The Molecular Hallmarks of the Serrated Pathway in Colorectal Cancer

 , , ,
, , ,
Abstract
:
1. Introduction
The Conventional Model of Colorectal Carcinogenesis and the “Serrated” Pathway
- CIMP-H, MLH1 methylated, MSI, BRAF mutated lesions, known as sporadic MSI;
- CIMP-H, MLH1 partially methylated, MSS, BRAF mutated lesions;
- CIMP-L, MGMT methylated, MSS, KRAS mutated lesions.
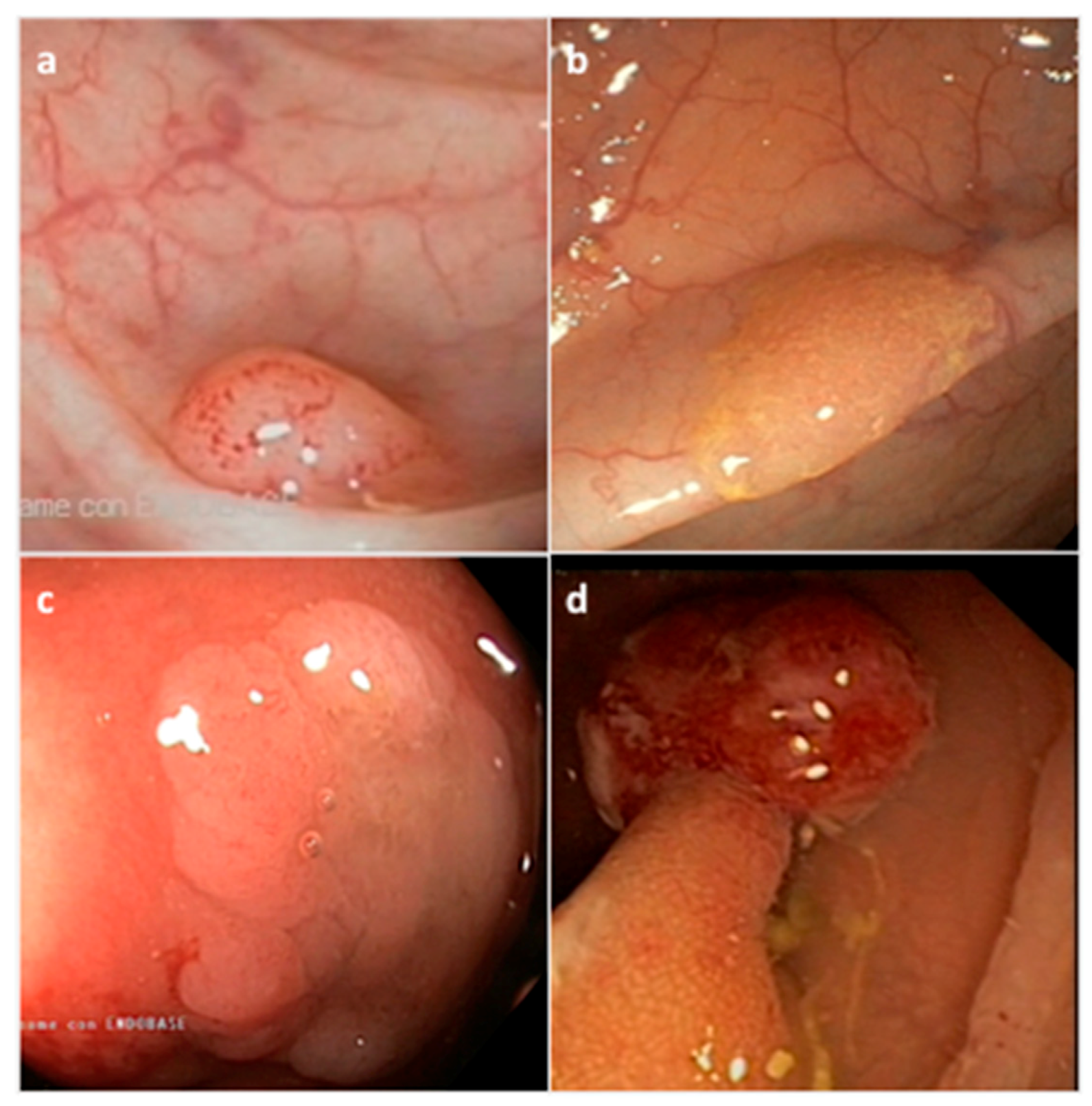
2. Histopathological and Endoscopic Features of Serrated Colorectal Lesions
3. CpG island Methylator Phenotype, an Epigenetic Signature of Serrated Colorectal Lesions
4. Molecular Features of the Serrated Colorectal Precursor Lesions
4.1. Hyperplastic Polyps
4.2. Sessile Serrated Adenoma/Polyps
4.3. Sessile Serrated Adenoma/Polyps with Dysplasia
4.4. Traditional Serrated Adenoma/Polyps with and without Dysplasia
5. Serrated Adenoma to Carcinoma Sequence: Initiation and Progression
A Distinct Variant of Colorectal Cancer: Serrated Adenocarcinoma
6. Superficially Serrated Adenoma
7. MicroRNAs and Long Non-Coding RNAs in Serrated Colorectal Pathway
8. Dysbiosis of the Gut Microbiota: A New Biomarker?
9. Clinical Relevance of Molecular Alterations in the Serrated Colorectal Pathway
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primer 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Nguyen, H.T.; Duong, H.-Q.; Duong, H.-Q. The molecular characteristics of colorectal cancer: Implications for diagnosis and therapy (Review). Oncol. Lett. 2018, 16, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogaert, J.; Prenen, H. Molecular genetics of colorectal cancer. Ann. Gastroenterol. 2014, 27, 9–14. [Google Scholar] [PubMed]
- Wong, K.; Xie, G. Updates on the Molecular Genetics of Colorectal Cancer. Colorectal Cancer Open Access 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, E.J.; Rösch, T.; Bretthauer, M. Colorectal cancer screening--optimizing current strategies and new directions. Nat. Rev. Clin. Oncol. 2013, 10, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Bardhan, K.; Liu, K. Epigenetics and colorectal cancer pathogenesis. Cancers 2013, 5, 676–713. [Google Scholar] [CrossRef] [PubMed]
- Grady, W.M.; Carethers, J.M. Genomic and epigenetic instability in colorectal cancer pathogenesis. Gastroenterology 2008, 135, 1079–1099. [Google Scholar] [CrossRef]
- Pancione, M.; Remo, A.; Colantuoni, V. Genetic and Epigenetic Events Generate Multiple Pathways in Colorectal Cancer Progression. Available online: https://www.hindawi.com/journals/pri/2012/509348/ (accessed on 27 February 2019).
