

A PDMS–Agar Hybrid Microfluidic Device for the Investigation of Chemical–Mechanical Associative Learning Behavior of C. elegans
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nematode Strains and Culture
2.3. Chip Design and Fabrication
2.4. Chemical–Mechanical Associative Learning Behavior Assay
3. Results and Discussion
3.1. Construction of Chemical Gradient
3.2. Associative Learning Behavior in Chip
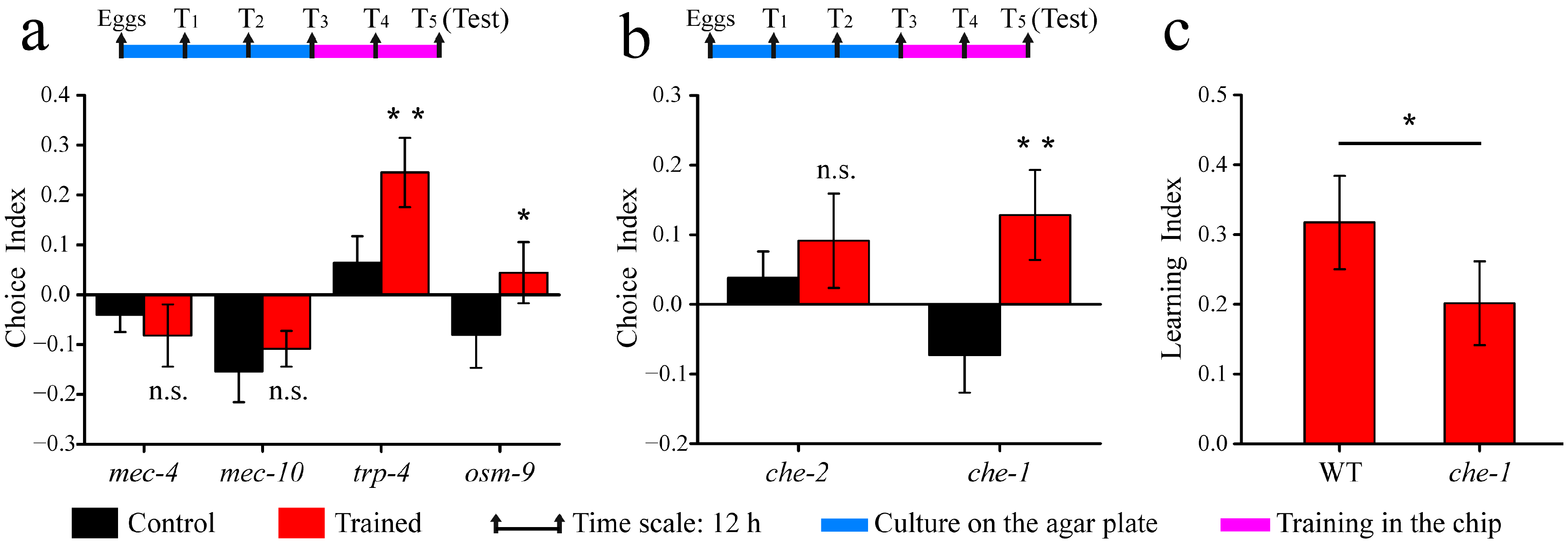
3.3. Associative Learning Behavior of Mechanosensory and Chemosensory Mutants
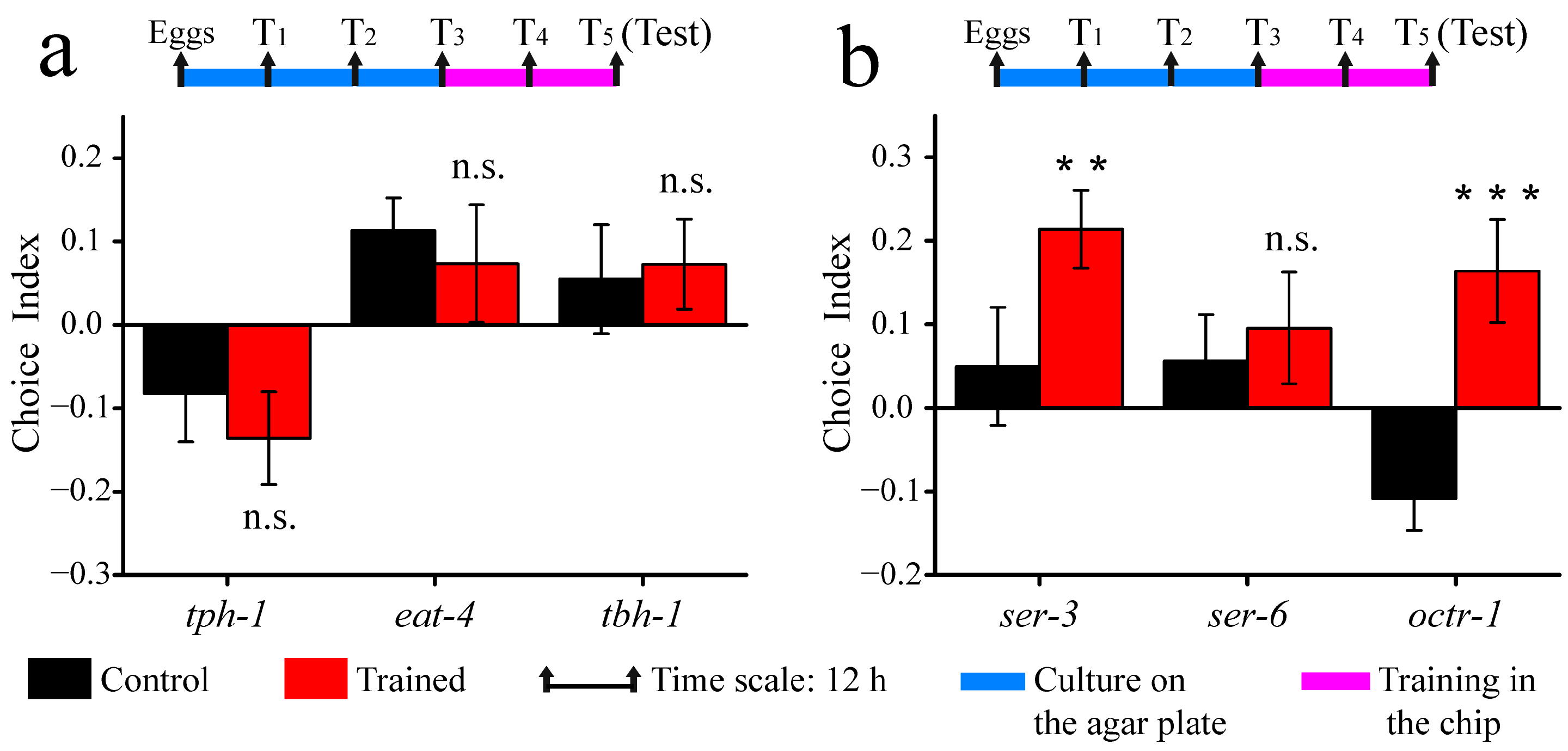
3.4. Associative Learning Behavior of Neurotransmitter and Receptor Mutants
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahmani, A.; Chew, Y.L. Investigating the molecular mechanisms of learning and memory using Caenorhabditis elegans. J. Neurochem. 2021, 159, 417–451. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Burgos, D. Associative learning and high-level cognitive processes in the control of food-related behaviors. Curr. Opin. Behav. Sci. 2022, 47, 101207. [Google Scholar] [CrossRef]
- Mitchell, C.J.; De Houwer, J.; Lovibond, P.F. The propositional nature of human associative learning. Behav. Brain Sci. 2009, 32, 183–198. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, M.L.; Tao, H.; Geng, C.; Guo, F.; Hu, B.; Wang, R.; Hou, X.Y. ErbB4 in parvalbumin-positive interneurons mediates proactive interference in olfactory associative reversal learning. Neuropsychopharmacology 2022, 47, 1292–1303. [Google Scholar] [CrossRef]
- Taylor, J.A.; Hasegawa, M.; Benoit, C.M.; Freire, J.A.; Theodore, M.; Ganea, D.A.; Innocenti, S.M.; Lu, T.; Gründemann, J. Single cell plasticity and population coding stability in auditory thalamus upon associative learning. Nat. Commun. 2021, 12, 2438. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Deng, J.; Zhang, Z.; Zhang, Z.-Y.; Sun, Y.-G.; Yang, T.; Yao, H. Orbitofrontal control of visual cortex gain promotes visual associative learning. Nat. Commun. 2020, 11, 2784. [Google Scholar] [CrossRef] [PubMed]
- Arieli, E.; Younis, N.; Moran, A. Distinct Progressions of Neuronal Activity Changes Underlie the Formation and Consolidation of a Gustatory Associative Memory. J. Neurosci. 2021, 42, 909–921. [Google Scholar] [CrossRef]
- Nishio, N.; Mohri-Shiomi, A.; Nishida, Y.; Hiramatsu, N.; Kodama-Namba, E.; Kimura, K.D.; Kuhara, A.; Mori, I. A novel and conserved protein AHO-3 is required for thermotactic plasticity associated with feeding states in Caenorhabditis elegans. Genes Cells 2012, 17, 365–386. [Google Scholar] [CrossRef] [Green Version]
