GSTO1*CC Genotype (rs4925) Predicts Shorter Survival in Clear Cell Renal Cell Carcinoma Male Patients

 ,
,  and
and
Abstract
:1. Introduction
2. Results
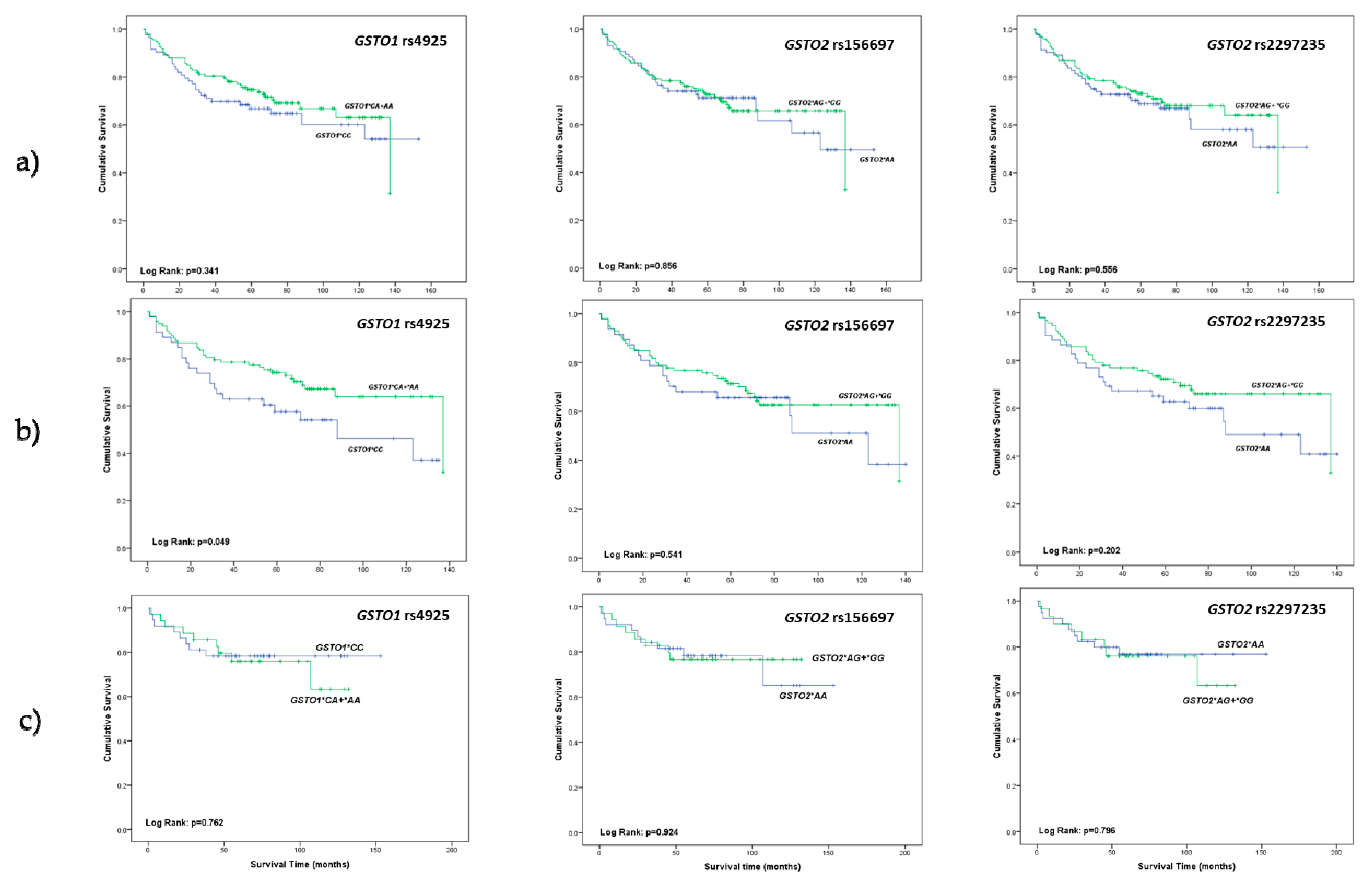
2.1. The Relevance of GSTO1 and GSTO2 Polymorphisms in Overall Survival of ccRCC Patients
2.2. Predicting Effect of GSTO1 and GSTO2 Polymorphisms on Overall Mortality in ccRCC Patients
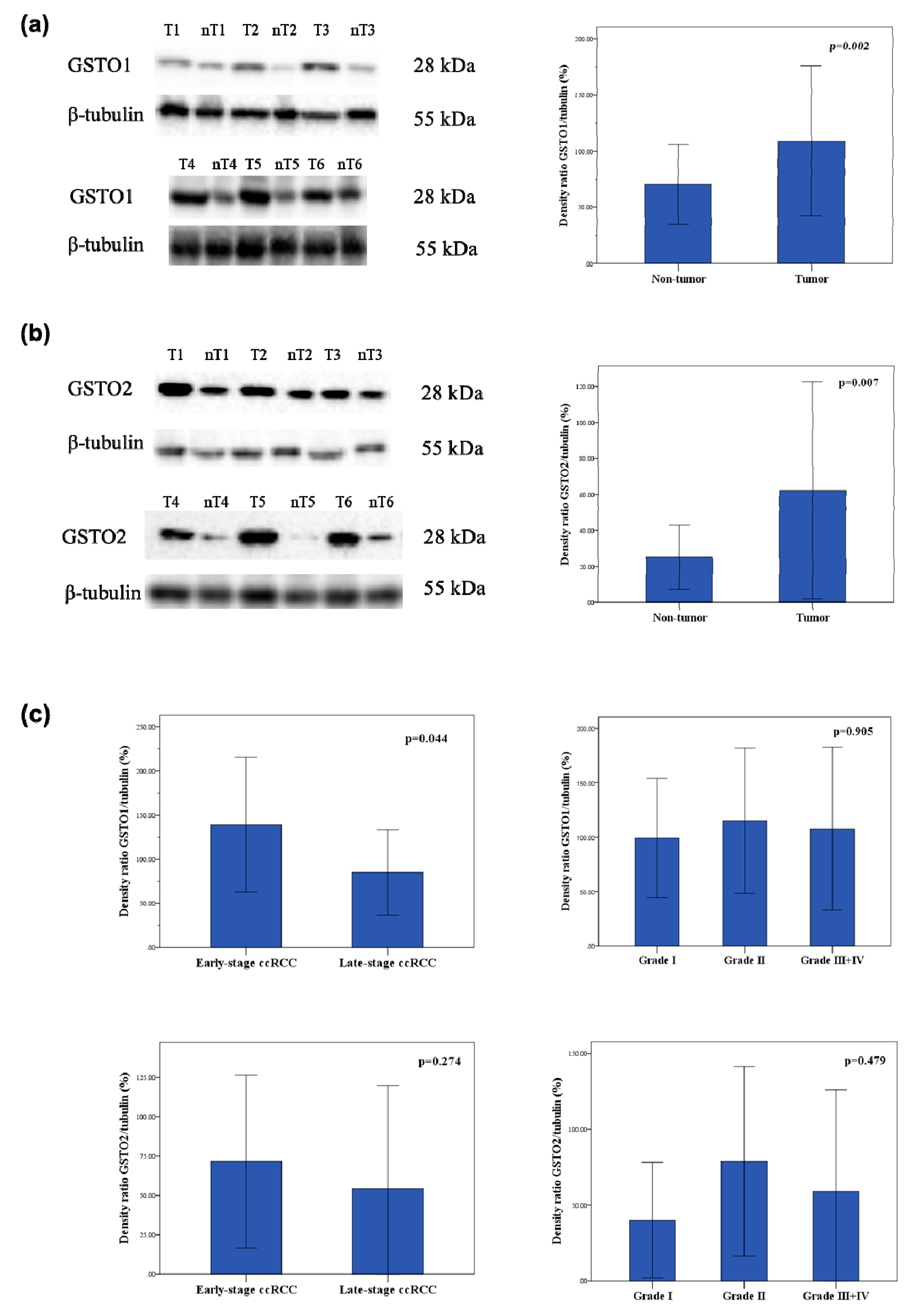
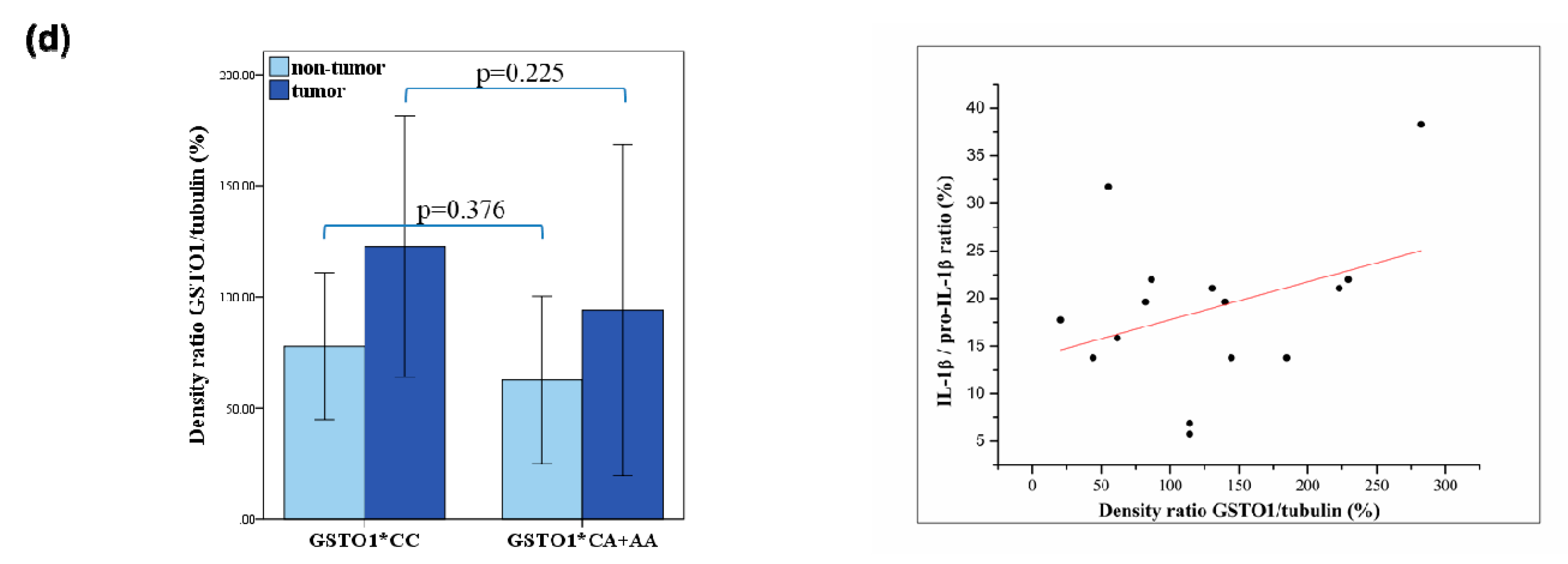
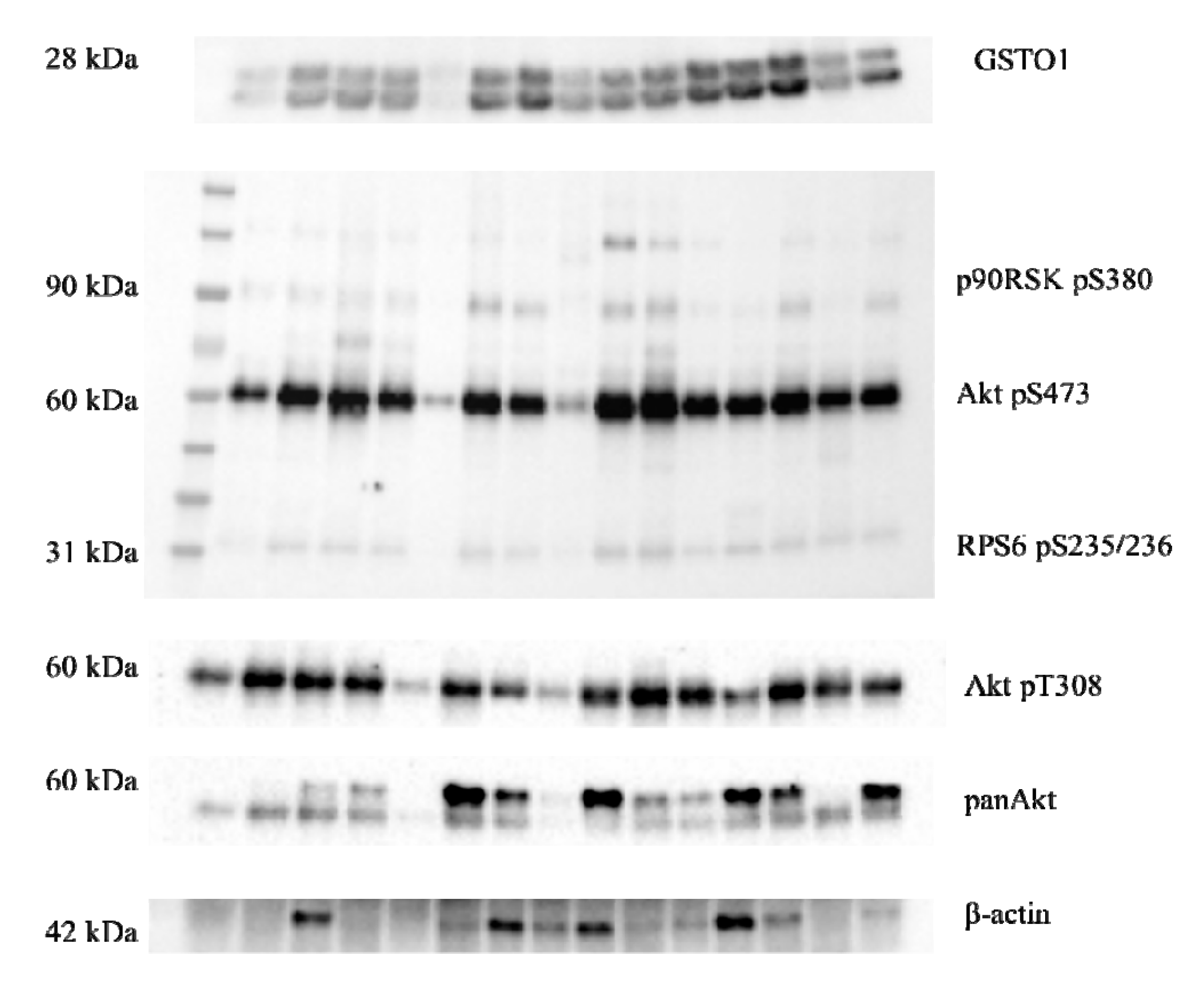
2.3. Expression of Glutathione Transferase Omega Class Enzymes and Downstream Effectors of PI3K/Akt and Raf/MEK/ERK Signaling Pathway in ccRCC
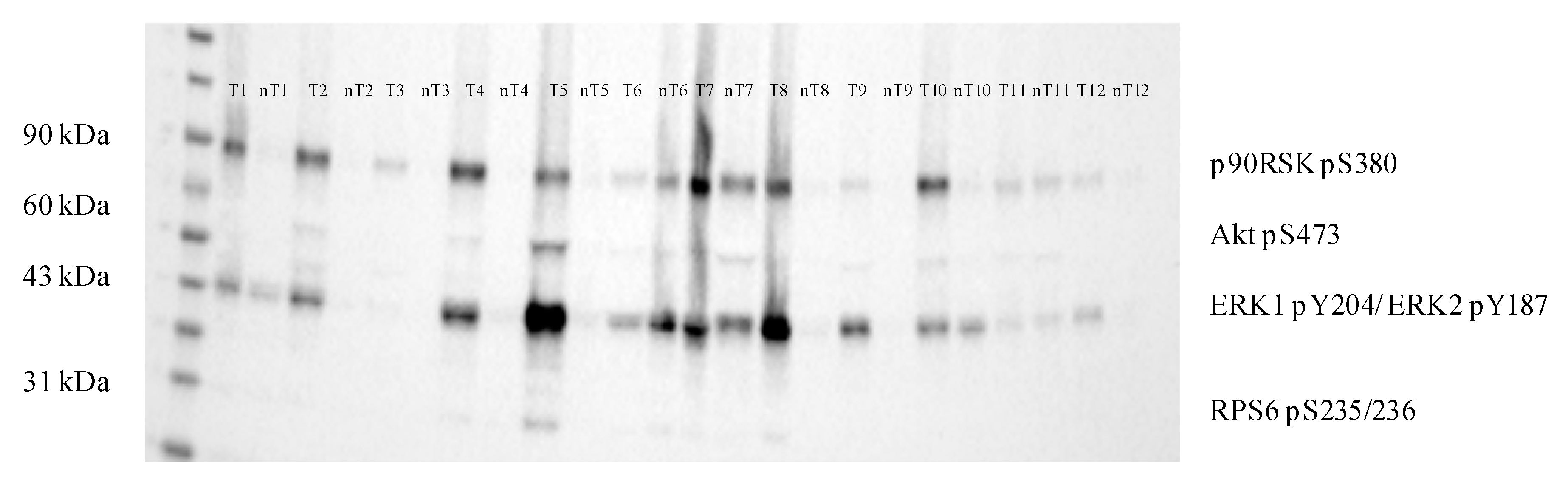
2.4. Immunoprecipitation of GSTO1 and Associated Proteins in Tumor ccRCC Tissue
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Sample Preparation
4.3. Genotyping
4.4. Western Blot and Immunoprecipitation
4.5. Determination of IL-1β and pro- IL-1β Levels
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature 2013, 499, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, J.J.; Manley, B.J.; Khan, N.; Gao, J.; Carlo, M.I.; Cheng, E.H. Overcome tumor heterogeneity-imposed therapeutic barriers through convergent genomic biomarker discovery: A braided cancer river model of kidney cancer. Semin. Cell Dev. Biol. 2017, 64, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehdi, A.; Riazalhosseini, Y. Epigenome Aberrations: Emerging Driving Factors of the Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2017, 18, 1774. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, D.J.; Simon, M.C. Genetic and metabolic hallmarks of clear cell renal cell carcinoma. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 23–31. [Google Scholar] [CrossRef]
- Kumar, A.; Kumari, N.; Gupta, V.; Prasad, R. Renal Cell Carcinoma: Molecular Aspects. Indian J. Clin. Biochem. 2018, 33, 246–254. [Google Scholar] [CrossRef]
- Guo, H.; German, P.; Bai, S.; Barnes, S.; Guo, W.; Qi, X.; Lou, H.; Liang, J.; Jonasch, E.; Mills, G.B.; et al. The PI3K/AKT Pathway and Renal Cell Carcinoma. J. Genet. Genomics 2015, 42, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Jann, J.; Xavier, C.; Wu, H. Glutaredoxin 1 (Grx1) Protects Human Retinal Pigment Epithelial Cells from Oxidative Damage by Preventing AKT Glutathionylation. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2821. [Google Scholar] [CrossRef]
