Allergic Signs in Glioma Pathology: Current Knowledge and Future Perspectives

Abstract
:1. Introduction
2. Epidemiology of Gliomas and GBM in Allergic Subjects
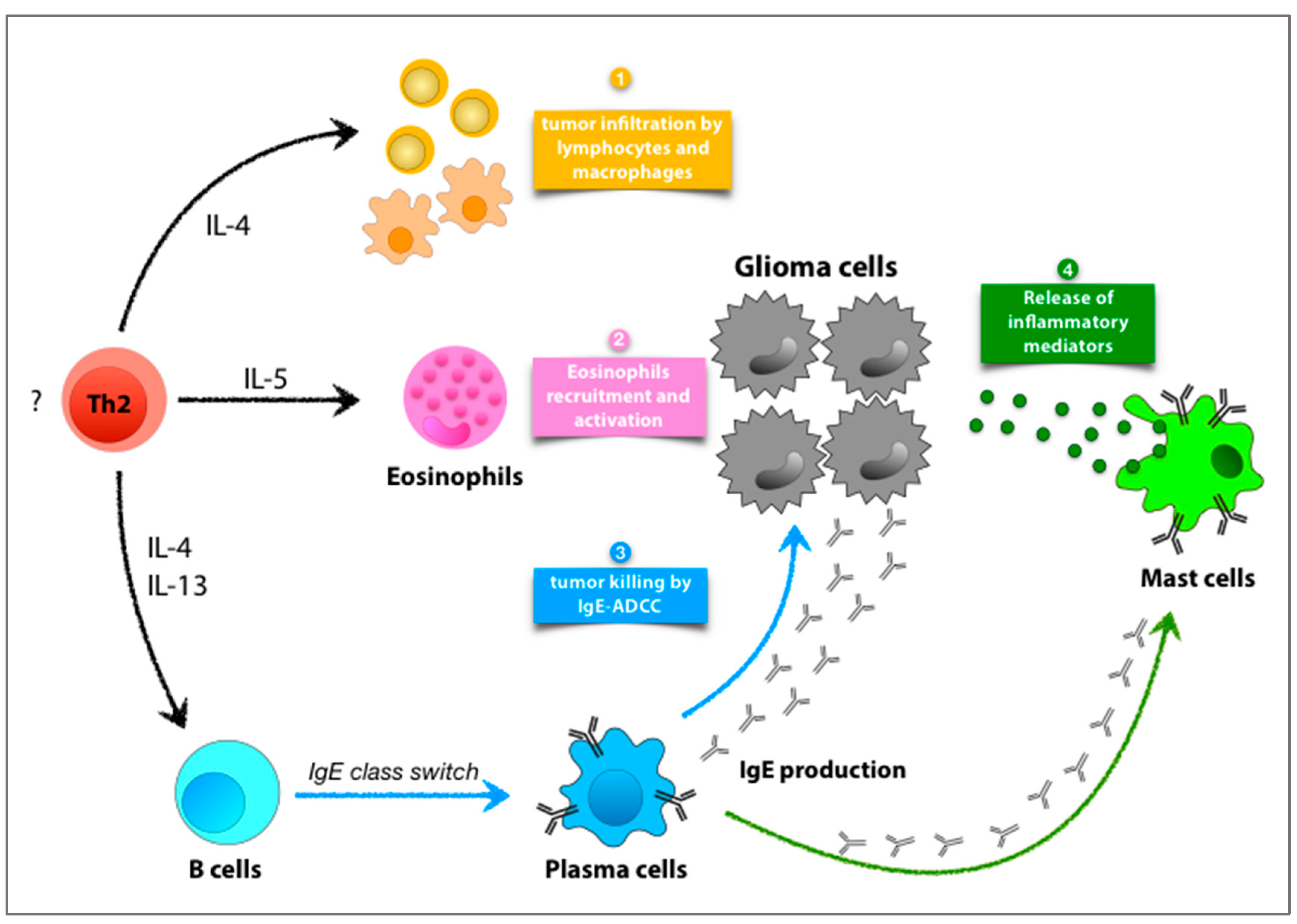
3. Allergic Mediators in Glioma and GBM
4. Conclusions and Future Perspectives
Author Contributions
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Wen, P.Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.E.; Kahng, J.Y.; Kim, S.H.; Park, J.S.; Yoon, S.J.; Um, J.-Y.; Kim, W.K.; Lee, J.-K.; Park, J.; et al. Human glioblastoma arises from subventricular zone cells with low-level driver mutations. Nature 2018, 560, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Orzan, F.; De Bacco, F.; Crisafulli, G.; Pellegatta, S.; Mussolin, B.; Siravegna, G.; D’Ambrosio, A.; Comoglio, P.M.; Finocchiaro, G.; Boccaccio, C. Genetic Evolution of Glioblastoma Stem-Like Cells From Primary to Recurrent Tumor. Stem Cells 2017, 35, 2218–2228. [Google Scholar] [CrossRef] [PubMed]
- Palm, N.W.; Rosenstein, R.K.; Medzhitov, R. Allergic host defences. Nature 2012, 484, 465–472. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Disis, M.L. Immune regulation of cancer. J. Clin. Oncol. 2010, 28, 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- DeNardo, D.G.; Barreto, J.B.; Andreu, P.; Vasquez, L.; Tawfik, D.; Kolhatkar, N.; Coussens, L.M. CD4(+) T cells regulate pulmonary metastasis of mammary carcinomas by enhancing protumor properties of macrophages. Cancer Cell 2009, 16, 91–102. [Google Scholar] [CrossRef]
- Crawford, G.; Hayes, M.D.; Seoane, R.C.; Ward, S.; Dalessandri, T.; Lai, C.; Healy, E.; Kipling, D.; Proby, C.; Moyes, C.; et al. Epithelial damage and tissue γδ T cells promote a unique tumor-protective IgE response. Nat. Immunol. 2018, 19, 859–870. [Google Scholar] [CrossRef]
- Rigoni, A.; Colombo, M.P.; Pucillo, C. Mast cells, basophils and eosinophils: From allergy to cancer. Semin. Immunol. 2018, 35, 29–34. [Google Scholar] [CrossRef]
- Hollande, C.; Boussier, J.; Ziai, J.; Nozawa, T.; Bondet, V.; Phung, W.; Lu, B.; Duffy, D.; Paradis, V.; Mallet, V.; et al. Inhibition of the dipeptidyl peptidase DPP4 (CD26) reveals IL-33-dependent eosinophil-mediated control of tumor growth. Nat. Immunol. 2019, 20, 257–264. [Google Scholar] [CrossRef]
- Munitz, A.; Hogan, S.P. Alarming eosinophils to combat tumors. Nat. Immunol. 2019, 20, 250–252. [Google Scholar] [CrossRef]
- Schlehofer, B.; Blettner, M.; Becker, N.; Martinsohn, C.; Wahrendorf, J. Medical risk factors and the development of brain tumors. Cancer 1992, 69, 2541–2547. [Google Scholar] [CrossRef] [Green Version]
