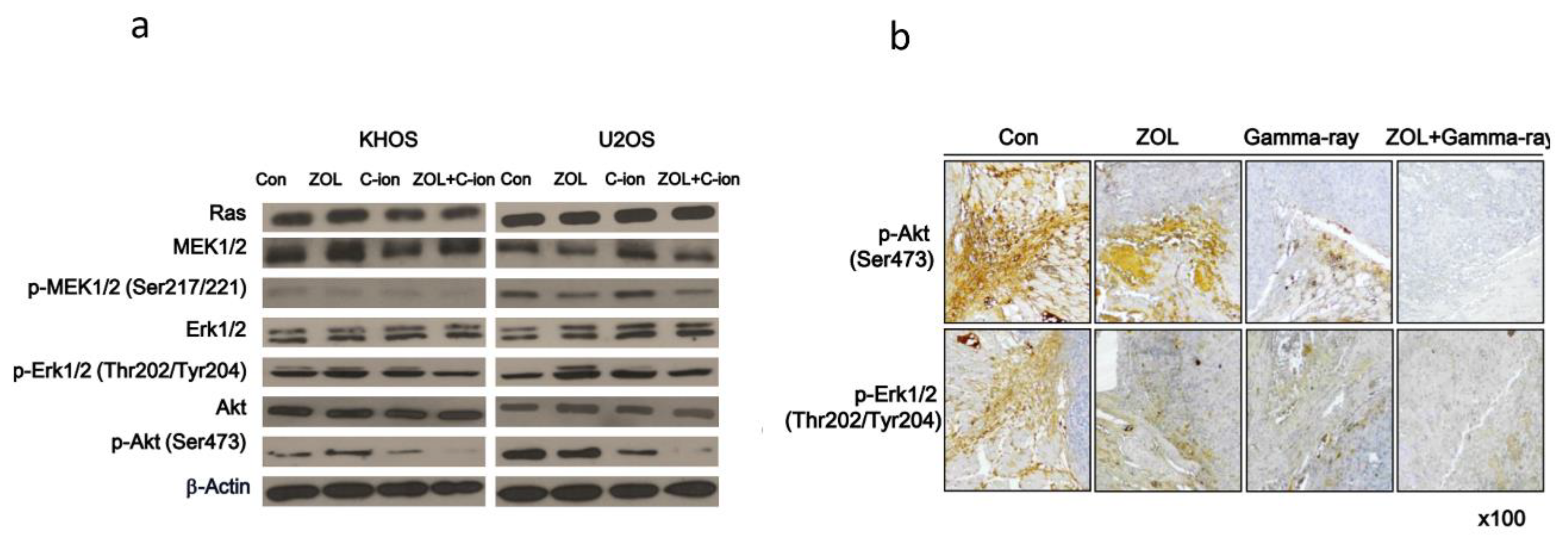
2.3. Involvement of PI3K–Akt and MAPK Signaling Pathways in OSA Cell Death after Carbon-Ion Beam Irradiation Alone or in Combination with ZOL
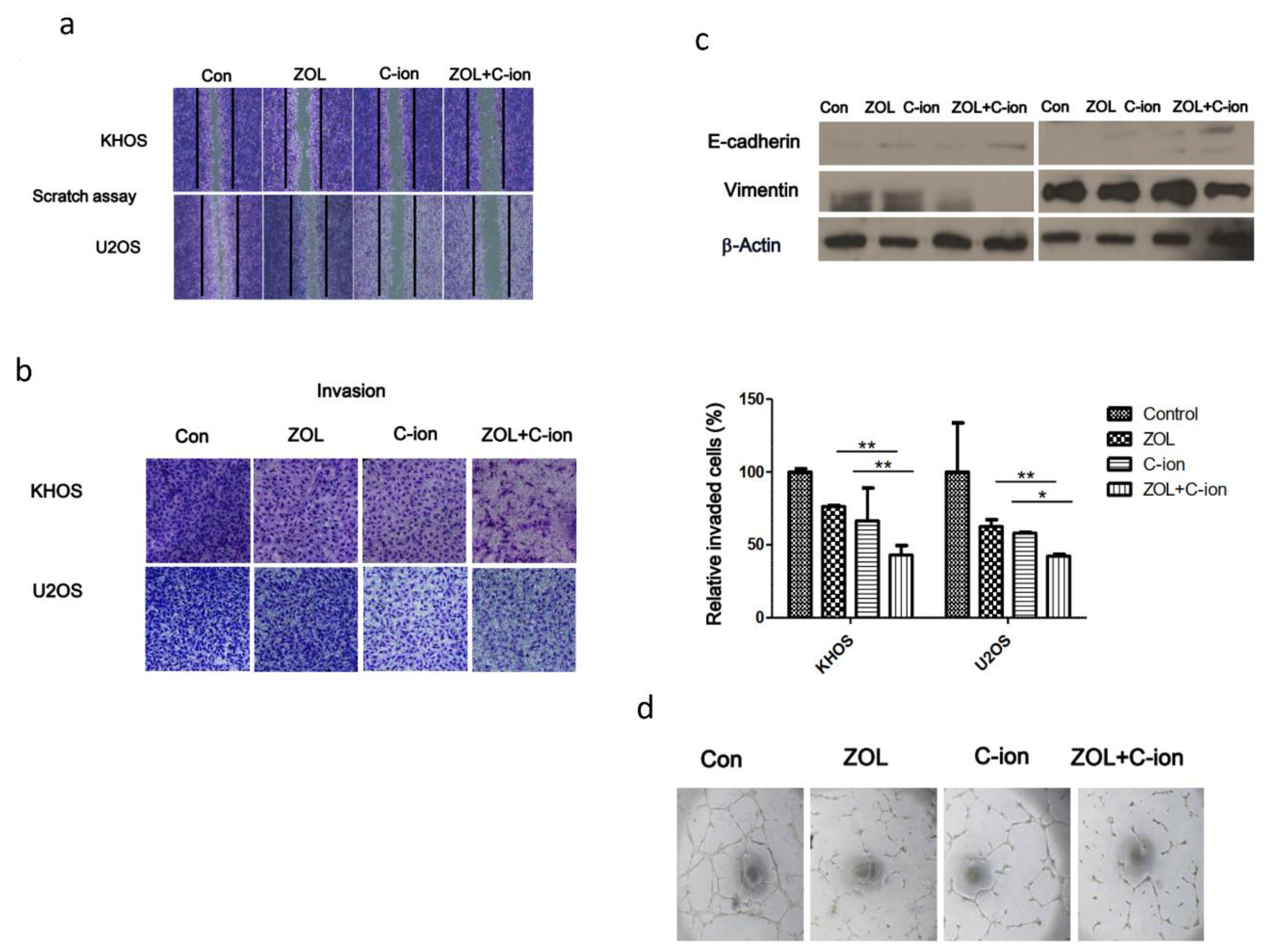
2.4. Inhibition of OSA Cell Motility, Invasion, and Angiogenesis after Treatment with Carbon-Ion Beam Irradiation Alone or in Combination with ZOL
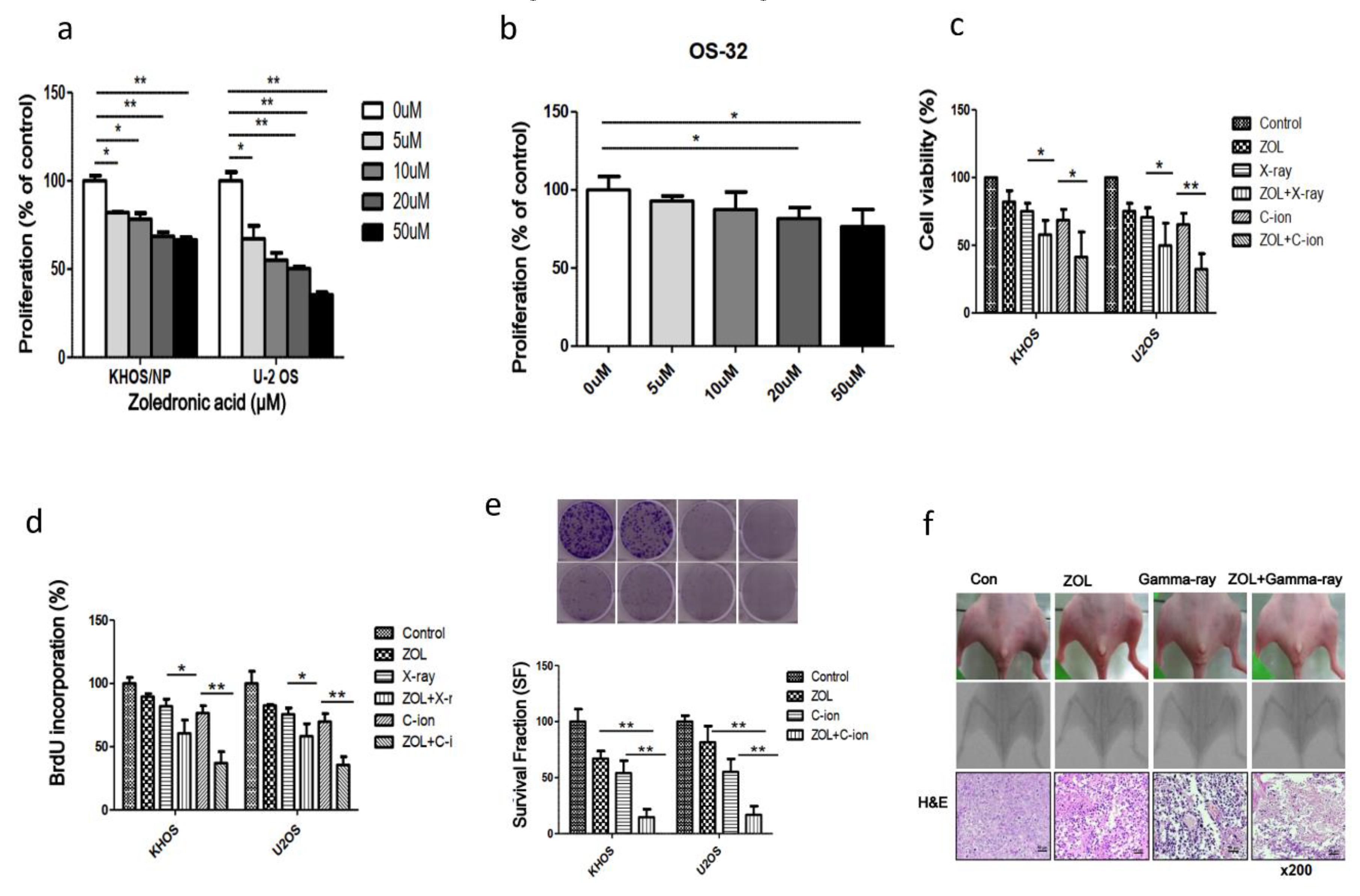
2.5. Effects of ZOL Alone or in Combination with Carbon-Ion Beam Irradiation on OSA Cell Proliferation
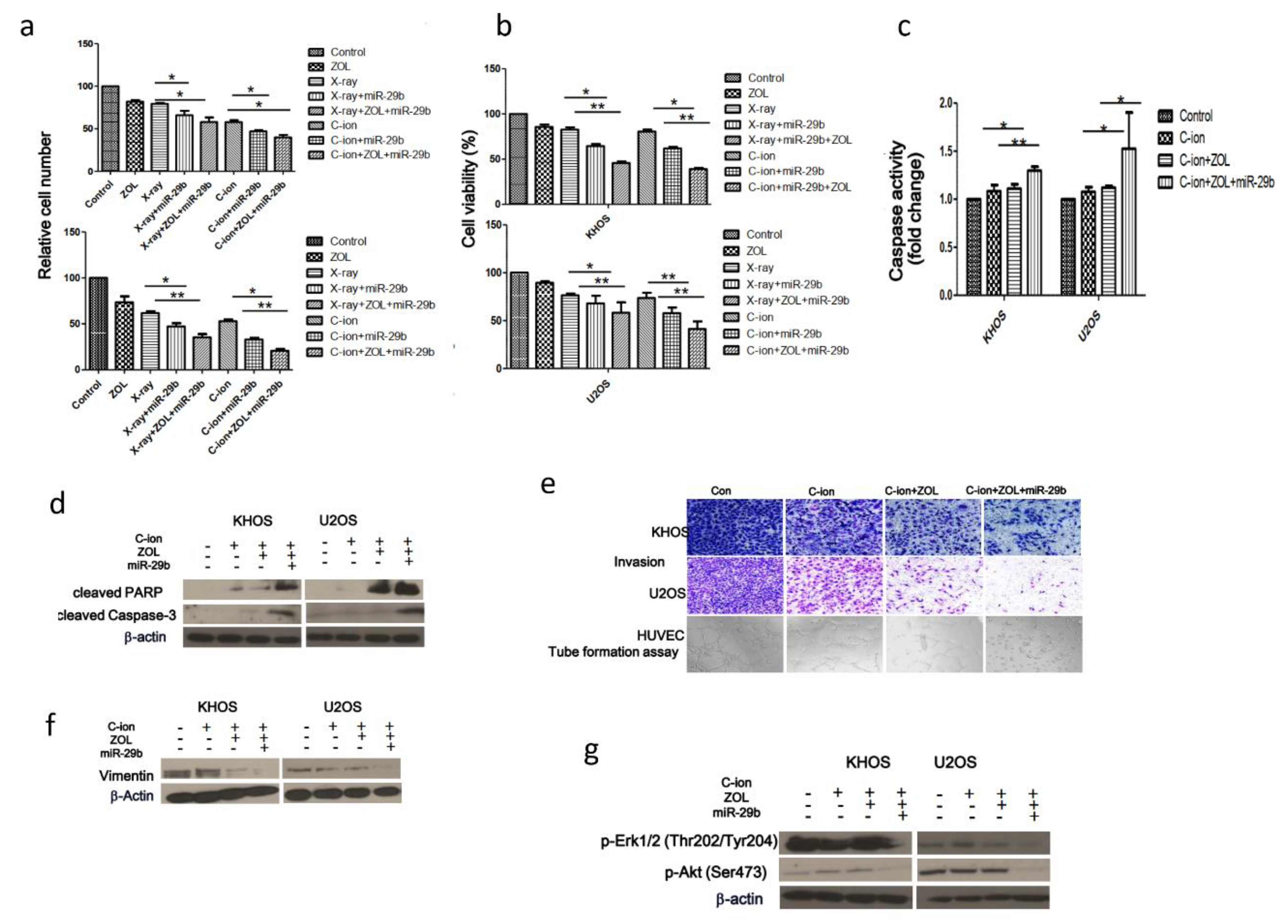
2.6. Suppression of OSA Cell Proliferation and Induction of Apoptosis After Treatment with Carbon-Ion Beam Irradiation Alone or in Combination with ZOL and the MiR-29b Mimic
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Tissue Samples
4.2. Reagents
4.3. Irradiation
4.4. Cell-Viability Assay
4.5. 5-Bromo-2′-Deoxyuridine (BrdU)-Labeling Assay
4.6. Colony Formation Assay
4.7. Orthotopic Animal Model and Histological Analysis
4.8. Caspase Activity Assay
4.9. Detection of Apoptotic Cells by Annexin V Staining
4.10. Western Blotting Analysis
4.11. Flow Cytometry
4.12. TUNEL Assays
4.13. Wound-Healing (Scratch) Assay
4.14. Transwell Chamber Assay
4.15. Matrigel-Based in Vitro Endothelial Tube-Formation Assay
4.16. miRNA and Transient Transfection
4.17. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Geller, D.S.; Gorlick, R. Osteosarcoma: A review of diagnosis, management, and treatment strategies. Clin. Adv. Hematol. Oncol. 2010, 8, 705–718. [Google Scholar] [PubMed]
- Lindsey, B.A.; Markel, J.E.; Kleinerman, E.S. Osteosarcoma overview. Rheumatol. Ther. 2017, 4, 25–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, J.; Ahluwalia, M.K.; Geller, D.; Gorlick, R. New targets and approaches in osteosarcoma. Pharmacol. Ther. 2013, 137, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Luetke, A.; Meyers, P.A.; Lewis, I.; Juergens, H. Osteosarcoma treatment—where do we stand? A state of the art review. Cancer Treat Rev. 2014, 40, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.; Bruland, O.; Cassoni, A.; Schomberg, P.; Bielack, S. The role of radiotherapy in oseosarcoma. Cancer Treat Res. 2009, 152, 147–164. [Google Scholar] [CrossRef]
- DeLaney, T.F.; Park, L.; Goldberg, S.I.; Hug, E.B.; Liebsch, N.J.; Munzenrider, J.E.; Suit, H.D. Radiotherapy for local control of osteosarcoma. Int. J. Radiat. Oncol. Biol. Phys. 2005, 61, 492–498. [Google Scholar] [CrossRef]
- Heng, M.; Gupta, A.; Chung, P.W.; Healey, J.H.; Vaynrub, M.; Rose, P.S.; Houdek, M.T.; Lin, P.P.; Bishop, A.J.; Hornicek, F.J.; et al. Japanese Musculoskeletal Oncology Group (JMOG); Soft Tissue Osteosarcoma International Collaborative (STOIC). The role of chemotherapy and radiotherapy in localized extraskeletal osteosarcoma. Eur. J. Cancer 2020, 125, 130–141. [Google Scholar] [CrossRef]
- Anderson, M.E. Update on survival in osteosarcoma. Orthop. Clin. North Am. 2016, 47, 283–292. [Google Scholar] [CrossRef]
- Kamada, T.; Tsujii, H.; Blakely, E.A.; Debus, J.; De Neve, W.; Durante, M.; Jäkel, O.; Mayer, R.; Orecchia, R.; Pötter, R.; et al. Carbon ion radiotherapy in Japan: An assessment of 20 years of clinical experience. Lancet. Oncol. 2015, 16, e93–e100. [Google Scholar] [CrossRef] [Green Version]
- Mohamad, O.; Imai, R.; Kamada, T.; Nitta, Y.; Araki, N. Working group for bone and soft tissue sarcoma. Carbon ion radiotherapy for inoperable pediatric osteosarcoma. Oncotarget 2018, 9, 22976–22985. [Google Scholar] [CrossRef] [Green Version]