- Tew, K.D.; Townsend, D.M. Glutathione-s-transferases as determinants of cell survival and death. Antioxid. Redox Signal. 2012, 17, 1728–1737. [Google Scholar] [CrossRef] [Green Version]
- Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Matic, M.; Coric, V.; Djukic, T.; Radic, T.; Simic, T. Glutathione Transferases: Potential Targets to Overcome Chemoresistance in Solid Tumors. Int. J. Mol. Sci. 2018, 19, 3785. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Dong, D. Human cytosolic glutathione transferases: Structure, function, and drug discovery. Trends Pharmacol. Sci. 2012, 33, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Abid, A.; Ajaz, S.; Khan, A.R.; Zehra, F.; Hasan, A.S.; Sultan, G.; Mohsin, R.; Hashmi, A.; Niamatullah, N.; Rizvi, S.A.-H.; et al. Analysis of the glutathione S-transferase genes polymorphisms in the risk and prognosis of renal cell carcinomas. Case-control and meta-analysis. Urol. Oncol. Semin. Orig. Investig. 2016, 34, 419.e1–419.e12. [Google Scholar] [CrossRef] [PubMed]
- Board, P.G.; Menon, D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3267–3288. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Shi, H.; Hou, Q.; Mo, Z.; Xie, X. GSTM1 and GSTT1 polymorphisms contribute to renal cell carcinoma risk: Evidence from an updated meta-analysis. Sci. Rep. 2015, 5, 17971. [Google Scholar] [CrossRef]
- De Martino, M.; Klatte, T.; Schatzl, G.; Remzi, M.; Waldert, M.; Haitel, A.; Stancik, I.; Kramer, G.; Marberger, M. Renal cell carcinoma Fuhrman grade and histological subtype correlate with complete polymorphic deletion of glutathione S-transferase M1 gene. J. Urol. 2010, 183, 878–883. [Google Scholar] [CrossRef]
- Coric, V.M.; Simic, T.P.; Pekmezovic, T.D.; Basta-Jovanovic, G.M.; Savic-Radojevic, A.R.; Radojevic-Skodric, S.M.; Matic, M.G.; Suvakov, S.R.; Dragicevic, D.P.; Radic, T.M.; et al. GSTM1 genotype is an independent prognostic factor in clear cell renal cell carcinoma. Urol. Oncol. 2017, 35, 409–417. [Google Scholar] [CrossRef]
- Whitbread, A.K.; Masoumi, A.; Tetlow, N.; Schmuck, E.; Coggan, M.; Board, P.G. Characterization of the Omega Class of Glutathione Transferases. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2005; Volume 401, pp. 78–99. ISBN 978-0-12-182806-6. [Google Scholar]
- Board, P.G.; Menon, D. Structure, function and disease relevance of Omega-class glutathione transferases. Arch. Toxicol. 2016, 90, 1049–1067. [Google Scholar] [CrossRef]