- Schlehofer, B.; Blettner, M.; Preston-Martin, S.; Niehoff, D.; Wahrendorf, J.; Arslan, A.; Ahlbom, A.; Choi, W.N.; Giles, G.G.; Howe, G.R.; et al. Role of medical history in brain tumour development. Results from the international adult brain tumour study. Int. J. Cancer 1999, 82, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Wiemels, J.L.; Wiencke, J.K.; Sison, J.D.; Miike, R.; McMillan, A.; Wrensch, M. History of allergies among adults with glioma and controls. Int. J. Cancer 2002, 98, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Wiemels, J.L.; Wiencke, J.K.; Patoka, J.; Moghadassi, M.; Chew, T.; McMillan, A.; Miike, R.; Barger, G.; Wrensch, M. Reduced immunoglobulin E and allergy among adults with glioma compared with controls. Cancer Res. 2004, 64, 8468–8473. [Google Scholar] [CrossRef]
- Brenner, A.V.; Linet, M.S.; Fine, H.A.; Shapiro, W.R.; Selker, R.G.; Black, P.M.; Inskip, P.D. History of allergies and autoimmune diseases and risk of brain tumors in adults. Int. J. Cancer 2002, 99, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Schwartzbaum, J.; Jonsson, F.; Ahlbom, A.; Preston-Martin, S.; Lönn, S.; Söderberg, K.C.; Feychting, M. Cohort studies of association between self-reported allergic conditions, immune-related diagnoses and glioma and meningioma risk. Int. J. Cancer 2003, 106, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Schoemaker, M.J.; Swerdlow, A.J.; Hepworth, S.J.; McKinney, P.A.; van Tongeren, M.; Muir, K.R. History of allergies and risk of glioma in adults. Int. J. Cancer 2006, 119, 2165–2172. [Google Scholar] [CrossRef] [Green Version]
- Linos, E.; Raine, T.; Alonso, A.; Michaud, D. Atopy and risk of brain tumors: A meta-analysis. J. Natl. Cancer Inst. 2007, 99, 1544–1550. [Google Scholar] [CrossRef]
- Wigertz, A.; Lönn, S.; Schwartzbaum, J.; Hall, P.; Auvinen, A.; Christensen, H.C.; Johansen, C.; Klaeboe, L.; Salminen, T.; Schoemaker, M.J.; et al. Allergic conditions and brain tumor risk. Am. J. Epidemiol. 2007, 166, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Berg-Beckhoff, G.; Schüz, J.; Blettner, M.; Münster, E.; Schlaefer, K.; Wahrendorf, J.; Schlehofer, B. History of allergic disease and epilepsy and risk of glioma and meningioma (INTERPHONE study group, Germany). Eur. J. Epidemiol. 2009, 24, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Il’yasova, D.; McCarthy, B.; Marcello, J.; Schildkraut, J.M.; Moorman, P.G.; Krishnamachari, B.; Ali-Osman, F.; Bigner, D.D.; Davis, F. Association between glioma and history of allergies, asthma and eczema: A case-control study with three groups of controls. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Lachance, D.H.; Yang, P.; Johnson, D.R.; Decker, P.A.; Kollmeyer, T.M.; McCoy, L.S.; Rice, T.; Xiao, Y.; Ali-Osman, F.; Wang, F.; et al. Associations of high-grade glioma with glioma risk alleles and histories of allergy and smoking. Am. J. Epidemiol. 2011, 174, 574–581. [Google Scholar] [CrossRef]