- Demizu, Y.; Jin, D.; Sulaiman, N.S.; Nagano, F.; Terashima, K.; Tokumaru, S.; Akagi, T.; Fujii, O.; Daimon, T.; Sasaki, R.; et al. Particle therapy using protons or carbon ions for unresectable or incompletely resected bone and soft tissue sarcomas of the pelvis. Int. J. Radiat. Oncol. Biol. Phys. 2017, 98, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Imai, R.; Kamada, T.; Tsuji, H.; Tsujii, H.; Tsuburai, Y.; Tatezaki, S. Working group for bone and soft tissue sarcomas. Cervical spine osteosarcoma treated with carbon-ion radiotherapy. Lancet. Oncol. 2006, 7, 1034–1035. [Google Scholar] [CrossRef]
- Matsunobu, A.; Imai, R.; Kamada, T.; Imaizumi, T.; Tsuji, H.; Tsujii, H.; Shioyama, Y.; Honda, H.; Tatezaki, S. Working Group for Bone and Soft Tissue Sarcomas. Impact of carbon ion radiotherapy for unresectable osteosarcoma of the trunk. Cancer 2012, 118, 4555–4563. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, S.; Kamada, T.; Imai, R.; Tsuji, H.; Kameda, N.; Okada, T.; Tsujii, H.; Tatezaki, S. Working Group for the Bone and Soft Tissue Sarcomas. Carbon ion radiotherapy for localized primary sarcoma of the extremities: Results of a phase I/II trial. Radiother Oncol. 2012, 105, 226–231. [Google Scholar] [CrossRef]
- Cui, X.; Oonishi, K.; Tsujii, H.; Yasuda, T.; Matsumoto, Y.; Furusawa, Y.; Akashi, M.; Kamada, T.; Okayasu, R. Effects of carbon ion beam on putative colon cancer stem cells and its comparison with X-rays. Cancer Res. 2011, 71, 3676–3687. [Google Scholar] [CrossRef] [Green Version]
- Oonishi, K.; Cui, X.; Hirakawa, H.; Fujimori, A.; Yamada, S.; Yokosuka, O.; Kamada, T. Different effects of carbon ion beams and X-rays on clonogenic survival and DNA repair in human pancreatic cancer stem-like cells. Radiother Oncol. 2012, 105, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Sai, S.; Wakai, T.; Vares, G.; Yamada, S.; Kamijo, T.; Kamada, T.; Shirai, T. Combination of carbon ion beam and gemcitabine causes unreparable DNA damage and death of radioresistant pancreatic cancer stem-like cells in vitro and in vivo. Oncotarget 2015, 6, 5517–5535. [Google Scholar] [CrossRef] [Green Version]