- Dulhunty, A.; Gage, P.; Curtis, S.; Chelvanayagam, G.; Board, P. The Glutathione Transferase Structural Family Includes a Nuclear Chloride Channel and a Ryanodine Receptor Calcium Release Channel Modulator. J. Biol. Chem. 2001, 276, 3319–3323. [Google Scholar] [CrossRef] [Green Version]
- Laliberte, R.E.; Perregaux, D.G.; Hoth, L.R.; Rosner, P.J.; Jordan, C.K.; Peese, K.M.; Eggler, J.F.; Dombroski, M.A.; Geoghegan, K.F.; Gabel, C.A. Glutathione S-Transferase Omega 1-1 Is a Target of Cytokine Release Inhibitory Drugs and May Be Responsible for Their Effect on Interleukin-1β Posttranslational Processing. J. Biol. Chem. 2003, 278, 16567–16578. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Brock, J.; Liu, D.; Board, P.G.; Oakley, A.J. Structural Insights into the Dehydroascorbate Reductase Activity of Human Omega-Class Glutathione Transferases. J. Mol. Biol. 2012, 420, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Menon, D.; Board, P.G. A Role for Glutathione Transferase Omega 1 (GSTO1-1) in the Glutathionylation Cycle. J. Biol. Chem. 2013, 288, 25769–25779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piaggi, S.; Raggi, C.; Corti, A.; Pitzalis, E.; Mascherpa, M.C.; Saviozzi, M.; Pompella, A.; Casini, A.F. Glutathione transferase omega 1-1 (GSTO1-1) plays an anti-apoptotic role in cell resistance to cisplatin toxicity. Carcinogenesis 2010, 31, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Pan, L.; Yuan, Y.; Lang, J.; Mao, N. Identification of Platinum-Resistance Associated Proteins through Proteomic Analysis of Human Ovarian Cancer Cells and Their Platinum-Resistant Sublines. J. Proteome Res. 2007, 6, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Chen, I.; Shimoda, L.A.; Park, Y.; Zhang, C.; Tran, L.; Zhang, H.; Semenza, G.L. Chemotherapy-Induced Ca2+ Release Stimulates Breast Cancer Stem Cell Enrichment. Cell Rep. 2017, 18, 1946–1957. [Google Scholar] [CrossRef]
- Allen, M.; Zou, F.; Chai, H.S.; Younkin, C.S.; Miles, R.; Nair, A.A.; Crook, J.E.; Pankratz, V.S.; Carrasquillo, M.M.; Rowley, C.N.; et al. Glutathione S-transferase omega genes in Alzheimer and Parkinson disease risk, age-at-diagnosis and brain gene expression: An association study with mechanistic implications. Mol. Neurodegener. 2012, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, B.; Salavaggione, O.E.; Pelleymounter, L.L.; Moon, I.; Eckloff, B.W.; Schaid, D.J.; Wieben, E.D.; Weinshilboum, R.M. Glutathione S-transferase omega 1 and omega 2 pharmacogenomics. Drug Metab. Dispos. 2006, 34, 1237–1246. [Google Scholar] [CrossRef]