- Vogelstein, B.; Fearon, E.R.; Hamilton, S.R.; Kern, S.E.; Preisinger, A.C.; Leppert, M.; Nakamura, Y.; White, R.; Smits, A.M.; Bos, J.L. Genetic alterations during colorectal-tumor development. N. Engl. J. Med. 1988, 319, 525–532. [Google Scholar] [CrossRef]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Cho, K.R.; Vogelstein, B. Genetic alterations in the adenoma--carcinoma sequence. Cancer 1992, 70, 1727–1731. [Google Scholar] [CrossRef]
- Armaghany, T.; Wilson, J.D.; Chu, Q.; Mills, G. Genetic Alterations in Colorectal Cancer. GCR 2012, 5, 19–27. [Google Scholar]
- Geigl, J.B.; Obenauf, A.C.; Schwarzbraun, T.; Speicher, M.R. Defining “chromosomal instability”. Trends Genet. 2008, 24, 64–69. [Google Scholar] [CrossRef]
- Boland, C.R.; Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 2010, 138, 2073–2087. [Google Scholar] [CrossRef]
- Gupta, R.; Sinha, S.; Paul, R.N. The impact of microsatellite stability status in colorectal cancer. Curr. Probl. Cancer 2018, 42, 548–559. [Google Scholar] [CrossRef]
- Pino, M.S.; Chung, D.C. The Chromosomal Instability Pathway in Colon Cancer. Gastroenterology 2010, 138, 2059–2072. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.J.; Yang, S.; Mack, C.; Xu, H.; Huang, C.S.; Mulcahy, E.; Amorosino, M.; Farraye, F.A. Comparison of microsatellite instability, CpG island methylation phenotype, BRAF and KRAS status in serrated polyps and traditional adenomas indicates separate pathways to distinct colorectal carcinoma end points. Am. J. Surg. Pathol. 2006, 30, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Gryfe, R.; Kim, H.; Hsieh, E.T.; Aronson, M.D.; Holowaty, E.J.; Bull, S.B.; Redston, M.; Gallinger, S. Tumor microsatellite instability and clinical outcome in young patients with colorectal cancer. N. Engl. J. Med. 2000, 342, 69–77. [Google Scholar] [CrossRef]
- Murcia, O.; Juárez, M.; Hernández-Illán, E.; Egoavil, C.; Giner-Calabuig, M.; Rodríguez-Soler, M.; Jover, R. Serrated colorectal cancer: Molecular classification, prognosis, and response to chemotherapy. World J. Gastroenterol. 2016, 22, 3516–3530. [Google Scholar] [CrossRef]
- Hegde, M.; Ferber, M.; Mao, R.; Samowitz, W.; Ganguly, A.; Working Group of the American College of Medical Genetics and Genomics (ACMG) Laboratory Quality Assurance Committee. ACMG technical standards and guidelines for genetic testing for inherited colorectal cancer (Lynch syndrome, familial adenomatous polyposis, and MYH-associated polyposis). Genet. Med. 2014, 16, 101–116. [Google Scholar] [CrossRef] [Green Version]
- Jass, J.R. Classification of colorectal cancer based on correlation of clinical, morphological and molecular features. Histopathology 2007, 50, 113–130. [Google Scholar] [CrossRef]
- Ogino, S.; Nosho, K.; Kirkner, G.J.; Kawasaki, T.; Meyerhardt, J.A.; Loda, M.; Giovannucci, E.L.; Fuchs, C.S. CpG island methylator phenotype, microsatellite instability, BRAF mutation and clinical outcome in colon cancer. Gut 2009, 58, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Kuroda, H.; Imai, Y.; Hiraishi, H. Molecular pathogenesis of sporadic colorectal cancers. Chin. J. Cancer 2016, 35. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.T.; Snyder, C.L.; Shaw, T.G.; Heinen, C.D.; Hitchins, M.P. Milestones of Lynch syndrome: 1895–2015. Nat. Rev. Cancer 2015, 15, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Tutlewska, K.; Lubinski, J.; Kurzawski, G. Germline deletions in the EPCAM gene as a cause of Lynch syndrome—Literature review. Hered. Cancer Clin. Pract. 2013, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Bessa, X.; Ballesté, B.; Andreu, M.; Castells, A.; Bellosillo, B.; Balaguer, F.; Castellví-Bel, S.; Paya, A.; Jover, R.; Alenda, C.; et al. A prospective, multicenter, population-based study of BRAF mutational analysis for Lynch syndrome screening. Clin. Gastroenterol. Hepatol. 2008, 6, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.T.; Ruszkiewicz, A. Serrated neoplasia of the colorectum. World J. Gastroenterol. 2007, 13, 3792–3798. [Google Scholar] [CrossRef] [PubMed]
- Rex, D.K.; Ahnen, D.J.; Baron, J.A.; Batts, K.P.; Burke, C.A.; Burt, R.W.; Goldblum, J.R.; Guillem, J.G.; Kahi, C.J.; Kalady, M.F.; et al. Serrated Lesions of the Colorectum: Review and Recommendations from an Expert Panel. Am. J. Gastroenterol. 2012, 107, 1315–1330. [Google Scholar] [CrossRef]
- Patai, Á.V.; Molnár, B.; Tulassay, Z.; Sipos, F. Serrated pathway: Alternative route to colorectal cancer. World J. Gastroenterol. 2013, 19, 607–615. [Google Scholar] [CrossRef]
- Leggett, B.; Whitehall, V. Role of the serrated pathway in colorectal cancer pathogenesis. Gastroenterology 2010, 138, 2088–2100. [Google Scholar] [CrossRef]
- Bettington, M.; Walker, N.; Clouston, A.; Brown, I.; Leggett, B.; Whitehall, V. The serrated pathway to colorectal carcinoma: Current concepts and challenges. Histopathology 2013, 62, 367–386. [Google Scholar] [CrossRef]
- Longacre, T.A.; Fenoglio-Preiser, C.M. Mixed hyperplastic adenomatous polyps/serrated adenomas. A distinct form of colorectal neoplasia. Am. J. Surg. Pathol. 1990, 14, 524–537. [Google Scholar] [CrossRef] [PubMed]
- East, J.E.; Atkin, W.S.; Bateman, A.C.; Clark, S.K.; Dolwani, S.; Ket, S.N.; Leedham, S.J.; Phull, P.S.; Rutter, M.D.; Shepherd, N.A.; et al. British Society of Gastroenterology position statement on serrated polyps in the colon and rectum. Gut 2017, 66, 1181–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, R.K.; Bettington, M.; Srivastava, A.; Rosty, C. An update on the morphology and molecular pathology of serrated colorectal polyps and associated carcinomas. Mod. Pathol. 2019, 1. [Google Scholar] [CrossRef] [PubMed]
- Kahi, C.J. Screening Relevance of Sessile Serrated Polyps. Clin. Endosc. 2019, 52. [Google Scholar] [CrossRef] [PubMed]
- Hamoudah, T.; Ma, K.; Esteban, M.; Hayat, W.; Berger, D.; Mahon, B.; Jakate, S.; Melson, J. Patients with small and diminutive proximal hyperplastic polyps have higher rates of synchronous advanced neoplasia compared with patients without serrated lesions. Gastrointest. Endosc. 2018, 87, 1518–1526. [Google Scholar] [CrossRef]
- Mojarad, E.N.; Kuppen, P.J.; Aghdaei, H.A.; Zali, M.R. The CpG island methylator phenotype (CIMP) in colorectal cancer. Gastroenterol. Hepatol. Bed Bench 2013, 6, 120–128. [Google Scholar]
- Barault, L.; Charon-Barra, C.; Jooste, V.; de la Vega, M.F.; Martin, L.; Roignot, P.; Rat, P.; Bouvier, A.-M.; Laurent-Puig, P.; Faivre, J.; et al. Hypermethylator phenotype in sporadic colon cancer: Study on a population-based series of 582 cases. Cancer Res. 2008, 68, 8541–8546. [Google Scholar] [CrossRef]
- Borowsky, J.; Dumenil, T.; Bettington, M.; Pearson, S.-A.; Bond, C.; Fennell, L.; Liu, C.; McKeone, D.; Rosty, C.; Brown, I.; et al. The role of APC in WNT pathway activation in serrated neoplasia. Mod. Pathol. 2018, 31, 495–504. [Google Scholar] [CrossRef]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The Consensus Molecular Subtypes of Colorectal Cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef]