- Cahoon, E.K.; Inskip, P.D.; Gridley, G.; Brenner, A.V. Immune-related conditions and subsequent risk of brain cancer in a cohort of 4.5 million male US veterans. Br. J. Cancer 2014, 110, 1825–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouchieu, C.; Raherison, C.; Piel, C.; Migault, L.; Carles, C.; Fabbro-Perray, P.; Loiseau, H.; Guillamo, J.-S.; Lebailly, P.; Baldi, I. Allergic conditions and risk of glioma and meningioma in the CERENAT case-control study. J. Neurooncol. 2018, 138, 271–281. [Google Scholar] [CrossRef]
- Schwartzbaum, J.; Ahlbom, A.; Malmer, B.; Lönn, S.; Brookes, A.J.; Doss, H.; Debinski, W.; Henriksson, R.; Feychting, M. Polymorphisms associated with asthma are inversely related to glioblastoma multiforme. Cancer Res. 2005, 65, 6459–6465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartzbaum, J.; Seweryn, M.; Holloman, C.; Harris, R.; Handelman, S.K.; Rempala, G.A.; Huang, R.-P.; Burkholder, B.; Brandemihl, A.; Kallberg, H.; et al. Association between Prediagnostic Allergy-Related Serum Cytokines and Glioma. PLoS ONE 2015, 10, e0137503. [Google Scholar] [CrossRef] [PubMed]
- Wiemels, J.L.; Wilson, D.; Patil, C.; Patoka, J.; McCoy, L.; Rice, T.; Schwartzbaum, J.; Heimberger, A.; Sampson, J.H.; Chang, S.; et al. IgE, allergy and risk of glioma: Update from the San Francisco Bay Area Adult Glioma Study in the temozolomide era. Int. J. Cancer 2009, 125, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Calboli, F.C.F.; Cox, D.G.; Buring, J.E.; Gaziano, J.M.; Ma, J.; Stampfer, M.; Willett, W.C.; Tworoger, S.S.; Hunter, D.J.; Camargo, C.A.; et al. Prediagnostic plasma IgE levels and risk of adult glioma in four prospective cohort studies. J. Natl. Cancer Inst. 2011, 103, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Schlehofer, B.; Siegmund, B.; Linseisen, J.; Schüz, J.; Rohrmann, S.; Becker, S.; Michaud, D.; Melin, B.; Bas Bueno-de-Mesquita, H.; Peeters, P.H.M.; et al. Primary brain tumours and specific serum immunoglobulin E: A case-control study nested in the European Prospective Investigation into Cancer and Nutrition cohort. Allergy 2011, 66, 1434–1441. [Google Scholar] [CrossRef]
- Schwartzbaum, J.; Ding, B.; Johannesen, T.B.; Osnes, L.T.N.; Karavodin, L.; Ahlbom, A.; Feychting, M.; Grimsrud, T.K. Association between prediagnostic IgE levels and risk of glioma. J. Natl. Cancer Inst. 2012, 104, 1251–1259. [Google Scholar] [CrossRef]