- Sai, S.; Vares, G.; Kim, E.H.; Karasawa, K.; Wang, B.; Nenoi, M.; Horimoto, Y.; Hayashi, M. Carbon ion beam combined with cisplatin effectively disrupts triple negative breast cancer stem-like cells in vitro. Mol. Cancer 2015, 14, 166. [Google Scholar] [CrossRef] [Green Version]
- Sai, S.; Suzuki, M.; Kim, E.H.; Hayahsi, M.; Yamamoto, N.; Miyamoto, T. Effects of carbon ion beam alone or in combination with cisplatin on malignant mesothelioma cells in vitro. Oncotarget 2018, 9, 14849–14861. [Google Scholar] [CrossRef] [Green Version]
- Polascik, T.J. Bisphosphonates in oncology: Evidence for the prevention of skeletal events in patients with bone metastases. Drug Des. Devel. Ther. 2009, 3, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Peng, W.; Yang, C.; Zheng, J. Teriparatide versus bisphosphonates for treatment of postmenopausal osteoporosis: A meta-analysis. Int. J. Surg. 2019, 66, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vadhan-Raj, S.; von Moos, R.; Fallowfield, L.J.; Patrick, D.L.; Goldwasser, F.; Cleeland, C.S.; Henry, D.H.; Novello, S.; Hungria, V.; Qian, Y.; et al. Clinical benefit in patients with metastatic bone disease: Results of a phase 3 study of denosumab versus zoledronic acid. Ann. Oncol. 2012, 23, 3045–3051. [Google Scholar] [CrossRef] [PubMed]
- Groenen, K.H.; Pouw, M.H.; Hannink, G.; Hosman, A.J.; van der Linden, Y.M.; Verdonschot, N.; Tanck, E. The effect of radiotherapy, and radiotherapy combined with bisphosphonates or RANK ligand inhibitors on bone quality in bone metastases. A systematic review. Radiother Oncol. 2016, 119, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, C.; Wang, Y.; Li, H.; Huang, Y.; Jiang, O.; You, Y.; Luo, F. Zoledronic acid augments the radiosensitivity of cancer cells through perturbing S- and M-phase cyclins and p21CIP1 expression. Oncol. Lett. 2017, 14, 4237–4242. [Google Scholar] [CrossRef]