- Bae, J.M.; Kim, J.H.; Kwak, Y.; Lee, D.-W.; Cha, Y.; Wen, X.; Lee, T.H.; Cho, N.-Y.; Jeong, S.-Y.; Park, K.J.; et al. Distinct clinical outcomes of two CIMP-positive colorectal cancer subtypes based on a revised CIMP classification system. Br. J. Cancer 2017, 116, 1012–1020. [Google Scholar] [CrossRef]
- Fennell, L.; Dumenil, T.; Wockner, L.; Hartel, G.; Nones, K.; Bond, C.; Borowsky, J.; Liu, C.; McKeone, D.; Bowdler, L.; et al. Integrative Genome-Scale DNA Methylation Analysis of a Large and Unselected Cohort Reveals Five Distinct Subtypes of Colorectal Adenocarcinomas. Cell. Mol. Gastroenterol. Hepatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kahi, C.J.; Hewett, D.G.; Rex, D.K. Relationship of non-polypoid colorectal neoplasms to quality of colonoscopy. Gastrointest. Endosc. Clin. N. Am. 2010, 20, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Kahi, C.J.; Hewett, D.G.; Norton, D.L.; Eckert, G.J.; Rex, D.K. Prevalence and variable detection of proximal colon serrated polyps during screening colonoscopy. Clin. Gastroenterol. Hepatol. 2011, 9, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Torlakovic, E.; Skovlund, E.; Snover, D.C.; Torlakovic, G.; Nesland, J.M. Morphologic reappraisal of serrated colorectal polyps. Am. J. Surg. Pathol. 2003, 27, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Bosman, F.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumours of the Digestive System, 4th ed.; World Health Organization, International Agency for Research on Cancer (IARC): Lyon, France, 2010. [Google Scholar]
- Cassese, G.; Amendola, A.; Maione, F.; Giglio, M.C.; Pagano, G.; Milone, M.; Aprea, G.; Luglio, G.; De Palma, G.D. Serrated Lesions of the Colon-Rectum: A Focus on New Diagnostic Tools and Current Management. Gastroenterol. Res. Pract. 2019, 2019, 9179718. [Google Scholar] [CrossRef] [PubMed]
- Aust, D.E.; Baretton, G.B.; Members of the Working Group GI-Pathology of the German Society of Pathology. Serrated polyps of the colon and rectum (hyperplastic polyps, sessile serrated adenomas, traditional serrated adenomas, and mixed polyps)-proposal for diagnostic criteria. Virchows Arch. Int. J. Pathol. 2010, 457, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Yamane, L.; Scapulatempo-Neto, C.; Reis, R.M.; Guimarães, D.P. Serrated pathway in colorectal carcinogenesis. World J. Gastroenterol. 2014, 20, 2634–2640. [Google Scholar] [CrossRef]
- IJspeert, J.E.G.; Vermeulen, L.; Meijer, G.A.; Dekker, E. Serrated neoplasia—role in colorectal carcinogenesis and clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 401–409. [Google Scholar] [CrossRef]
- Murakami, T.; Sakamoto, N.; Nagahara, A. Endoscopic diagnosis of sessile serrated adenoma/polyp with and without dysplasia/carcinoma. World J. Gastroenterol. 2018, 24, 3250–3259. [Google Scholar] [CrossRef]
- Torlakovic, E.; Snover, D.C. Serrated adenomatous polyposis in humans. Gastroenterology 1996, 110, 748–755. [Google Scholar] [CrossRef]
- Yantiss, R.K.; Oh, K.Y.; Chen, Y.-T.; Redston, M.; Odze, R.D. Filiform serrated adenomas: A clinicopathologic and immunophenotypic study of 18 cases. Am. J. Surg. Pathol. 2007, 31, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Kalimuthu, S.N.; Chelliah, A.; Chetty, R. From traditional serrated adenoma to tubulovillous adenoma and beyond. World J. Gastrointest. Oncol. 2016, 8, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Samaranayake, M.; Pradhan, S. Epigenetic mechanisms in mammals. Cell. Mol. Life Sci. CMLS 2009, 66, 596–612. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.R.; Yarychkivska, O.; Boulard, M.; Bestor, T.H. DNA methylation and DNA methyltransferases. Epigenetics Chromatin 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Robertson, K.D. DNA methylation and human disease. Nat. Rev. Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef]
- Yokoi, K.; Harada, H.; Yokota, K.; Ishii, S.; Tanaka, T.; Nishizawa, N.; Shimazu, M.; Kojo, K.; Miura, H.; Yamanashi, T.; et al. Epigenetic Status of CDO1 Gene May Reflect Chemosensitivity in Colon Cancer with Postoperative Adjuvant Chemotherapy. Ann. Surg. Oncol. 2019, 26, 406–414. [Google Scholar] [CrossRef]
- Xue, W.; Wu, X.; Wang, F.; Han, P.; Cui, B. Genome-wide methylation analysis identifies novel prognostic methylation markers in colon adenocarcinoma. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 108, 288–296. [Google Scholar] [CrossRef]
- Kokelaar, R.F.; Jones, H.; Beynon, J.; Evans, M.E.; Harris, D.A. Meta-analysis of the prognostic value of CpG island methylator phenotype in rectal cancer. Int. J. Colorectal Dis. 2018, 33, 995–1000. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Bettington, M.L.; Walker, N.I.; Dwine, J.; Hartel, G.F.; Leggett, B.A.; Whitehall, V.L.J. CpG Island Methylation in Sessile Serrated Adenomas Increases With Age, Indicating Lower Risk of Malignancy in Young Patients. Gastroenterology 2018, 155, 1362–1365. [Google Scholar] [CrossRef]
- Ogino, S.; Cantor, M.; Kawasaki, T.; Brahmandam, M.; Kirkner, G.J.; Weisenberger, D.J.; Campan, M.; Laird, P.W.; Loda, M.; Fuchs, C.S. CpG island methylator phenotype (CIMP) of colorectal cancer is best characterised by quantitative DNA methylation analysis and prospective cohort studies. Gut 2006, 55, 1000–1006. [Google Scholar] [CrossRef]
- Noffsinger, A.E. Serrated polyps and colorectal cancer: New pathway to malignancy. Annu. Rev. Pathol. 2009, 4, 343–364. [Google Scholar] [CrossRef] [PubMed]
- Advani, S.M.; Advani, P.; DeSantis, S.M.; Brown, D.; VonVille, H.M.; Lam, M.; Loree, J.M.; Sarshekeh, A.M.; Bressler, J.; Lopez, D.S.; et al. Clinical, Pathological, and Molecular Characteristics of CpG Island Methylator Phenotype in Colorectal Cancer: A Systematic Review and Meta-analysis. Transl. Oncol. 2018, 11, 1188–1201. [Google Scholar] [CrossRef]
- Ogino, S.; Odze, R.D.; Kawasaki, T.; Brahmandam, M.; Kirkner, G.J.; Laird, P.W.; Loda, M.; Fuchs, C.S. Correlation of pathologic features with CpG island methylator phenotype (CIMP) by quantitative DNA methylation analysis in colorectal carcinoma. Am. J. Surg. Pathol. 2006, 30, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Toyota, M.; Kondo, Y.; Lin, E.; Zhang, L.; Guo, Y.; Hernandez, N.S.; Chen, X.; Ahmed, S.; Konishi, K.; et al. Integrated genetic and epigenetic analysis identifies three different subclasses of colon cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 18654–18659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisenberger, D.J.; Levine, A.J.; Long, T.I.; Buchanan, D.D.; Walters, R.; Clendenning, M.; Rosty, C.; Joshi, A.D.; Stern, M.C.; LeMarchand, L.; et al. Association of the Colorectal CpG Island Methylator Phenotype with molecular features, risk factors and family history. Cancer Epidemiol. Biomark. Prev. 2015, 24, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Ou, J.; Hutchinson, L.; Green, M.R. The BRAF oncoprotein functions through the transcriptional repressor MAFG to mediate the CpG Island Methylator phenotype. Mol. Cell 2014, 55, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Bond, C.E.; Liu, C.; Kawamata, F.; McKeone, D.M.; Fernando, W.; Jamieson, S.; Pearson, S.-A.; Kane, A.; Woods, S.L.; Lannagan, T.R.M.; et al. Oncogenic BRAF mutation induces DNA methylation changes in a murine model for human serrated colorectal neoplasia. Epigenetics 2018, 13, 40–48. [Google Scholar] [CrossRef]