- Djukic, T.I.; Savic-Radojevic, A.R.; Pekmezovic, T.D.; Matic, M.G.; Pljesa-Ercegovac, M.S.; Coric, V.M.; Radic, T.M.; Suvakov, S.R.; Krivic, B.N.; Dragicevic, D.P.; et al. Glutathione S-transferase T1, O1 and O2 polymorphisms are associated with survival in muscle invasive bladder cancer patients. PLoS ONE 2013, 8, e74724. [Google Scholar] [CrossRef] [Green Version]
- Radic, T.M.; Coric, V.M.; Pljesa-Ercegovac, M.S.; Basta-Jovanovic, G.M.; Radojevic-Skodric, S.M.; Dragicevic, D.P.; Matic, M.G.; Bogdanovic, L.M.; Dzamic, Z.M.; Simic, T.P.; et al. Concomitance of Polymorphisms in Glutathione Transferase Omega Genes Is Associated with Risk of Clear Cell Renal Cell Carcinoma. Tohoku J. Exp. Med. 2018, 246, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.Y.; Jung, H.J.; Moon, A. Molecular Markers in Sex Differences in Cancer. Toxicol. Res. 2019, 35, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Coll, R.C.; O’Neill, L.A.J. The Cytokine Release Inhibitory Drug CRID3 Targets ASC Oligomerisation in the NLRP3 and AIM2 Inflammasomes. PLoS ONE 2011, 6, e29539. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Ikemoto, S.; Narita, K.; Sugimura, K.; Wada, S.; Yasumoto, R.; Kishimoto, T.; Nakatani, T. Interleukin-6, tumour necrosis factor α and interleukin-1β in patients with renal cell carcinoma. Br. J. Cancer 2002, 86, 1396–1400. [Google Scholar] [CrossRef] [PubMed]
- Petrella, B.L.; Vincenti, M.P. Interleukin-1β mediates metalloproteinase-dependent renal cell carcinoma tumor cell invasion through the activation of CCAAT enhancer binding protein β. Cancer Med. 2012, 1, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Dorak, M.T.; Karpuzoglu, E. Gender Differences in Cancer Susceptibility: An Inadequately Addressed Issue. Front. Genet. 2012, 3, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djukic, T.; Simic, T.; Pljesa-Ercegovac, M.; Matic, M.; Suvakov, S.; Coric, V.; Dragicevic, D.; Savic-Radojevic, A. Upregulated glutathione transferase omega-1 correlates with progression of urinary bladder carcinoma. Redox Rep. 2017, 22, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Urzúa, U.; Roby, K.F.; Gangi, L.M.; Cherry, J.M.; Powell, J.I.; Munroe, D.J. Transcriptomic analysis of an in vitro murine model of ovarian carcinoma: Functional similarity to the human disease and identification of prospective tumoral markers and targets. J. Cell. Physiol. 