- Rouhrazi, H.; Turgan, N.; Oktem, G. Zoledronic acid overcomes chemoresistance by sensitizing cancer stem cells to apoptosis. Biotech. Histochem. 2018, 93, 77–88. [Google Scholar] [CrossRef]
- Koto, K.; Murata, H.; Kimura, S.; Sawai, Y.; Horie, N.; Matsui, T.; Ryu, K.; Ashihara, E.; Maekawa, T.; Kubo, T.; et al. Zoledronic acid significantly enhances radiation induced apoptosis against human fibrosarcoma cells by inhibiting radioadaptive signaling. Int. J. Oncol. 2013, 42, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, T.D.; Pillai, S.P.; Hildreth, B.E., 3rd; Lanigan, L.G.; Martin, C.K.; Werbeck, J.L.; Rosol, T.J. Effect of zoledronic acid and amputation on bone invasion and lung metastasis of canine osteosarcoma in nude mice. Clin. Exp. Metastasis 2011, 28, 377–389. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Wang, Q.; Li, H.; Ma, Y.; Deng, Y.; Ye, Z.; Bai, F. Zoledronic acid exhibits radio-sensitizing activity in human pancreatic cancer cells via inactivation of STAT3/NF-κB signaling. Onco. Targets Ther. 2019, 12, 4323–4330. [Google Scholar] [CrossRef] [Green Version]
- Bühler, H.; Hoberg, C.; Fakhrian, K.; Adamietz, I.A. Zoledronic acid inhibits the motility of cancer stem-like cells from the human breast cancer cell line MDA-MB 231. In Vivo 2016, 30, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Kijima, T.; Koga, F.; Fujii, Y.; Yoshida, S.; Tatokoro, M.; Kihara, K. Zoledronic acid sensitizes renal cell carcinoma cells to radiation by downregulating STAT1. PLoS ONE 2013, 8, e64615. [Google Scholar] [CrossRef] [Green Version]