- Tao, Y.; Kang, B.; Petkovich, D.A.; Bhandari, Y.R.; In, J.; Stein-O’Brien, G.; Kong, X.; Xie, W.; Zachos, N.; Maegawa, S.; et al. Aging-like Spontaneous Epigenetic Silencing Facilitates Wnt Activation, Stemness, and BrafV600E-Induced Tumorigenesis. Cancer Cell 2019, 35, 315–328. [Google Scholar] [CrossRef]
- Yagi, K.; Akagi, K.; Hayashi, H.; Nagae, G.; Tsuji, S.; Isagawa, T.; Midorikawa, Y.; Nishimura, Y.; Sakamoto, H.; Seto, Y.; et al. Three DNA methylation epigenotypes in human colorectal cancer. Clin. Cancer Res. 2010, 16, 21–33. [Google Scholar] [CrossRef]
- Ogino, S.; Kawasaki, T.; Kirkner, G.J.; Kraft, P.; Loda, M.; Fuchs, C.S. Evaluation of markers for CpG island methylator phenotype (CIMP) in colorectal cancer by a large population-based sample. J. Mol. Diagn. 2007, 9, 305–314. [Google Scholar] [CrossRef]
- Ogino, S.; Kawasaki, T.; Kirkner, G.J.; Loda, M.; Fuchs, C.S. CpG island methylator phenotype-low (CIMP-low) in colorectal cancer: Possible associations with male sex and KRAS mutations. J. Mol. Diagn. 2006, 8, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, N.; Norrie, M.; Cheong, K.; Mokany, E.; Ku, S.-L.; Meagher, A.; O’Connor, T.; Ward, R. CpG island methylation in sporadic colorectal cancers and its relationship to microsatellite instability. Gastroenterology 2002, 122, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Ahuja, N.; Ohe-Toyota, M.; Herman, J.G.; Baylin, S.B.; Issa, J.P. CpG island methylator phenotype in colorectal cancer. Proc. Natl. Acad. Sci. USA 1999, 96, 8681–8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisenberger, D.J.; Siegmund, K.D.; Campan, M.; Young, J.; Long, T.I.; Faasse, M.A.; Kang, G.H.; Widschwendter, M.; Weener, D.; Buchanan, D.; et al. CpG island methylator phenotype underlies sporadic microsatellite instability and is tightly associated with BRAF mutation in colorectal cancer. Nat. Genet. 2006, 38, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Ogino, S.; Kawasaki, T.; Kirkner, G.J.; Suemoto, Y.; Meyerhardt, J.A.; Fuchs, C.S. Molecular correlates with MGMT promoter methylation and silencing support CpG island methylator phenotype-low (CIMP-low) in colorectal cancer. Gut 2007, 56, 1564–1571. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.R.; Orjuela, S.; Martinho Oliveira, A.; Cereatti, F.; Sauter, M.; Heinrich, H.; Tanzi, G.; Weber, A.; Komminoth, P.; Vavricka, S.; et al. The proto CpG island methylator phenotype of sessile serrated adenomas/polyps. Epigenetics 2018, 13, 1088–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperiale, T.F.; Ransohoff, D.F.; Itzkowitz, S.H. Multitarget stool DNA testing for colorectal-cancer screening. N. Engl. J. Med. 2014, 371, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Kadiyska, T.; Nossikoff, A. Stool DNA methylation assays in colorectal cancer screening. World J. Gastroenterol. 2015, 21, 10057–10061. [Google Scholar] [CrossRef]
- Hinoue, T.; Weisenberger, D.J.; Lange, C.P.E.; Shen, H.; Byun, H.-M.; Van Den Berg, D.; Malik, S.; Pan, F.; Noushmehr, H.; van Dijk, C.M.; et al. Genome-scale analysis of aberrant DNA methylation in colorectal cancer. Genome Res. 2012, 22, 271–282. [Google Scholar] [CrossRef]
- Yang, S.; Farraye, F.A.; Mack, C.; Posnik, O.; O’Brien, M.J. BRAF and KRAS Mutations in hyperplastic polyps and serrated adenomas of the colorectum: Relationship to histology and CpG island methylation status. Am. J. Surg. Pathol. 2004, 28, 1452–1459. [Google Scholar] [CrossRef]
- Spring, K.J.; Zhao, Z.Z.; Karamatic, R.; Walsh, M.D.; Whitehall, V.L.J.; Pike, T.; Simms, L.A.; Young, J.; James, M.; Montgomery, G.W.; et al. High Prevalence of Sessile Serrated Adenomas with BRAF Mutations: A Prospective Study of Patients Undergoing Colonoscopy. Gastroenterology 2006, 131, 1400–1407. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.A.; Hahn, H.P.; Shahsafaei, A.; Odze, R.D. MUC expression in hyperplastic and serrated colonic polyps: Lack of specificity of MUC6. Am. J. Surg. Pathol. 2011, 35, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Renaud, F.; Mariette, C.; Vincent, A.; Wacrenier, A.; Maunoury, V.; Leclerc, J.; Coppin, L.; Crépin, M.; Seuningen, I.V.; Leteurtre, E.; et al. The serrated neoplasia pathway of colorectal tumors: Identification of MUC5AC hypomethylation as an early marker of polyps with malignant potential. Int. J. Cancer 2016, 138, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Mitomi, H.; Saito, T.; Takahashi, M.; Sakamoto, N.; Fukui, N.; Yao, T.; Watanabe, S. Distinct WNT/β-catenin signaling activation in the serrated neoplasia pathway and the adenoma-carcinoma sequence of the colorectum. Mod. Pathol. 2015, 28, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Ashktorab, H.; Delker, D.; Kanth, P.; Goel, A.; Carethers, J.M.; Brim, H. Molecular Characterization of sessile serrated adenoma/polyps from a large African American cohort. Gastroenterology 2019. [Google Scholar] [CrossRef]
- Beggs, A.D.; Jones, A.; Shepherd, N.; Arnaout, A.; Finlayson, C.; Abulafi, A.M.; Morton, D.G.; Matthews, G.M.; Hodgson, S.V.; Tomlinson, I.P.M. Loss of Expression and Promoter Methylation of SLIT2 Are Associated with Sessile Serrated Adenoma Formation. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef]
- Caruso, M.; Moore, J.; Goodall, G.J.; Thomas, M.; Phillis, S.; Tyskin, A.; Cheetham, G.; Lerda, N.; Takahashi, H.; Ruszkiewicz, A. Over-expression of cathepsin E and trefoil factor 1 in sessile serrated adenomas of the colorectum identified by gene expression analysis. Virchows Arch. Int. J. Pathol. 2009, 454, 291–302. [Google Scholar] [CrossRef]
- Gonzalo, D.H.; Lai, K.K.; Shadrach, B.; Goldblum, J.R.; Bennett, A.E.; Downs-Kelly, E.; Liu, X.; Henricks, W.; Patil, D.T.; Carver, P.; et al. Gene expression profiling of serrated polyps identifies annexin A10 as a marker of a sessile serrated adenoma/polyp. J. Pathol. 2013, 230, 420–429. [Google Scholar] [CrossRef]
- Kanth, P.; Bronner, M.P.; Boucher, K.M.; Burt, R.W.; Neklason, D.W.; Hagedorn, C.H.; Delker, D.A. Gene Signature in Sessile Serrated Polyps Identifies Colon Cancer Subtype. Cancer Prev. Res. Phila. Pa 2016, 9, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.M.; Kim, J.H.; Rhee, Y.-Y.; Cho, N.-Y.; Kim, T.-Y.; Kang, G.H. Annexin A10 expression in colorectal cancers with emphasis on the serrated neoplasia pathway. World J. Gastroenterol. 2015, 21, 9749–9757. [Google Scholar] [CrossRef]