2006, 206, 594–602. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.; Peng, B.; Shao, Q.; Qian, W.; Zhang, J.-Y. Identification of glutathione S-transferase omega 1 (GSTO1) protein as a novel tumor-associated antigen and its autoantibody in human esophageal squamous cell carcinoma. Tumor Biol. 2014, 35, 10871–10877. [Google Scholar] [CrossRef]
- Piaggi, S.; Marchi, S.; Ciancia, E.; DeBortoli, N.; Lazzarotti, A.; Saviozzi, M.; Raggi, C.; Fierabracci, V.; Visvikis, A.; Bisgaard, H.C.; et al. Nuclear translocation of glutathione transferase omega is a progression marker in Barrett’s esophagus. Oncol. Rep. 2009, 21, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, S.; Fuoco, I.; Di Fluri, G.; Costa, F.; Ricchiuti, A.; Biondi, G.; Nardini, V.; Scarpato, R. Genomic instability and cellular stress in organ biopsies and peripheral blood lymphocytes from patients with colorectal cancer and predisposing pathologies. Oncotarget 2015, 6, 14852–14864. [Google Scholar] [CrossRef] [Green Version]
- Lusini, L.; Tripodi, S.A.; Rossi, R.; Giannerini, F.; Giustarini, D.; Del Vecchio, M.T.; Barbanti, G.; Cintorino, M.; Tosi, P.; Di Simplicio, P. Altered glutathione anti-oxidant metabolism during tumor progression in human renal-cell carcinoma. Int. J. Cancer 2001, 91, 55–59. [Google Scholar] [CrossRef]
- Rodic, S.; Vincent, M.D. Reactive oxygen species (ROS) are a key determinant of cancer’s metabolic phenotype: ROS are a key determinant of cancer’s metabolic phenotype. Int. J. Cancer 2018, 142, 440–448. [Google Scholar] [CrossRef]
- Pljesa-Ercegovac, M.; Mimic-Oka, J.; Dragicevic, D.; Savic-Radojevic, A.; Opacic, M.; Pljesa, S.; Radosavljevic, R.; Simic, T. Altered antioxidant capacity in human renal cell carcinoma: Role of glutathione associated enzymes. Urol. Oncol. 2008, 26, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ye, Z.; Singh, S.; Townsend, D.M.; Tew, K.D. An evolving understanding of the S-glutathionylation cycle in pathways of redox regulation. Free Radic. Biol. Med. 2018, 120, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Scirè, A.; Cianfruglia, L.; Minnelli, C.; Bartolini, D.; Torquato, P.; Principato, G.; Galli, F.; Armeni, T. Glutathione compartmentalization and its role in glutathionylation and other regulatory processes of cellular pathways. BioFactors 2019, 45, 152–168. