Human Colorectal Carcinoma Infiltrating B Lymphocytes Are Active Secretors of the Immunoglobulin Isotypes A, G, and M


Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
2.2. Detailed Analyses of Tumor Tissue Composition and Ig Secretion in CRC Patients
2.3. Analyses of Primary CRC Tumors and CRC Metastasis
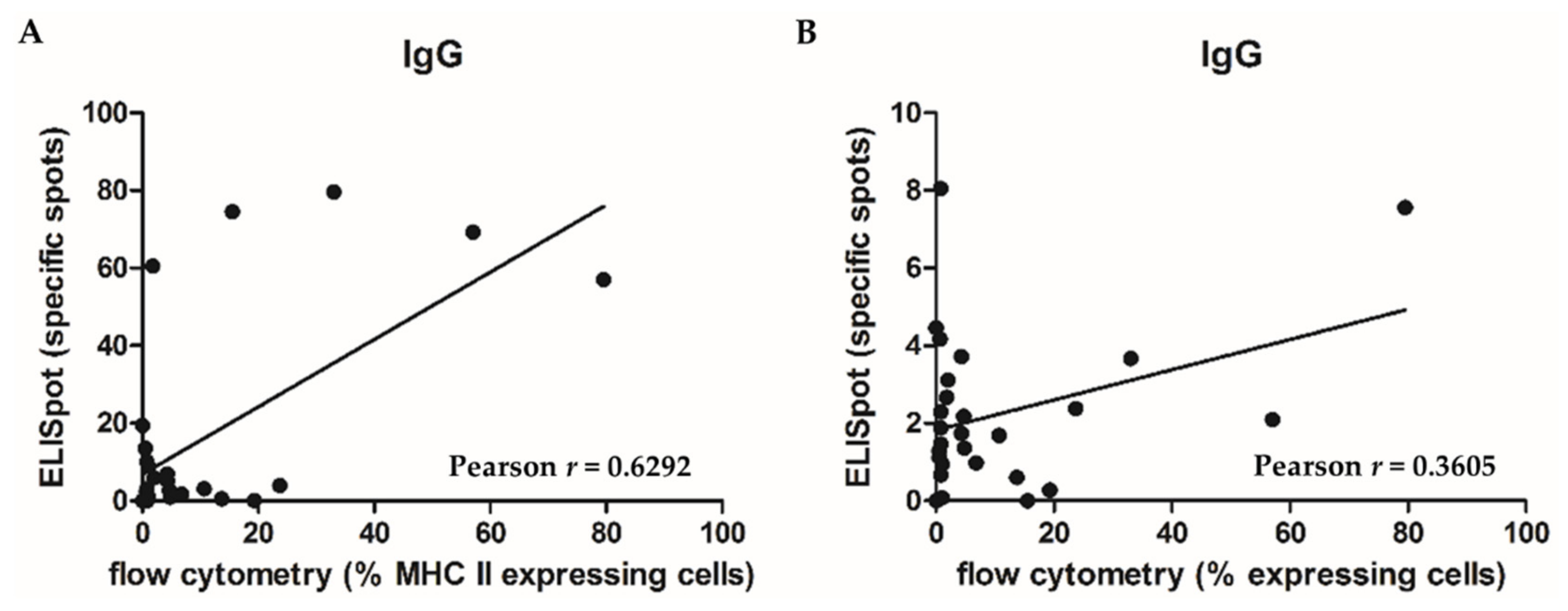
2.4. Correlation of Tumor Tissue Composition (Including the Immune Cell Populations) with Ig Secretion
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Tumor Preparation
4.3. Flow Cytometry
4.4. FluoroSpot
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting:Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Waldner, M.; Obenauf, A.C.; Angell, H.; Fredriksen, T.; Lafontaine, L.; Berger, A.; et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity 2013, 39, 782–795. [Google Scholar] [CrossRef] [PubMed]
- Chin, Y.; Janseens, J.; Vandepitte, J.; Vandenbrande, J.; Opdebeek, L.; Raus, J. Phenotypic analysis of tumor-infiltrating lymphocytes from human breast cancer. Anticancer Res. 1992, 12, 1463–1466. [Google Scholar] [PubMed]
- Nelson, B.H. CD20+ B cells:The other tumor-infiltrating lymphocytes. J. Immunol. 2010, 185, 4977–4982. [Google Scholar] [CrossRef] [PubMed]
- Linnebacher, M. Tumor-infiltrating B cells come into vogue. World J. Gastroenterol. 2013, 19, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, D.R.; Milne, K.; Nelson, B.H. Tumor-Infiltrating Plasma Cells Are Associated with Tertiary Lymphoid Structures, Cytolytic T-Cell Responses, and Superior Prognosis in Ovarian Cancer. Clin. Cancer Res. 2016, 22, 3005–3015. [Google Scholar] [CrossRef] [PubMed]
- Maletzki, C.; Jahnke, A.; Ostwald, C.; Klar, E.; Prall, F.; Linnebacher, M. Ex-vivo clonally expanded B lymphocytes infiltrating colorectal carcinoma are of mature immunophenotype and produce functional IgG. PLoS ONE 2012, 7, e32639. [Google Scholar] [CrossRef] [PubMed]
- Theurich, S.; Schlaak, M.; Steguweit, H.; Heukamp, L.C.; Wennhold, K.; Kurschat, P.; Rabenhorst, A.; Hartmann, K.; Schlösser, H.; Shimabukuro-Vornhagen, A.; et al. Targeting Tumor-Infiltrating B Cells in Cutaneous T-Cell Lymphoma. J. Clin. Oncol. 2016, 34, e110–e116. [Google Scholar] [CrossRef] [PubMed]