- Cui, M.; Awadallah, A.; Liu, W.; Zhou, L.; Xin, W. Loss of Hes1 Differentiates Sessile Serrated Adenoma/Polyp from Hyperplastic Polyp. Am. J. Surg. Pathol. 2016, 40, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Delker, D.A.; McGettigan, B.M.; Kanth, P.; Pop, S.; Neklason, D.W.; Bronner, M.P.; Burt, R.W.; Hagedorn, C.H. RNA Sequencing of Sessile Serrated Colon Polyps Identifies Differentially Expressed Genes and Immunohistochemical Markers. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Rahmatallah, Y.; Khaidakov, M.; Lai, K.K.; Goyne, H.E.; Lamps, L.W.; Hagedorn, C.H.; Glazko, G. Platform-independent gene expression signature differentiates sessile serrated adenomas/polyps and hyperplastic polyps of the colon. BMC Med. Genomics 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Mikhaleva, L.M.; Vandysheva, R.A.; Shakhpazyan, N.K.; Fedorov, E.D.; Biryukov, A.E.; Midiber, K.Y.; Pechnikova, V.V. [Comparative assessment of the expression of Muc 2, Muc 5AC, and Muc 6 in serrated neoplasms of the colon]. Arkh. Patol. 2019, 81, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Owens, S.R.; Chiosea, S.I.; Kuan, S.-F. Selective expression of gastric mucin MUC6 in colonic sessile serrated adenoma but not in hyperplastic polyp aids in morphological diagnosis of serrated polyps. Mod. Pathol. 2008, 21, 660–669. [Google Scholar] [CrossRef]
- Travaglino, A.; D’Armiento, F.P.; Cassese, G.; Campanino, M.R.; Borrelli, G.; Pignatiello, S.; Luglio, G.; Maione, F.; De Palma, G.D.; D’Armiento, M. Clinico-pathological factors associated with BRAF-V600E mutation in colorectal serrated adenomas. Histopathology 2019. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Akazawa, Y.; Yatagai, N.; Hiromoto, T.; Sasahara, N.; Saito, T.; Sakamoto, N.; Nagahara, A.; Yao, T. Molecular characterization of sessile serrated adenoma/polyps with dysplasia/carcinoma based on immunohistochemistry, next-generation sequencing, and microsatellite instability testing: A case series study. Diagn. Pathol. 2018, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Yozu, M.; Kem, M.; Cenaj, O.; Mino-Kenudson, M.; Odze, R.D.; Misdraji, J. Loss of expression of MLH1 in non-dysplastic crypts is a harbinger of neoplastic progression in sessile serrated adenomas/polyps. Histopathology 2019. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Yamashita, S.; Yoshida, H.; Taniguchi, H.; Ushijima, T.; Yamada, T.; Saito, Y.; Ochiai, A.; Sekine, S.; Hiraoka, N. WNT Pathway Gene Mutations Are Associated with the Presence of Dysplasia in Colorectal Sessile Serrated Adenoma/Polyps. Am. J. Surg. Pathol. 2017, 41, 1188–1197. [Google Scholar] [CrossRef]
- Bond, C.E.; McKeone, D.M.; Kalimutho, M.; Bettington, M.L.; Pearson, S.-A.; Dumenil, T.D.; Wockner, L.F.; Burge, M.; Leggett, B.A.; Whitehall, V.L.J. RNF43 and ZNRF3 are commonly altered in serrated pathway colorectal tumorigenesis. Oncotarget 2016, 7, 70589–70600. [Google Scholar] [CrossRef]
- Koinuma, K.; Yamashita, Y.; Liu, W.; Hatanaka, H.; Kurashina, K.; Wada, T.; Takada, S.; Kaneda, R.; Choi, Y.L.; Fujiwara, S.-I.; et al. Epigenetic silencing of AXIN2 in colorectal carcinoma with microsatellite instability. Oncogene 2006, 25, 139–146. [Google Scholar] [CrossRef]
- Liu, N.Q.; ter Huurne, M.; Nguyen, L.N.; Peng, T.; Wang, S.-Y.; Studd, J.B.; Joshi, O.; Ongen, H.; Bramsen, J.B.; Yan, J.; et al. The non-coding variant rs1800734 enhances DCLK3 expression through long-range interaction and promotes colorectal cancer progression. Nat. Commun. 2017, 8, 14418. [Google Scholar] [CrossRef] [Green Version]
- Fennell, L.J.; Jamieson, S.; McKeone, D.; Corish, T.; Rohdmann, M.; Furner, T.; Bettington, M.; Liu, C.; Kawamata, F.; Bond, C.; et al. MLH1-93 G/a polymorphism is associated with MLH1 promoter methylation and protein loss in dysplastic sessile serrated adenomas with BRAFV600E mutation. BMC Cancer 2018, 18, 35. [Google Scholar] [CrossRef]
- Torlakovic, E.E.; Gomez, J.D.; Driman, D.K.; Parfitt, J.R.; Wang, C.; Benerjee, T.; Snover, D.C. Sessile serrated adenoma (SSA) vs. traditional serrated adenoma (TSA). Am. J. Surg. Pathol. 2008, 32, 21–29. [Google Scholar] [CrossRef]
- Bettington, M.L.; Walker, N.I.; Rosty, C.; Brown, I.S.; Clouston, A.D.; McKeone, D.M.; Pearson, S.-A.; Klein, K.; Leggett, B.A.; Whitehall, V.L.J. A clinicopathological and molecular analysis of 200 traditional serrated adenomas. Mod. Pathol. 2015, 28, 414–427. [Google Scholar] [CrossRef]
- Whitehall, V.L.J.; Walsh, M.D.; Young, J.; Leggett, B.A.; Jass, J.R. Methylation of O–6-Methylguanine DNA Methyltransferase Characterizes a Subset of Colorectal Cancer with Low-level DNA Microsatellite Instability. Cancer Res. 2001, 61, 827–830. [Google Scholar]
- Hashimoto, T.; Ogawa, R.; Yoshida, H.; Taniguchi, H.; Kojima, M.; Saito, Y.; Sekine, S. Acquisition of WNT Pathway Gene Alterations Coincides With the Transition From Precursor Polyps to Traditional Serrated Adenomas. Am. J. Surg. Pathol. 2019, 43, 132–139. [Google Scholar] [CrossRef]
- Sekine, S.; Yamashita, S.; Tanabe, T.; Hashimoto, T.; Yoshida, H.; Taniguchi, H.; Kojima, M.; Shinmura, K.; Saito, Y.; Hiraoka, N.; et al. Frequent PTPRK-RSPO3 fusions and RNF43 mutations in colorectal traditional serrated adenoma. J. Pathol. 2016, 239, 133–138. [Google Scholar] [CrossRef]
- Tsai, J.-H.; Jeng, Y.-M.; Yuan, C.-T.; Lin, Y.-L.; Cheng, M.-L.; Liau, J.-Y. Traditional Serrated Pathway-associated Colorectal Carcinoma: Morphologic Reappraisal of Serrated Morphology, Tumor Budding, and Identification of Frequent PTEN Alterations. Am. J. Surg. Pathol. 2019, 43. [Google Scholar] [CrossRef]
- Aoki, H.; Yamamoto, E.; Takasawa, A.; Niinuma, T.; Yamano, H.-O.; Harada, T.; Matsushita, H.-O.; Yoshikawa, K.; Takagi, R.; Harada, E.; et al. Epigenetic silencing of SMOC1 in traditional serrated adenoma and colorectal cancer. Oncotarget 2017, 9, 4707–4721. [Google Scholar] [CrossRef] [Green Version]
- Naito, T.; Nosho, K.; Ito, M.; Igarashi, H.; Mitsuhashi, K.; Yoshii, S.; Aoki, H.; Nomura, M.; Sukawa, Y.; Yamamoto, E.; et al. IGF2 differentially methylated region hypomethylation in relation to pathological and molecular features of serrated lesions. World J. Gastroenterol. 2014, 20, 10050–10061. [Google Scholar] [CrossRef]
- Kerachian, M.A.; Kerachian, M. Long interspersed nucleotide element-1 (LINE-1) methylation in colorectal cancer. Clin. Chim. Acta Int. J. Clin. Chem. 2019, 488, 209–214. [Google Scholar] [CrossRef]
- Nagai, Y.; Sunami, E.; Yamamoto, Y.; Hata, K.; Okada, S.; Murono, K.; Yasuda, K.; Otani, K.; Nishikawa, T.; Tanaka, T.; et al. LINE-1 hypomethylation status of circulating cell-free DNA in plasma as a biomarker for colorectal cancer. Oncotarget 2017, 8, 11906–11916. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.-H.; Liau, J.-Y.; Lin, Y.-L.; Lin, L.-I.; Cheng, Y.-C.; Cheng, M.-L.; Jeng, Y.-M. Traditional serrated adenoma has two pathways of neoplastic progression that are distinct from the sessile serrated pathway of colorectal carcinogenesis. Mod. Pathol. 2014, 27, 1375–1385. [Google Scholar] [CrossRef] [Green Version]