- Denham, J.W.; Joseph, D.; Lamb, D.S.; Spry, N.A.; Duchesne, G.; Matthews, J.; Atkinson, C.; Tai, K.H.; Christie, D.; Kenny, L.; et al. Short-term androgen suppression and radiotherapy versus intermediate-term androgen suppression and radiotherapy, with or without zoledronic acid, in men with locally advanced prostate cancer (TROG 03.04 RADAR): 10-year results from a randomised, phase 3, factorial trial. Lancet Oncol. 2019, 20, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, C.L.; Moriarty, J.P.; Dusetzina, S.; Himelstein, A.L.; Foster, J.C.; Grubbs, S.S.; Novotny, P.J.; Borah, B.J. Cost-effectiveness analysis of monthly zoledronic acid, zoledronic acid every 3 months, and monthly denosumab in women with breast cancer and skeletal metastases: CALGB 70604 (Alliance). J. Clin. Oncol. 2017, 35, 3949–3955. [Google Scholar] [CrossRef]
- Pichon, B.; Campion, L.; Delpon, G.; Thillays, F.; Carrie, C.; Cellier, P.; Pommier, P.; Laude, C.; Mervoyer, A.; Hamidou, H.; et al. High-dose hypofractionated radiation therapy for noncompressive vertebral metastases in combination with zoledronate: A phase 1 study. Int. J. Radiat. Oncol. Biol. Phys. 2016, 96, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Kim, M.S.; Lee, K.H.; Koh, J.S.; Jung, W.G.; Kong, C.B. Zoledronic acid is an effective radiosensitizer in the treatment of osteosarcoma. Oncotarget 2016, 7, 70869–70880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, D.; Qiu, X.; Zhuang, M.; Zhu, C.; Zou, H.; Liu, Z. MicroRNAs with prognostic significance in osteosarcoma: A systemic review and meta-analysis. Oncotarget 2017, 8, 81062–81074. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Kim, E.H.; Lee, Y.J.; Sai, S.; Lim, S.H.; Park, J.W.; Chung, H.K.; Kim, J.; Vares, G.; Takahashi, A.; et al. Synergistic autophagy effect of miR-212-3p in zoledronic acid-treated in vitro and orthotopic in vivo models and in patient-derived osteosarcoma cells. Cancers 2019, 11, 1812. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. microRNAs in cancer management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef]