- Yu, J.S.; Wei, M.X.; Chiocca, E.A.; Martuza, R.L.; Tepper, R.I. Treatment of glioma by engineered interleukin 4-secreting cells. Cancer Res. 1993, 53, 3125–3128. [Google Scholar]
- Hayes, R.L.; Koslow, M.; Hiesiger, E.M.; Hymes, K.B.; Hochster, H.S.; Moore, E.J.; Pierz, D.M.; Chen, D.K.; Budzilovich, G.N.; Ransohoff, J. Improved long term survival after intracavitary interleukin-2 and lymphokine-activated killer cells for adults with recurrent malignant glioma. Cancer 1995, 76, 840–852. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, S.; Dimeco, F.; Pollo, B.; Cirenei, N.; Colombo, B.M.; Bruzzone, M.G.; Cattaneo, E.; Vescovi, A.; Didonato, S.; Colombo, M.P.; et al. Limited efficacy of the HSV-TK/GCV system for gene therapy of malignant gliomas and perspectives for the combined transduction of the interleukin-4 gene. Hum. Gene Ther. 1997, 8, 1345–1353. [Google Scholar] [CrossRef]
- Andreansky, S.; He, B.; van Cott, J.; McGhee, J.; Markert, J.M.; Gillespie, G.Y.; Roizman, B.; Whitley, R.J. Treatment of intracranial gliomas in immunocompetent mice using herpes simplex viruses that express murine interleukins. Gene Ther. 1998, 5, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedetti, S.; Bruzzone, M.G.; Pollo, B.; DiMeco, F.; Magrassi, L.; Pirola, B.; Cirenei, N.; Colombo, M.P.; Finocchiaro, G. Eradication of rat malignant gliomas by retroviral-mediated, in vivo delivery of the interleukin 4 gene. Cancer Res. 1999, 59, 645–652. [Google Scholar] [PubMed]
- Benedetti, S.; Pirola, B.; Pollo, B.; Magrassi, L.; Bruzzone, M.G.; Rigamonti, D.; Galli, R.; Selleri, S.; Di Meco, F.; De Fraja, C.; et al. Gene therapy of experimental brain tumors using neural progenitor cells. Nat. Med. 2000, 6, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Kalesnikoff, J.; Huber, M.; Lam, V.; Damen, J.E.; Zhang, J.; Siraganian, R.P.; Krystal, G. Monomeric IgE stimulates signaling pathways in mast cells that lead to cytokine production and cell survival. Immunity 2001, 14, 801–811. [Google Scholar] [CrossRef]
- Sayed, B.A.; Christy, A.; Quirion, M.R.; Brown, M.A. The master switch: The role of mast cells in autoimmunity and tolerance. Annu. Rev. Immunol. 2008, 26, 705–739. [Google Scholar] [CrossRef]