- Wajapeyee, N.; Serra, R.W.; Zhu, X.; Mahalingam, M.; Green, M.R. Role for IGFBP7 in senescence induction by BRAF. Cell 2010, 141, 746–747. [Google Scholar] [CrossRef]
- Kriegl, L.; Neumann, J.; Vieth, M.; Greten, F.R.; Reu, S.; Jung, A.; Kirchner, T. Up and downregulation of p16(Ink4a) expression in BRAF-mutated polyps/adenomas indicates a senescence barrier in the serrated route to colon cancer. Mod. Pathol. 2011, 24, 1015–1022. [Google Scholar] [CrossRef]
- Carragher, L.A.S.; Snell, K.R.; Giblett, S.M.; Aldridge, V.S.S.; Patel, B.; Cook, S.J.; Winton, D.J.; Marais, R.; Pritchard, C.A. V600EBraf induces gastrointestinal crypt senescence and promotes tumour progression through enhanced CpG methylation of p16INK4a. EMBO Mol. Med. 2010, 2, 458–471. [Google Scholar] [CrossRef]
- García-Solano, J.; Turpin, M.C.; Torres-Moreno, D.; Huertas-López, F.; Tuomisto, A.; Mäkinen, M.J.; Conesa, A.; Conesa-Zamora, P. Two histologically colorectal carcinomas subsets from the serrated pathway show different methylome signatures and diagnostic biomarkers. Clin. Epigenetics 2018, 10, 141. [Google Scholar] [CrossRef]
- Mäkinen, M.J. Colorectal serrated adenocarcinoma. Histopathology 2007, 50, 131–150. [Google Scholar] [CrossRef]
- Jass, J.R.; Smith, M. Sialic acid and epithelial differentiation in colorectal polyps and cancer—A morphological, mucin and lectin histochemical study. Pathology (Phila.) 1992, 24, 233–242. [Google Scholar] [CrossRef]
- García-Solano, J.; Pérez-Guillermo, M.; Conesa-Zamora, P.; Acosta-Ortega, J.; Trujillo-Santos, J.; Cerezuela-Fuentes, P.; Mäkinen, M.J. Clinicopathologic study of 85 colorectal serrated adenocarcinomas: Further insights into the full recognition of a new subset of colorectal carcinoma. Hum. Pathol. 2010, 41, 1359–1368. [Google Scholar] [CrossRef]
- Stefanius, K.; Ylitalo, L.; Tuomisto, A.; Kuivila, R.; Kantola, T.; Sirniö, P.; Karttunen, T.J.; Mäkinen, M.J. Frequent mutations of KRAS in addition to BRAF in colorectal serrated adenocarcinoma. Histopathology 2011, 58, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Laiho, P.; Kokko, A.; Vanharanta, S.; Salovaara, R.; Sammalkorpi, H.; Järvinen, H.; Mecklin, J.-P.; Karttunen, T.J.; Tuppurainen, K.; Davalos, V.; et al. Serrated carcinomas form a subclass of colorectal cancer with distinct molecular basis. Oncogene 2007, 26, 312–320. [Google Scholar] [CrossRef]
- Tuomisto, A.; García-Solano, J.; Sirniö, P.; Väyrynen, J.; Pérez-Guillermo, M.; Mäkinen, M.J.; Conesa-Zamora, P. HIF-1α expression and high microvessel density are characteristic features in serrated colorectal cancer. Virchows Arch. Int. J. Pathol. 2016, 469, 395–404. [Google Scholar] [CrossRef]
- García-Solano, J.; García-Solano, M.E.; Torres-Moreno, D.; Carbonell, P.; Trujillo-Santos, J.; Pérez-Guillermo, M.; Conesa-Zamora, P. Biomarkers for the identification of precursor polyps of colorectal serrated adenocarcinomas. Cell. Oncol. Dordr. 2016, 39, 243–252. [Google Scholar] [CrossRef]
- Lian, H.; Jia, X.; Shi, N.; Xie, S.; Wang, J.; Wang, W.; Ma, F.; Liu, H.; Wang, A.; Cheng, X.; et al. Notch signaling promotes serrated neoplasia pathway in colorectal cancer through epigenetic modification of EPHB2 and EPHB4. Cancer Manag. Res. 2018, 10, 6129–6141. [Google Scholar] [CrossRef]
- Alazzouzi, H.; Davalos, V.; Kokko, A.; Domingo, E.; Woerner, S.M.; Wilson, A.J.; Konrad, L.; Laiho, P.; Espín, E.; Armengol, M.; et al. Mechanisms of inactivation of the receptor tyrosine kinase EPHB2 in colorectal tumors. Cancer Res. 2005, 65, 10170–10173. [Google Scholar] [CrossRef]
- Conesa-Zamora, P.; García-Solano, J.; García-García, F.; Turpin, M.D.C.; Trujillo-Santos, J.; Torres-Moreno, D.; Oviedo-Ramírez, I.; Carbonell-Muñoz, R.; Muñoz-Delgado, E.; Rodriguez-Braun, E.; et al. Expression profiling shows differential molecular pathways and provides potential new diagnostic biomarkers for colorectal serrated adenocarcinoma. Int. J. Cancer 2013, 132, 297–307. [Google Scholar] [CrossRef]
- Conesa-Zamora, P.; García-Solano, J.; Turpin, M.D.C.; Sebastián-León, P.; Torres-Moreno, D.; Estrada, E.; Tuomisto, A.; Wilce, J.; Mäkinen, M.J.; Pérez-Guillermo, M.; et al. Methylome profiling reveals functions and genes which are differentially methylated in serrated compared to conventional colorectal carcinoma. Clin. Epigenetics 2015, 7, 101. [Google Scholar] [CrossRef]
- García-Solano, J.; Del Carmen Turpin, M.; García-García, F.; Carbonell-Muñoz, R.; Torres-Moreno, D.; Conesa, A.; Conesa-Zamora, P. Differences in gene expression profiling and biomarkers between histological colorectal carcinoma subsets from the serrated pathway. Histopathology 2019. [Google Scholar] [CrossRef]
- Hashimoto, T.; Tanaka, Y.; Ogawa, R.; Mori, T.; Yoshida, H.; Taniguchi, H.; Hiraoka, N.; Kojima, M.; Oono, Y.; Saito, Y.; et al. Superficially serrated adenoma: A proposal for a novel subtype of colorectal serrated lesion. Mod. Pathol. 2018, 31, 1588–1598. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Tsikitis, V.L.; Potter, A.; Mori, M.; Buckmeier, J.A.; Preece, C.R.; Harrington, C.A.; Bartley, A.N.; Bhattacharyya, A.K.; Hamilton, S.R.; Lance, M.P.; et al. MicroRNA Signatures of Colonic Polyps on Screening and Histology. Cancer Prev. Res. Phila. Pa 2016, 9, 942–949. [Google Scholar] [CrossRef]
- Kanth, P.; Hazel, M.W.; Boucher, K.M.; Yang, Z.; Wang, L.; Bronner, M.P.; Boylan, K.E.; Burt, R.W.; Westover, M.; Neklason, D.W.; et al. Small RNA sequencing of sessile serrated polyps identifies microRNA profile associated with colon cancer. Genes. Chromosomes Cancer 2019, 58, 23–33. [Google Scholar] [CrossRef]
- James de Bony, E.; Bizet, M.; Van Grembergen, O.; Hassabi, B.; Calonne, E.; Putmans, P.; Bontempi, G.; Fuks, F. Comprehensive identification of long noncoding RNAs in colorectal cancer. Oncotarget 2018, 9, 27605–27629. [Google Scholar] [Green Version]
- Aoki, H.; Nosho, K.; Igarashi, H.; Ito, M.; Mitsuhashi, K.; Naito, T.; Yamamoto, E.; Tanuma, T.; Nomura, M.; Maguchi, H.; et al. MicroRNA-31 expression in colorectal serrated pathway progression. World J. Gastroenterol. 2014, 20, 12346–12349. [Google Scholar] [CrossRef]
- Slattery, M.L.; Herrick, J.S.; Wolff, R.K.; Mullany, L.E.; Stevens, J.R.; Samowitz, W. The miRNA Landscape of Colorectal Polyps. Genes. Chromosomes Cancer 2017, 56, 347–353. [Google Scholar] [CrossRef]
- Chen, H.; Xu, J.; Hong, J.; Tang, R.; Zhang, X.; Fang, J.-Y. Long noncoding RNA profiles identify five distinct molecular subtypes of colorectal cancer with clinical relevance. Mol. Oncol. 2014, 8, 1393–1403. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [Green Version]
- D’Argenio, V.; Salvatore, F. The role of the gut microbiome in the healthy adult status. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 451, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef]
- Zitvogel, L.; Ayyoub, M.; Routy, B.; Kroemer, G. Microbiome and Anticancer Immunosurveillance. Cell 2016, 165, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Kroemer, G.; Zitvogel, L. Cancer immunotherapy in 2017: The breakthrough of the microbiota. Nat. Rev. Immunol. 2018, 18, 87–88. [Google Scholar] [CrossRef]