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Wu, S.; Tong, H.; He, W.; Gou, X. HOXA6 inhibits cell proliferation and induces apoptosis by suppressing the PI3K/Akt signaling pathway in clear cell renal cell carcinoma. Int. J. Oncol. 2019, 54, 2095–2105. [Google Scholar] [CrossRef]
- Hemmings, B.A.; Restuccia, D.F. PI3K-PKB/Akt Pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011189. [Google Scholar] [CrossRef] [Green Version]
- Takata, T.; Tsuchiya, Y.; Watanabe, Y. 90-kDa ribosomal S6 kinase 1 is inhibited by S-glutathionylation of its active-site cysteine residue during oxidative stress. FEBS Lett. 2013, 587, 1681–1686. [Google Scholar] [CrossRef] [Green Version]
- Teppo, H.-R.; Soini, Y.; Karihtala, P. Reactive Oxygen Species-Mediated Mechanisms of Action of Targeted Cancer Therapy. Oxid. Med. Cell. Longev. 2017, 2017, 1485283. [Google Scholar] [CrossRef]
- Chiou, J.-F.; Tai, C.-J.; Wang, Y.-H.; Liu, T.-Z.; Jen, Y.-M.; Shiau, C.-Y. Sorafenib induces preferential apoptotic killing of a drug- and radio-resistant hep G2 cells through a mitochondria-dependent oxidative stress mechanism. Cancer Biol. Ther. 2009, 8, 1904–1913. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Zhang, Z.-J.; Hu, S.-Q.; Mak, S.-H.; Xu, D.-P.; Choi, C.-L.; Wang, Y.-Q.; Tsim, W.-K.; Lee, M.-Y.; Rong, J.-H.; et al. Sunitinib Produces Neuroprotective Effect Via Inhibiting Nitric Oxide Overproduction. CNS Neurosci. Ther. 2014, 20, 244–252. [Google Scholar] [CrossRef]
- Tsuboi, K.; Bachovchin, D.A.; Speers, A.E.; Spicer, T.P.; Fernandez-Vega, V.; Hodder, P.; Rosen, H.; Cravatt, B.F. Potent and Selective Inhibitors of Glutathione S-transferase Omega 1 that Impair Cancer Drug Resistance. J. Am. Chem. Soc. 2011, 133, 16605–16616. [Google Scholar] [CrossRef] [Green Version]
- Ramkumar, K.; Samanta, S.; Kyani, A.; Yang, S.; Tamura, S.; Ziemke, E.; Stuckey, J.A.; Li, S.; Chinnaswamy, K.; Otake, H.; et al. Mechanistic evaluation and transcriptional signature of a glutathione S-transferase omega 1 inhibitor. Nat. Commun. 2016, 7, 13084. [Google Scholar] [CrossRef] [PubMed]
- Srigley, J.R.; Delahunt, B.; Eble, J.N.; Egevad, L.; Epstein, J.I.; Grignon, D.; Hes, O.; Moch, H.; Montironi, R.; Tickoo, S.K.; et al. The International Society of Urological Pathology (ISUP) Vancouver Classification of Renal Neoplasia. Am. J. Surg. Pathol. 2013, 37, 1469–1489. [Google Scholar] [CrossRef] [PubMed]