- D’Aquin, S.; Szonyi, A.; Mahn, M.; Krabbe, S.; Grundemann, J.; Luthi, A. Compartmentalized dendritic plasticity during associative learning. Science 2022, 376, eabf7052. [Google Scholar] [CrossRef]
- Chiang, Y.-C.; Liao, C.-P.; Pan, C.-L. A serotonergic circuit regulates aversive associative learning under mitochondrial stress in C. elegans. Proc. Natl. Acad. Sci. USA 2022, 119, e2115533119. [Google Scholar] [CrossRef]
- Adel, M.; Griffith, L.C. The Role of Dopamine in Associative Learning in Drosophila: An Updated Unified Model. Neurosci. Bull. 2021, 37, 831–852. [Google Scholar] [CrossRef] [PubMed]
- Baracchi, D.; Cabirol, A.; Devaud, J.-M.; Haase, A.; D’ettorre, P.; Giurfa, M. Pheromone components affect motivation and induce persistent modulation of associative learning and memory in honey bees. Commun. Biol. 2020, 3, 447. [Google Scholar] [CrossRef] [PubMed]
- Totani, Y.; Aonuma, H.; Oike, A.; Watanabe, T.; Hatakeyama, D.; Sakakibara, M.; Lukowiak, K.; Ito, E. Monoamines, Insulin and the Roles They Play in Associative Learning in Pond Snails. Front. Behav. Neurosci. 2019, 13, 65. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Ni, J.Z.; Vargas-Velazquez, A.M.; Gu, S.G.; Frøkjær-Jensen, C. Reprogramming the piRNA pathway for multiplexed and transgenerational gene silencing in C. elegans. Nat. Methods 2022, 19, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Ge, A.; Hu, L.; Fan, J.; Ge, M.; Wang, X.; Wang, S.; Feng, X.; Du, W.; Liu, B.-F. A low-cost microfluidic platform coupled with light emitting diode for optogenetic analysis of neuronal response in C. elegans. Talanta 2020, 223, 121646. [Google Scholar] [CrossRef] [PubMed]
- Brandel-Ankrapp, K.L.; Arey, R.N. Uncovering novel regulators of memory using C. elegans genetic and genomic analysis. Biochem. Soc. Trans. 2023, 51, 161–171. [Google Scholar] [CrossRef]
- De Fruyt, N.; Yu, A.J.; Rankin, C.H.; Beets, I.; Chew, Y.L. The role of neuropeptides in learning: Insights from C. elegans. Int. J. Biochem. Cell Biol. 2020, 125, 105801. [Google Scholar] [CrossRef]
- Choi, M.-K.; Liu, H.; Wu, T.; Yang, W.; Zhang, Y. NMDAR-mediated modulation of gap junction circuit regulates olfactory learning in C. elegans. Nat. Commun. 2020, 11, 3467. [Google Scholar] [CrossRef]
- Liao, C.-P.; Chiang, Y.-C.; Tam, W.H.; Chen, Y.-J.; Chou, S.-H.; Pan, C.-L. Neurophysiological basis of stress-induced aversive memory in the nematode Caenorhabditis elegans. Curr. Biol. 2022, 32, 5309–5322.e6. [Google Scholar] [CrossRef]