- Mangifesta, M.; Mancabelli, L.; Milani, C.; Gaiani, F.; de’Angelis, N.; de’Angelis, G.L.; van Sinderen, D.; Ventura, M.; Turroni, F. Mucosal microbiota of intestinal polyps reveals putative biomarkers of colorectal cancer. Sci. Rep. 2018, 8, 13974. [Google Scholar] [CrossRef]
- Jahani-Sherafat, S.; Alebouyeh, M.; Moghim, S.; Ahmadi Amoli, H.; Ghasemian-Safaei, H. Role of gut microbiota in the pathogenesis of colorectal cancer; a review article. Gastroenterol. Hepatol. Bed Bench 2018, 11, 101–109. [Google Scholar]
- Chen, G.Y. The Role of the Gut Microbiome in Colorectal Cancer. Clin. Colon Rectal Surg. 2018, 31, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.A.; Dominianni, C.; Shapiro, J.A.; Church, T.R.; Wu, J.; Miller, G.; Yuen, E.; Freiman, H.; Lustbader, I.; Salik, J.; et al. The gut microbiota in conventional and serrated precursors of colorectal cancer. Microbiome 2016, 4. [Google Scholar] [CrossRef]
- Yoon, H.; Kim, N.; Park, J.H.; Kim, Y.S.; Lee, J.; Kim, H.W.; Choi, Y.J.; Shin, C.M.; Park, Y.S.; Lee, D.H.; et al. Comparisons of Gut Microbiota Among Healthy Control, Patients with Conventional Adenoma, Sessile Serrated Adenoma, and Colorectal Cancer. J. Cancer Prev. 2017, 22, 108–114. [Google Scholar] [CrossRef]
- Holt, R.A.; Cochrane, K. Tumor Potentiating Mechanisms of Fusobacterium nucleatum, A Multifaceted Microbe. Gastroenterology 2017, 152, 694–696. [Google Scholar] [CrossRef]
- Shang, F.-M.; Liu, H.-L. Fusobacterium nucleatum and colorectal cancer: A review. World J. Gastrointest. Oncol. 2018, 10, 71–81. [Google Scholar] [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Ma, C.-T.; Luo, H.-S.; Gao, F.; Tang, Q.-C.; Chen, W. Fusobacterium nucleatum promotes the progression of colorectal cancer by interacting with E-cadherin. Oncol. Lett. 2018, 16, 2606–2612. [Google Scholar] [CrossRef]
- Lee, D.-W.; Han, S.-W.; Kang, J.-K.; Bae, J.M.; Kim, H.-P.; Won, J.-K.; Jeong, S.-Y.; Park, K.J.; Kang, G.H.; Kim, T.-Y. Association Between Fusobacterium nucleatum, Pathway Mutation, and Patient Prognosis in Colorectal Cancer. Ann. Surg. Oncol. 2018, 25, 3389–3395. [Google Scholar] [CrossRef]
- Ito, M.; Kanno, S.; Nosho, K.; Sukawa, Y.; Mitsuhashi, K.; Kurihara, H.; Igarashi, H.; Takahashi, T.; Tachibana, M.; Takahashi, H.; et al. Association of Fusobacterium nucleatum with clinical and molecular features in colorectal serrated pathway. Int. J. Cancer 2015, 137, 1258–1268. [Google Scholar] [CrossRef]
- Park, C.H.; Han, D.S.; Oh, Y.-H.; Lee, A.; Lee, Y.; Eun, C.S. Role of Fusobacteria in the serrated pathway of colorectal carcinogenesis. Sci. Rep. 2016, 6, 25271. [Google Scholar] [CrossRef]
- The Integrative HMP (iHMP) Research Network Consortium. The Integrative Human Microbiome Project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, B.M.; Crockett, S.D. The clinical impact of serrated colorectal polyps. Clin. Epidemiol. 2017, 9, 113–125. [Google Scholar] [CrossRef]
- Bond, C.E.; Whitehall, V.L.J. How the BRAF V600E Mutation Defines a Distinct Subgroup of Colorectal Cancer: Molecular and Clinical Implications. Gastroenterol. Res. Pract. 2018, 2018. [Google Scholar] [CrossRef]
- Rozek, L.S.; Herron, C.M.; Greenson, J.K.; Moreno, V.; Capella, G.; Rennert, G.; Gruber, S.B. Smoking, gender, and ethnicity predict somatic BRAF mutations in colorectal cancer. Cancer Epidemiol. Biomark. Prev. 2010, 19, 838–843. [Google Scholar] [CrossRef]
- Yaeger, R.; Cercek, A.; Chou, J.F.; Sylvester, B.E.; Kemeny, N.E.; Hechtman, J.F.; Ladanyi, M.; Rosen, N.; Weiser, M.R.; Capanu, M.; et al. BRAF mutation predicts for poor outcomes after metastasectomy in patients with metastatic colorectal cancer. Cancer 2014, 120, 2316–2324. [Google Scholar] [CrossRef]
- Prasanna, T.; Karapetis, C.S.; Roder, D.; Tie, J.; Padbury, R.; Price, T.; Wong, R.; Shapiro, J.; Nott, L.; Lee, M.; et al. The survival outcome of patients with metastatic colorectal cancer based on the site of metastases and the impact of molecular markers and site of primary cancer on metastatic pattern. Acta Oncol. Stockh. Swed. 2018, 57, 1438–1444. [Google Scholar] [CrossRef] [Green Version]
- Ursem, C.; Atreya, C.E.; Van Loon, K. Emerging treatment options for BRAF-mutant colorectal cancer. Gastrointest. Cancer Targets Ther. 2018, 8, 13–23. [Google Scholar] [CrossRef]
- Brändstedt, J.; Wangefjord, S.; Nodin, B.; Eberhard, J.; Sundström, M.; Manjer, J.; Jirström, K. Associations of anthropometric factors with KRAS and BRAF mutation status of primary colorectal cancer in men and women: A cohort study. PLoS ONE 2014, 9, e98964. [Google Scholar]
- Gil Ferreira, C.; Aran, V.; Zalcberg-Renault, I.; Victorino, A.P.; Salem, J.H.; Bonamino, M.H.; Vieira, F.M.; Zalis, M. KRAS mutations: Variable incidences in a Brazilian cohort of 8234 metastatic colorectal cancer patients. BMC Gastroenterol. 2014, 14. [Google Scholar] [CrossRef]
- Reggiani Bonetti, L.; Barresi, V.; Maiorana, A.; Manfredini, S.; Caprera, C.; Bettelli, S. Clinical Impact and Prognostic Role of KRAS/BRAF/PIK3CA Mutations in Stage I Colorectal Cancer. Dis. Markers 2018, 2018. [Google Scholar] [CrossRef]
- Therkildsen, C.; Bergmann, T.K.; Henrichsen-Schnack, T.; Ladelund, S.; Nilbert, M. The predictive value of KRAS, NRAS, BRAF, PIK3CA and PTEN for anti-EGFR treatment in metastatic colorectal cancer: A systematic review and meta-analysis. Acta Oncol. Stockh. Swed. 2014, 53, 852–864. [Google Scholar] [CrossRef]
- Nash, G.M.; Gimbel, M.; Cohen, A.M.; Zeng, Z.-S.; Ndubuisi, M.I.; Nathanson, D.R.; Ott, J.; Barany, F.; Paty, P.B. KRAS mutation and microsatellite instability: Two genetic markers of early tumor development that influence the prognosis of colorectal cancer. Ann. Surg. Oncol. 2010, 17, 416–424. [Google Scholar] [CrossRef]
- Mohan, H.M.; Ryan, E.; Balasubramanian, I.; Kennelly, R.; Geraghty, R.; Sclafani, F.; Fennelly, D.; McDermott, R.; Ryan, E.J.; O’Donoghue, D.; et al. Microsatellite instability is associated with reduced disease specific survival in stage III colon cancer. Eur. J. Surg. Oncol. 2016, 42, 1680–1686. [Google Scholar] [CrossRef]
- Popat, S.; Hubner, R.; Houlston, R.S. Systematic review of microsatellite instability and colorectal cancer prognosis. J. Clin. Oncol. 2005, 23, 609–618. [Google Scholar] [CrossRef]
- Kim, C.G.; Ahn, J.B.; Jung, M.; Beom, S.H.; Kim, C.; Kim, J.H.; Heo, S.J.; Park, H.S.; Kim, J.H.; Kim, N.K.; et al. Effects of microsatellite instability on recurrence patterns and outcomes in colorectal cancers. Br. J. Cancer 2016, 115, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Inamura, K. Colorectal Cancers: An Update on Their Molecular Pathology. Cancers 2018, 10, 26. [Google Scholar] [CrossRef]
- Venderbosch, S.; Nagtegaal, I.D.; Maughan, T.S.; Smith, C.G.; Cheadle, J.P.; Fisher, D.; Kaplan, R.; Quirke, P.; Seymour, M.T.; Richman, S.D.; et al. Mismatch repair status and BRAF mutation status in metastatic colorectal cancer patients: A pooled analysis of the CAIRO, CAIRO2, COIN, and FOCUS studies. Clin. Cancer Res. 2014, 20, 5322–5330. [Google Scholar] [CrossRef]