- Põlajeva, J.; Sjösten, A.M.; Lager, N.; Kastemar, M.; Waern, I.; Alafuzoff, I.; Smits, A.; Westermark, B.; Pejler, G.; Uhrbom, L.; et al. Mast cell accumulation in glioblastoma with a potential role for stem cell factor and chemokine CXCL12. PLoS ONE 2011, 6, e25222. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Hui, A.-M.; Su, Q.; Vortmeyer, A.; Kotliarov, Y.; Pastorino, S.; Passaniti, A.; Menon, J.; Walling, J.; Bailey, R.; et al. Neuronal and glioma-derived stem cell factor induces angiogenesis within the brain. Cancer Cell 2006, 9, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Põlajeva, J.; Bergström, T.; Edqvist, P.-H.; Lundequist, A.; Sjösten, A.; Nilsson, G.; Smits, A.; Bergqvist, M.; Pontén, F.; Westermark, B.; et al. Glioma-derived macrophage migration inhibitory factor (MIF) promotes mast cell recruitment in a STAT5-dependent manner. Mol. Oncol. 2014, 8, 50–58. [Google Scholar] [CrossRef]
- Roy, A.; Coum, A.; Marinescu, V.D.; Põlajeva, J.; Smits, A.; Nelander, S.; Uhrbom, L.; Westermark, B.; Forsberg-Nilsson, K.; Pontén, F.; et al. Glioma-derived plasminogen activator inhibitor-1 (PAI-1) regulates the recruitment of LRP1 positive mast cells. Oncotarget 2015, 6, 23647–23661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehrer, S.; Rheinstein, P.H.; Rosenzweig, K.E. Allergy may confer better survival on patients with gliomas. Clin. Neurol. Neurosurg. 2018, 177, 63–67. [Google Scholar] [CrossRef]
- Tepper, R.I.; Pattengale, P.K.; Leder, P. Murine interleukin-4 displays potent anti-tumor activity in vivo. Cell 1989, 57, 503–512. [Google Scholar] [CrossRef]
- Karagiannis, S.N.; Wang, Q.; East, N.; Burke, F.; Riffard, S.; Bracher, M.G.; Thompson, R.G.; Durham, S.R.; Schwartz, L.B.; Balkwill, F.R.; et al. Activity of human monocytes in IgE antibody-dependent surveillance and killing of ovarian tumor cells. Eur. J. Immunol. 2003, 33, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Ehtesham, M.; Samoto, K.; Kabos, P.; Acosta, F.L.; Gutierrez, M.A.R.; Black, K.L.; Yu, J.S. Treatment of intracranial glioma with in situ interferon-gamma and tumor necrosis factor-alpha gene transfer. Cancer Gene Ther. 2002, 9, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef]
- Lu, L.-F.; Lind, E.F.; Gondek, D.C.; Bennett, K.A.; Gleeson, M.W.; Pino-Lagos, K.; Scott, Z.A.; Coyle, A.J.; Reed, J.L.; Van Snick, J.; et al. Mast cells are essential intermediaries in regulatory T-cell tolerance. Nature 2006, 442, 997–1002. [Google Scholar] [CrossRef]
- de Vries, V.C.; Wasiuk, A.; Bennett, K.A.; Benson, M.J.; Elgueta, R.; Waldschmidt, T.J.; Noelle, R.J. Mast cell degranulation breaks peripheral tolerance. Am. J. Transplant. 2009, 9, 2270–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cildir, G.; Pant, H.; Lopez, A.F.; Tergaonkar, V. The transcriptional program, functional heterogeneity and clinical targeting of mast cells. J. Exp. Med. 2017, 214, 2491–2506. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.; Kohanbash, G.; Ohkuri, T.; Kosaka, A.; Chen, X.; Ikeura, M.; Wang, T.C.; Okada, H. Histamine deficiency promotes accumulation of immunosuppressive immature myeloid cells and growth of murine gliomas. Oncoimmunology 2015, 4, e1047581. [Google Scholar] [CrossRef] [Green Version]