- Li, G.; Zhao, J.; Peng, X.; Liang, J.; Deng, X.; Chen, Y. The mechanism involved in the loss of PTEN expression in NSCLC tumor cells. Biochem. Biophys. Res. Commun. 2012, 17, 547–552. [Google Scholar] [CrossRef]
- Zhang, T.; Xue, X.; Peng, H. Therapeutic delivery of miR-29b enhances radiosensitivity in cervical cancer. Mol. Ther. 2019, 27, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- Dillon, R.L.; White, D.E.; Muller, W.J. The phosphatidyl inositol 3-kinase signaling network: Implications for human breast cancer. Oncogene 2007, 26, 1338–1345. [Google Scholar] [CrossRef]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polascik, T.J.; Mouraviev, V. Zoledronic acid in the management of metastatic bone disease. Ther. Clin. Risk Manag. 2008, 4, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orita, Y.; Sugitani, I.; Takao, S.; Toda, K.; Manabe, J.; Miyata, S. Prospective evaluation of zoledronic acid in the treatment of bone metastases from differentiated thyroid carcinoma. Ann. Surg. Oncol. 2015, 22, 4008–4013. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Y.; Zhou, Y.; Wang, J.; Tu, L.; Sun, Z.; Wang, X.; Luo, F. Zoledronic acid inhibits the growth of cancer stem cell derived from cervical cancer cell by attenuating their stemness phenotype and inducing apoptosis and cell cycle arrest through the Erk1/2 and Akt pathways. J. Exp. Clin. Cancer Res. 2019, 38, 93. [Google Scholar] [CrossRef] [Green Version]
- Fragni, M.; Bonini, S.A.; Bettinsoli, P.; Bodei, S.; Generali, D.; Bottini, A.; Spano, P.F.; Memo, M.; Sigala, S. The miR-21/PTEN/Akt signaling pathway is involved in the anti-tumoral effects of zoledronic acid in human breast cancer cell lines. Naunyn. Schmiedebergs Arch. Pharmacol. 2016, 389, 529–538. [Google Scholar] [CrossRef]