- Benson, A.B.; Venook, A.P.; Cederquist, L.; Chan, E.; Chen, Y.-J.; Cooper, H.S.; Deming, D.; Engstrom, P.F.; Enzinger, P.C.; Fichera, A.; et al. Colon Cancer, Version 1.2017, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2017, 15, 370–398. [Google Scholar] [CrossRef]
- Webber, E.M.; Kauffman, T.L.; O’Connor, E.; Goddard, K.A.B. Systematic review of the predictive effect of MSI status in colorectal cancer patients undergoing 5FU-based chemotherapy. BMC Cancer 2015, 15, 156. [Google Scholar] [CrossRef]
- Kang, S.; Na, Y.; Joung, S.Y.; Lee, S.I.; Oh, S.C.; Min, B.W. The significance of microsatellite instability in colorectal cancer after controlling for clinicopathological factors. Medicine (Baltimore) 2018, 97. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, D.; Jin, L.; Wu, G.; Bai, Z.; Wang, J.; Yao, H.; Zhang, Z. Prognostic value of the combination of microsatellite instability and BRAF mutation in colorectal cancer. Cancer Manag. Res. 2018, 10, 3911–3929. [Google Scholar] [CrossRef]
- Seppälä, T.T.; Böhm, J.P.; Friman, M.; Lahtinen, L.; Väyrynen, V.M.J.; Liipo, T.K.E.; Ristimäki, A.P.; Kairaluoma, M.V.J.; Kellokumpu, I.H.; Kuopio, T.H.I.; et al. Combination of microsatellite instability and BRAF mutation status for subtyping colorectal cancer. Br. J. Cancer 2015, 112, 1966–1975. [Google Scholar] [CrossRef]
- Samowitz, W.S.; Sweeney, C.; Herrick, J.; Albertsen, H.; Levin, T.R.; Murtaugh, M.A.; Wolff, R.K.; Slattery, M.L. Poor survival associated with the BRAF V600E mutation in microsatellite-stable colon cancers. Cancer Res. 2005, 65, 6063–6069. [Google Scholar] [CrossRef]
- Diaz, L.A.; Le, D.T. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N. Engl. J. Med. 2015, 373, 1979. [Google Scholar]
- Lee, S.; Cho, N.-Y.; Choi, M.; Yoo, E.J.; Kim, J.-H.; Kang, G.H. Clinicopathological features of CpG island methylator phenotype-positive colorectal cancer and its adverse prognosis in relation to KRAS/BRAF mutation. Pathol. Int. 2008, 58, 104–113. [Google Scholar] [CrossRef]
- Dahlin, A.M.; Palmqvist, R.; Henriksson, M.L.; Jacobsson, M.; Eklöf, V.; Rutegård, J.; Oberg, A.; Van Guelpen, B.R. The role of the CpG island methylator phenotype in colorectal cancer prognosis depends on microsatellite instability screening status. Clin. Cancer Res. 2010, 16, 1845–1855. [Google Scholar] [CrossRef]
- Ward, R.L.; Cheong, K.; Ku, S.-L.; Meagher, A.; O’Connor, T.; Hawkins, N.J. Adverse prognostic effect of methylation in colorectal cancer is reversed by microsatellite instability. J. Clin. Oncol. 2003, 21, 3729–3736. [Google Scholar] [CrossRef]
- Murcia, O.; Juárez, M.; Rodríguez-Soler, M.; Hernández-Illán, E.; Giner-Calabuig, M.; Alustiza, M.; Egoavil, C.; Castillejo, A.; Alenda, C.; Barberá, V.; et al. Colorectal cancer molecular classification using BRAF, KRAS, microsatellite instability and CIMP status: Prognostic implications and response to chemotherapy. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Juo, Y.Y.; Johnston, F.M.; Zhang, D.Y.; Juo, H.H.; Wang, H.; Pappou, E.P.; Yu, T.; Easwaran, H.; Baylin, S.; van Engeland, M.; et al. Prognostic value of CpG island methylator phenotype among colorectal cancer patients: A systematic review and meta-analysis. Ann. Oncol. 2014, 25, 2314–2327. [Google Scholar] [CrossRef]
- Shiovitz, S.; Bertagnolli, M.M.; Renfro, L.A.; Nam, E.; Foster, N.R.; Dzieciatkowski, S.; Luo, Y.; Lao, V.V.; Monnat, R.J.; Emond, M.J.; et al. CpG island methylator phenotype is associated with response to adjuvant irinotecan-based therapy for stage III colon cancer. Gastroenterology 2014, 147, 637–645. [Google Scholar] [CrossRef]
- Vodolazhsky, D.I.; Timoshkina, N.N.; Panina, S.B. LINE-1 hypomethylation in colon cancer. J. Clin. Oncol. 2018, 36, 588. [Google Scholar] [CrossRef]
- Zhou, N.; Gu, Q. Prognostic and clinicopathological value of p16 protein aberrant expression in colorectal cancer. Medicine (Baltimore) 2018, 97. [Google Scholar] [CrossRef]
- Fu, T.; Liu, Y.; Li, K.; Wan, W.; Pappou, E.P.; Iacobuzio-Donahue, C.A.; Kerner, Z.; Baylin, S.B.; Wolfgang, C.L.; Ahuja, N. Tumors with unmethylated MLH1 and the CpG island methylator phenotype are associated with a poor prognosis in stage II colorectal cancer patients. Oncotarget 2016, 7, 86480–86489. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological Classification | Frequency (%) * | Location | Shape | Mucin Type | Size |
|---|---|---|---|---|---|
| Hyperplastic polyp (HP) | 80–90% | Distal | Sessile, Flat | Variable | <5 mm |
| Microvesicular HP (MVHP) | 60% | Distal | Sessile | Microvesicular | <5 mm |
| Goblet cell HP (GCHP) | 30% | Distal | Sessile | Goblet cells | <5 mm |
| Mucin poor HP (MPHP) | 10% | Distal | Sessile | Poor | <5 mm |
| Sessile serrated adenoma/polyp (SSA/P) | 15–20% | Proximal | Sessile/Flat | Microvesicular | >5 mm |
| Traditional serrated polyp (TSA) | 1–6% | Distal | Sessile/Pedunculated | Not present | >5 mm |
| CIMP Phenotype | CIMP-0 | CIMP-L | CIMP-H |
|---|---|---|---|
| Location | Distal colon | Proximal colon | Proximal colon |
| Gender | no gender bias | Male | Female |
| Pathway | Conventional adenoma | Serrated or Conventional | Serrated adenoma |
| Gene mutations | TP53 mutations | KRAS, TP53 mutations | BRAF mutations |
| Epigenetic alterations | no MLH1 methylation | no MLH1 methylation | MLH1 hypermethylation |
| MSI rate | MSS | MSS | MSI |
| CIN association | positive | positive | negative |
| Prognosis | Variable | High | Poor |
| Serrated Lesion | BRAF/KRAS Status | CIMP Rate | Gene Methylation | MSI Rate |
|---|---|---|---|---|
| HP | BRAF mutated | CIMP-H | MLH1 not methylated | MSS |
| MPHP * | controversial | CIMP-H | controversial | controversial |
| GCHP * | KRAS mutated | CIMP-L | MLH1 not methylated | MSS |
| MVHP * | BRAF mutated | CIMP-H | MLH1 not methylated | MSS |
| SSA/P | BRAF mutated | CIMP-H | MLH1 not methylated | MSS |
| SSAD | BRAF mutated | CIMP-H | MLH1 hypermethylated | MSI |
| TSA | KRAS/BRAF mutated or neither | CIMP-L/-H | MLH1 not methylated | MSS |
| TSA HGD | KRAS mutated | CIMP-L | MGMT hypermethylated | MSS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Palma, F.D.E.; D’Argenio, V.; Pol, J.; Kroemer, G.; Maiuri, M.C.; Salvatore, F. The Molecular Hallmarks of the Serrated Pathway in Colorectal Cancer. Cancers 2019, 11, 1017. https://doi.org/10.3390/cancers11071017
De Palma FDE, D’Argenio V, Pol J, Kroemer G, Maiuri MC, Salvatore F. The Molecular Hallmarks of the Serrated Pathway in Colorectal Cancer. Cancers. 2019; 11(7):1017. https://doi.org/10.3390/cancers11071017
Chicago/Turabian StyleDe Palma, Fatima Domenica Elisa, Valeria D’Argenio, Jonathan Pol, Guido Kroemer, Maria Chiara Maiuri, and Francesco Salvatore. 2019. "The Molecular Hallmarks of the Serrated Pathway in Colorectal Cancer" Cancers 11, no. 7: 1017. https://doi.org/10.3390/cancers11071017