- Chiaruttini, G.; Mele, S.; Opzoomer, J.; Crescioli, S.; Ilieva, K.M.; Lacy, K.E.; Karagiannis, S.N. B cells and the humoral response in melanoma:The overlooked players of the tumor microenvironment. Oncoimmunology 2017, 6, e1294296. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.S.; Sahota, R.A.; Milne, K.; Kost, S.E.; Nesslinger, N.J.; Watson, P.H.; Nelson, B.H. CD20+ tumor-infiltrating lymphocytes have an atypical CD27- memory phenotype and together with CD8+ T cells promote favorable prognosis in ovarian cancer. Clin. Cancer Res. 2012, 18, 3281–3292. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro-Vornhagen, A.; Schlößer, H.A.; Gryschok, L.; Malcher, J.; Wennhold, K.; Garcia-Marquez, M.; Herbold, T.; Neuhaus, L.S.; Becker, H.J.; Fiedler, A.; et al. Characterization of tumor-associated B-cell subsets in patients with colorectal cancer. Oncotarget 2014, 5, 4651–4664. [Google Scholar] [CrossRef] [PubMed]
- Spear, S.; Candido, J.B.; McDermott, J.R.; Ghirelli, C.; Maniati, E.; Beers, S.A.; Balkwill, F.R.; Kocher, H.M.; Capasso, M. Discrepancies in the Tumor Microenvironment of Spontaneous and Orthotopic Murine Models of Pancreatic Cancer Uncover a New Immunostimulatory Phenotype for B Cells. Front. Immunol. 2019, 10, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagès, F.; Galon, J.; Dieu-Nosjean, M.C.; Tartour, E.; Sautès-Fridman, C.; Fridman, W.H. Immune infiltration in human tumors:A prognostic factor that should not be ignored. Oncogene 2010, 29, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Nzula, S.; Going, J.J.; Stott, D.I. Antigen-driven clonal proliferation, somatic hypermutation, and selection of B lymphocytes infiltrating human ductal breast carcinomas. Cancer Res. 2003, 63, 3275–3280. [Google Scholar] [PubMed]
- Brandtzaeg, P.; Farstad, I.N.; Johansen, F.E.; Morton, H.C.; Norderhaug, I.N.; Yamanaka, T. The B-cell system of human mucosae and exocrine glands. Immunol. Rev. 1999, 171, 45–87. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, F.; Mullins, C.S.; Krohn, M.; Harnack, C.; Ramer, R.; Krämer, O.H.; Klar, E.; Huehns, M.; Linnebacher, M. Establishment and characterization of HROC69—A Crohn’s related colonic carcinoma cell line and its matched patient-derived xenograft. Sci. Rep. 2016, 6, 24671. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age | Tumor Site | TNM Stage | Tumor Grade | UICC Stage | Follow Up | |
|---|---|---|---|---|---|---|---|---|
| Outcome | Years | |||||||
| HROC225 | F | 76 | Rectum | T4 N2 M1 L0 V0 | G3 | UICC IV | dead | 4.0 |
| HROC228 | M | 54 | Sigmoid | T3 N0 M0 L0 V0 | G2 | UICC IIa | tumor free | 2.9 |
| HROC245 | M | 73 | Rectum | T3 N0 M0 L0 V0 | G3 | UICC IIa | tumor free | 4.2 |
| HROC247 | M | 74 | right flexure | T3 N0 M0 L0 V0 | G2 | UICC IIa | dead | 4.5 |
| HROC248 | M | 65 | Coecum | T4 N2 M1 L0 V0 | G2 | UICC IV | dead | 2.3 |
| HROC251 | M | 85 | Ascending | T3 N0 M0 L0 V0 | G1 | UICC IIa | dead | 2.2 |