- Jia, L.F.; Huang, Y.P.; Zheng, Y.F.; Lyu, M.Y.; Wei, S.B.; Meng, Z.; Gan, Y.H. miR-29b suppresses proliferation, migration, and invasion of tongue squamous cell carcinoma through PTEN-AKT signaling pathway by targeting Sp1. Oral Oncol. 2014, 50, 1062–1071. [Google Scholar] [CrossRef]
- Feng, C.; Liu, X.; Li, X.; Guo, F.; Huang, C.; Qin, Q.; Wang, Y. Zoledronic acid increases the antitumor effect of gefitinib treatment for non-small cell lung cancer with EGFR mutations. Oncol. Rep. 2016, 35, 3460–3470. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.; Shim, H.G.; Hwang, T.; Kim, H.; Kang, S.H.; Dho, Y.S.; Park, S.H.; Kim, S.J.; Park, C.K. Restoration of miR-29b exerts anti-cancer effects on glioblastoma. Cancer Cell Int. 2017, 17, 104. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.H.; Kim, M.-S.; Takahashi, A.; Suzuki, M.; Vares, G.; Uzawa, A.; Fujimori, A.; Ohno, T.; Sai, S. Carbon-Ion Beam Irradiation Alone or in Combination with Zoledronic acid Effectively Kills Osteosarcoma Cells. Cancers 2020, 12, 698. https://doi.org/10.3390/cancers12030698
Kim EH, Kim M-S, Takahashi A, Suzuki M, Vares G, Uzawa A, Fujimori A, Ohno T, Sai S. Carbon-Ion Beam Irradiation Alone or in Combination with Zoledronic acid Effectively Kills Osteosarcoma Cells. Cancers. 2020; 12(3):698. https://doi.org/10.3390/cancers12030698
Chicago/Turabian StyleKim, Eun Ho, Mi-Sook Kim, Akihisa Takahashi, Masao Suzuki, Guillaume Vares, Akiko Uzawa, Akira Fujimori, Tatsuya Ohno, and Sei Sai. 2020. "Carbon-Ion Beam Irradiation Alone or in Combination with Zoledronic acid Effectively Kills Osteosarcoma Cells" Cancers 12, no. 3: 698. https://doi.org/10.3390/cancers12030698