- Liang, J.J.H.; McKinnon, I.A.; Rankin, C.H. The contribution of C. elegans neurogenetics to understanding neurodegenerative diseases. J. Neurogenet. 2020, 34, 527–548. [Google Scholar] [CrossRef]
- Chen, Z.; Lv, Z.; Zhang, Z.; Weitz, D.A.; Zhang, H.; Zhang, Y.; Cui, W. Advanced microfluidic devices for fabricating multi-structural hydrogel microsphere. Exploration 2021, 1, 20210036. [Google Scholar] [CrossRef]
- Buttkewitz, M.A.; Heuer, C.; Bahnemann, J. Sensor integration into microfluidic systems: Trends and challenges. Curr. Opin. Biotechnol. 2023, 83, 102978. [Google Scholar] [CrossRef]
- Chang, D.; Wang, Z.; Flynn, C.D.; Mahmud, A.; Labib, M.; Wang, H.; Geraili, A.; Li, X.; Zhang, J.; Sargent, E.H.; et al. A high-dimensional microfluidic approach for selection of aptamers with programmable binding affinities. Nat. Chem. 2023, 15, 773–780. [Google Scholar] [CrossRef]
- San-Miguel, A.; Lu, H. Microfluidics as a tool for C. elegans research. Wormbook 2013, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Viri, V.; Arveiler, M.; Lehnert, T.; Gijs, M.A.M. An In Vivo Microfluidic Study of Bacterial Load Dynamics and Absorption in the C. elegans Intestine. Micromachines 2021, 12, 832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.; Yuan, H.; Yong, R.; Duan, S.; Li, Y.; Spencer, J.; Lim, E.G.; Yu, L.; Song, P. Deep Learning for Microfluidic-Assisted Caenorhabditis elegans Multi-Parameter Identification Using YOLOv7. Micromachines 2023, 14, 1339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, H.; Bargmann, C.I. Pathogenic bacteria induce aversive olfactory learning in Caenorhabditis elegans. Nature 2005, 438, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wheeler, A.R. Maze exploration and learning in C. elegans. Lab Chip 2006, 7, 186–192. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Ge, A.; Hu, L.; Du, W.; Liu, B.F. A dual-stimulation strategy in a micro-chip for the investigation of mechanical associative learning behavior of C. elegans. Talanta 2020, 215, 120900. [Google Scholar] [CrossRef]
- Bargmann, C.I. Chemosensation in C. elegans. WormBook 2006, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Goodman, M.B. Mechanosensation. WormBook 2006, 1–14. [Google Scholar] [CrossRef]
- Butler, J.M.; Maruska, K.P. Mechanosensory signaling as a potential mode of communication during social interactions in fishes. J. Exp. Biol. 2016, 219, 2781–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitesides, G.M.; Ostuni, E.; Takayama, S.; Jiang, X.; Ingber, D.E. Soft Lithography in Biology and Biochemistry. Annu. Rev. Biomed. Eng. 2001, 3, 335–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, W.R. Mechanosensory molecules and circuits in C. elegans. Pflügers Arch.