- Sobin, L.H.; Gospodarowicz, M.K.; Wittekind, C.; International Union against Cancer (Eds.) TNM Classification of Malignant Tumours, 7th ed.; Wiley-Blackwell: Chichester, UK; Hoboken, NJ, USA, 2010; ISBN 978-1-4443-3241-4. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Patients, n (%) | Male Patients, n (%) |
|---|---|---|
| Age, years (mean ± SD) | 59.19 ± 11.58 | 57.79 ± 11.22 |
| Gender, n (%) | ||
| Female | 74 (32) | |
| Male | 154 (68) | |
| Fuhrman nuclear grade, n (%) | ||
| G1 | 28 (14) | 17 (12) |
| G2 | 111 (56) | 76 (57) |
| G3 | 50 (26) | 36 (27) |
| G4 | 8 (4) | 5 (4) |
| pT stage, n (%) | ||
| pT1 | 97 (45) | 61 (40) |
| pT2 | 24 (11) | 14 (10) |
| pT3 | 90 (42) | 69 (47) |
| pT4 | 5 (2) | 4 (3) |
| Variable | Category | Events, n (%) | HR (95% CI) b | p |
|---|---|---|---|---|
| GSTO1 rs4925 | ||||
| FNR a | G1/G2/G3/G4 | 2 (7)/32 (29)/29 (58)/5 (63) | 1.57 (1.08–2.27) | 0.017 |
| pT stage | pT1/pT2/pT3/pT4 | 14 (14)/8 (33)/ 51 (57)/3 (60) | 2.01 (1.46–2.76) | <0.001 |
| GSTO1 rs4925 | ||||
| *CC | 32 (38) | 1.53 (0.91–2.58) | 0.107 | |
| *CA + *AA | 47 (34) | 1.00 | ||
| GSTO2 rs156697 | ||||
| FNR | G1/G2/G3/G4 | 2 (7)/32 (29)/29 (58)/5 (63) | 1.58 (1.09–2.27) | 0.015 |
| pT stage | pT1/pT2/pT3/pT4 | 14 (14)/8 (33)/51 (57)/3 (60) | 1.97 (1.43–2.70) | <0.001 |
| GSTO2 rs156697 | ||||
| *AA | 31 (35) | 1.11 (0.66–1.88) | 0.689 | |
| *AG + *GG | 48 (34) | 1.00 | ||
| GSTO2 rs2297235 | ||||
| FNR | G1/G2/G3/G4 | 2 (7)/32 (29)/29 (58)/5 (63) | 1.57 (1.08–2.27) | 0.016 |
| pT stage | pT1/pT2/pT3/pT4 | 14 (14)/8 (33)/51 (57)/3 (60) | 1.96 (1.43–2.69) | <0.001 |
| GSTO2 rs2297235 | ||||
| *AA | 34 (36) | 1.24 (0.74–2.07) | 0.425 | |
| *AG + *GG | 44 (34) | 1.00 | ||
| Variable | Category | Events, n (%) | HR (95% CI) b | p |
|---|---|---|---|---|
| GSTO1 rs4925 | ||||
| FNR a | G1/G2/G3/G4 | 2 (12)/23 (30)/26 (72)/2 (40) | 1.58 (1.03–2.43) | 0.037 |
| pT stage | pT1/pT2/pT3/pT4 | 10(16)/5 (36)/42 (61)/2 (50) | 1.83 (1.28–2.62) | 0.001 |
| GSTO1 rs4925 | ||||
| *CC | 23 (49) | 1.89 (1.04–3.42) | 0.037 | |
| *CA + *AA | 38 (36) | 1.00 | ||
| GSTO2 rs156697 | ||||
| FNR | G1/G2/G3/G4 | 2 (12)/23 (30)/26 (72)/2 (40) | 1.56 (1.02–2.38) | 0.040 |
| pT stage | pT1/pT2/pT3/pT4 | 10(16)/5 (36)/42 (61)/2 (50) | 1.83 (1.29–2.60) | 0.001 |