| HROC252 | M | 45 | Descending | T4 N0 M0 L0 V1 | G3 | UICC IIb | tumor free | 3.6 |
| Sigmoid | T4 N0 M0 L0 V1 | G3 | UICC IIb | |||||
| Rectum | T4 N0 M0 L0 V1 | G3 | UICC IIb | |||||
| HROC256 | M | 70 | Descending | T3 N0 M0 L0 V0 | G2 | UICC IIa | tumor free | 4.3 |
| HROC258 | F | 77 | Recto-sigmoid | T3 N0 M0 L0 V0 | G2 | UICC IIa | dead | 2.6 |
| HROC274 | M | 64 | Sigmoid | T3 N1 M1 L0 V1 | G2 | UICC IV | progression free | 4.0 |
| HROC277 | M | 77 | Coecum | T4 N0 M1 L0 V1 | G2 | UICC IV | dead | 3.3 |
| Liver metastasis | ||||||||
| HROC278 | F | 76 | Ascending | T4 N2 M1 L1 V1 | G3 | UICC IV | dead | 2.1 |
| Located peritoneal metastasis | ||||||||
| HROC283 | F | 48 | Sigmoid | T3 N1 M0 L1 V0 | G2 | UICC IIIa | tumor free | 3.4 |
| HROC285 | F | 30 | Descending | T4 N2 M1 L1 V0 | G2 | UICC IV | tumor free | 3.7 |
| HROC296 | F | 92 | Ascending | T3 N0 M0 L0 V0 | G2 | UICC IIa | tumor free | 3.6 |
| HROC309 | M | 86 | Descending | T3 N0 M0 L0 V1 | G2 | UICC IIa | dead | 1.2 |
| HROC310 | M | 76 | Ascending | T3 N0 M0 L0 V0 | G2 | UICC IIa | tumor free | 3.4 |
| HROC314 | F | 76 | Sigmoid | T4 N2 M1 L0 V0 | G2 | UICC IV | dead | 2.9 |
| HROC315 | F | 42 | Left flexure | T3 N2 M0 L0 V1 | G3 | UICC IIIb | tumor free | 2.6 |
| HROC319 | M | 67 | Coecum | T4 N2 M0 L1 V0 | G3 | UICC IIIb | tumor free | 2.5 |
| HROC325 | F | 80 | Sigmoid | T3 N0 M0 L0 V0 | G2 | UICC IIa | tumor free | 3.1 |
| HROC230 | M | 74 | Liver metastasis colon cancer | UICC IV | dead | 6.2 | ||
| HROC317 | M | 72 | Liver metastasis colon cancer | UICC IV | dead | 6.5 | ||
| HROC253 | M | 61 | Liver metastasis rectal cancer | UICC IV | dead | 4.6 | ||
| HROC111 | M | 78 | Brain metastasis colon cancer | UICC IV | dead | 3.1 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mullins, C.S.; Gock, M.; Krohn, M.; Linnebacher, M. Human Colorectal Carcinoma Infiltrating B Lymphocytes Are Active Secretors of the Immunoglobulin Isotypes A, G, and M. Cancers 2019, 11, 776. https://doi.org/10.3390/cancers11060776
Mullins CS, Gock M, Krohn M, Linnebacher M. Human Colorectal Carcinoma Infiltrating B Lymphocytes Are Active Secretors of the Immunoglobulin Isotypes A, G, and M. Cancers. 2019; 11(6):776. https://doi.org/10.3390/cancers11060776
Chicago/Turabian StyleMullins, Christina Susanne, Michael Gock, Mathias Krohn, and Michael Linnebacher. 2019. "Human Colorectal Carcinoma Infiltrating B Lymphocytes Are Active Secretors of the Immunoglobulin Isotypes A, G, and M" Cancers 11, no. 6: 776. https://doi.org/10.3390/cancers11060776