-Eur. J. Physiol. 2014, 467, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Kerr, R.; Bianchi, L.; Frokjaer-Jensen, C.; Slone, D.; Xue, J.; Gerstbrein, B.; Driscoll, M.; Schafer, W.R. In vivo imaging of C. elegans mechanosensory neurons demonstrates a specific role for the MEC-4 channel in the process of gentle touch sensation. Neuron 2003, 39, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Árnadóttir, J.; O’Hagan, R.; Chen, Y.; Goodman, M.B.; Chalfie, M. The DEG/ENaC Protein MEC-10 Regulates the Transduction Channel Complex in Caenorhabditis elegans Touch Receptor Neurons. J. Neurosci. 2011, 31, 12695–12704. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.M.; Madsen, D.M.; Kahn-Kirby, A.; Peckol, E.L.; Moulder, G.; Barstead, R.; Maricq, A.V.; Bargmann, C.I. Combinatorial expression of TRPV channel proteins defines their sensory functions and subcellular localization in C. elegans neurons. Neuron 2002, 35, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Feng, Z.; Sternberg, P.W.; Xu, X.Z.S. A C. elegans stretch receptor neuron revealed by a mechanosensitive TRP channel homologue. Nature 2006, 440, 684–687. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; DU, M.; Zhang, Y.; Wang, D. Adverse effects of metal exposure on chemotaxis towards water-soluble attractants regulated mainly by ASE sensory neuron in nematode Caenorhabditis elegans. J. Environ. Sci. 2009, 21, 1684–1694. [Google Scholar] [CrossRef]
- Iliadi, K.G.; Iliadi, N.; Boulianne, G.L. Drosophila mutants lacking octopamine exhibit impairment in aversive olfactory associative learning. Eur. J. Neurosci. 2017, 46, 2080–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Wang, Y.; Tang, S.; Su, H.; Wang, X.; Du, W.; Wang, Y.; Liu, B.-F. A PDMS–Agar Hybrid Microfluidic Device for the Investigation of Chemical–Mechanical Associative Learning Behavior of C. elegans. Micromachines 2023, 14, 1576. https://doi.org/10.3390/mi14081576
Zhu J, Wang Y, Tang S, Su H, Wang X, Du W, Wang Y, Liu B-F. A PDMS–Agar Hybrid Microfluidic Device for the Investigation of Chemical–Mechanical Associative Learning Behavior of C. elegans. Micromachines. 2023; 14(8):1576. https://doi.org/10.3390/mi14081576
Chicago/Turabian StyleZhu, Jinchi, Yu Wang, Shuting Tang, Huiying Su, Xixian Wang, Wei Du, Yun Wang, and Bi-Feng Liu. 2023. "A PDMS–Agar Hybrid Microfluidic Device for the Investigation of Chemical–Mechanical Associative Learning Behavior of C. elegans" Micromachines 14, no. 8: 1576. https://doi.org/10.3390/mi14081576