| GSTO2 rs156697 | ||||
| *AA | 21 (43) | 1.32 (0.71–2.43) | 0.380 | |
| *AG + *GG | 40 (38) | 1.00 | ||
| GSTO2 rs2297235 | ||||
| FNR | G1/G2/G3/G4 | 2 (12)/23 (30)/26 (72)/2 (40) | 1.59 (1.04–2.46) | 0.034 |
| pT stage | pT1/pT2/pT3/pT4 | 10(16)/5 (36)/42 (61)/2 (50) | 1.81 (1.27–2.59) | 0.001 |
| GSTO2 rs2297235 | ||||
| *AA | 24 (45) | 1.60 (0.88–2.92) | 0.127 | |
| *AG + *GG | 36 (37) | 1.00 | ||
| Variable | Category | Events, n (%) | HR (95% CI) b | p |
|---|---|---|---|---|
| GSTO1 rs4925 | ||||
| FNR a | G1/G2/G3/G4 | 0 (0)/9 (26)/3 (21)/3 (100) | 1.92 (0.87–4.26) | 0.107 |
| pT stage | pT1/pT2/pT3/pT4 | 4(11)/3 (30)/9 (43)/1 (100) | 2.20 (1.08–4.46) | 0.029 |
| GSTO1 rs4925 | ||||
| *CC | 9 (24) | 1.01 (0.34–3.00) | 0.992 | |
| *CA + *AA | 9 (26) | 1.00 | ||
| GSTO2 rs156697 | ||||
| FNR | G1/G2/G3/G4 | 0 (0)/9 (26)/3 (21)/3 (100) | 1.95 (0.89–4.27) | 0.098 |
| pT stage | pT1/pT2/pT3/pT4 | 4(11)/3 (30)/9 (43)/1 (100) | 2.23 (1.11–4.47) | 0.024 |
| GSTO2 rs156697 | ||||
| *AA | 10 (26) | 0.78 (0.27–2.28) | 0.654 | |
| *AG+*GG | 8 (23) | 1.00 | ||
| GSTO2 rs2297235 | ||||
| FNR | G1/G2/G3/G4 | 0 (0)/9 (26)/3 (21)/3 (100) | 1.99 (0.90–4.38) | 0.089 |
| pT stage | pT1/pT2/pT3/pT4 | 4(11)/3 (30)/9 (43)/1 (100) | 2.11 (1.05–4.24) | 0.035 |
| GSTO2 rs2297235 | ||||
| *AA | 10 (24) | 0.77 (0.26–2.31) | 0.645 | |
| *AG+*GG | 8 (27) | 1.00 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radic, T.; Coric, V.; Bukumiric, Z.; Pljesa-Ercegovac, M.; Djukic, T.; Avramovic, N.; Matic, M.; Mihailovic, S.; Dragicevic, D.; Dzamic, Z.; et al. GSTO1*CC Genotype (rs4925) Predicts Shorter Survival in Clear Cell Renal Cell Carcinoma Male Patients. Cancers 2019, 11, 2038. https://doi.org/10.3390/cancers11122038
Radic T, Coric V, Bukumiric Z, Pljesa-Ercegovac M, Djukic T, Avramovic N, Matic M, Mihailovic S, Dragicevic D, Dzamic Z, et al. GSTO1*CC Genotype (rs4925) Predicts Shorter Survival in Clear Cell Renal Cell Carcinoma Male Patients. Cancers. 2019; 11(12):2038. https://doi.org/10.3390/cancers11122038
Chicago/Turabian StyleRadic, Tanja, Vesna Coric, Zoran Bukumiric, Marija Pljesa-Ercegovac, Tatjana Djukic, Natasa Avramovic, Marija Matic, Smiljana Mihailovic, Dejan Dragicevic, Zoran Dzamic, and et al. 2019. "GSTO1*CC Genotype (rs4925) Predicts Shorter Survival in Clear Cell Renal Cell Carcinoma Male Patients" Cancers 11, no. 12: 2038. https://doi.org/10.3390/cancers11122038