- Ohtsu, H.; Tanaka, S.; Terui, T.; Hori, Y.; Makabe-Kobayashi, Y.; Pejler, G.; Tchougounova, E.; Hellman, L.; Gertsenstein, M.; Hirasawa, N.; et al. Mice lacking histidine decarboxylase exhibit abnormal mast cells. FEBS Lett. 2001, 502, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, S.; Reilly, K.M. The role of the immune system in neurofibromatosis type 1-associated nervous system tumors. CNS Oncol. 2017, 6, 45–60. [Google Scholar] [CrossRef]


{kind=link}
| Study Type | Number of Glioma Cases | Number of Control Subjects | Association with Allergy (95% CI) | Year [Ref.] |
|---|---|---|---|---|
| Case-control | 115 | 418 | RR = 0.71 (0.5–1.0) | 1992 [13] |
| Case-control | 1178 | 2493 | RR = 0.59 (0.49–0.71) | 1999 [14] |
| Case-control | 405 | 402 | OR = 0.5 (0.3–0.7) | 2002 [15] |
| Case-control | 489 | 799 | OR = 0.67 (0.52–0.86) | 2002 [17] |
| Cohort study | Cohort I (14535 subjects/33 glioma cases) | HR = 0.38 (0.05–3.13) 1 HR = 0.46 (0.18–1.21) 2 | 2003 [18] | |
| Cohort II (29573 subjects/42 glioma cases) | HR = 2.60 (0.86–7.81) 1 HR = 0.45 (0.11–1.92) 2 | |||
| Case-control | 965 | 1716 | OR = 0.63 (0.53–0.76) | 2006 [19] |
| Meta-Analysis | Participants (53223 subjects/3450 glioma cases) | RR = 0.61 (0.55–0.67) | 2007 [20] | |
| Case-control | 1527 | 3309 | OR = 0.70 (0.61–0.80) | 2007 [21] |
| Case-control | 366 | 1494 | OR = 0.92 (0.70–1.22) | 2009 [22] |
| Case-control | 388 | 80 (siblings) | OR = 0.53 (0.15–1.84) | 2009 [23] |
| 191 (friends) | OR = 0.54 (0.28–1.07) | |||
| 177 (clinic-based controls) | OR = 0.34 (0.23–0.50) | |||
| Case-control | 855 | 1160 | OR = 0.62 (0.51–0.76) | 2011 [24] |
| Cohort study | 4.5 million subjects/4383 malignant neoplasm brain 3 | Rate ratio = 0.60 (0.43–0.83) | 2014 [25] | |
| Case-control | 273 | 982 | OR = 0.52 (0.36–0.75) | 2018 [26] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanza, M.; Finocchiaro, G. Allergic Signs in Glioma Pathology: Current Knowledge and Future Perspectives. Cancers 2019, 11, 404. https://doi.org/10.3390/cancers11030404
Costanza M, Finocchiaro G. Allergic Signs in Glioma Pathology: Current Knowledge and Future Perspectives. Cancers. 2019; 11(3):404. https://doi.org/10.3390/cancers11030404
Chicago/Turabian StyleCostanza, Massimo, and Gaetano Finocchiaro. 2019. "Allergic Signs in Glioma Pathology: Current Knowledge and Future Perspectives" Cancers 11, no. 3: 404. https://doi.org/10.3390/cancers11030404