
Cyborg Moth Flight Control Based on Fuzzy Deep Learning

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- We present a noninvasive cyborg moth design approach based on fuzzy deep learning for flight control;
- We propose a novel hierarchical fuzzy deep learning model that effectively learns the species common behaviors, group-specific behaviors, and individual-specific behaviors to achieve a high control success rate.
- We propose a new fuzzy clustering method based on Pythagorean-type fuzzy sets for moth grouping.
- The proposed approach can be easily extended for behavior learning of other cyborg animals and, therefore, contributes to the development of biobots.
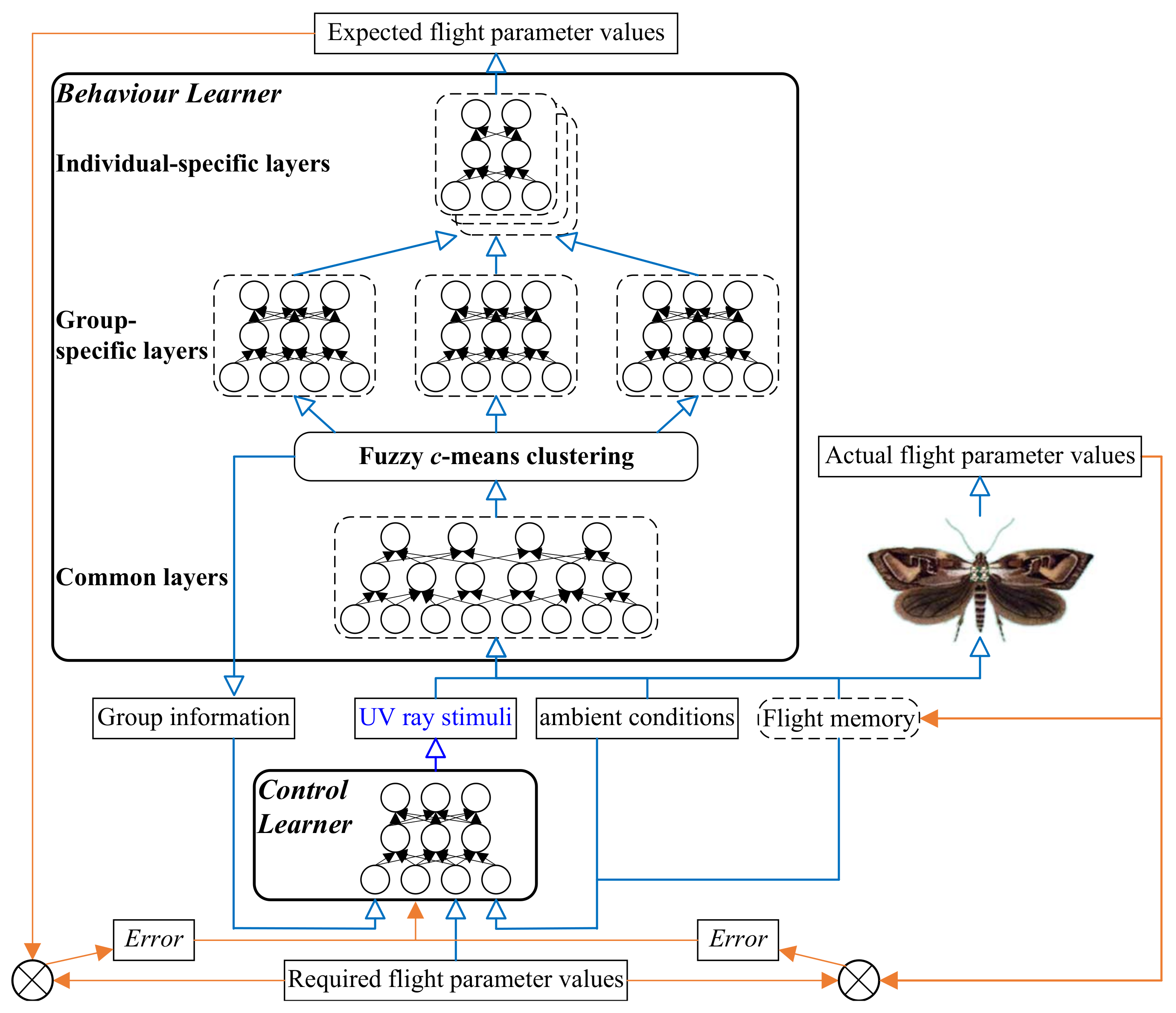
2. Overview of the Model for Cyborg Flight Control
2.1. Model Architecture
- The UV ray stimulation, which is described by 32 variables, i.e., the light intensities, exposure durations, pattern moving velocities (in the x-, y-, and z-axes), and pattern moving distances (in the x-, y-, and z-axes) of the four lamps.
- The ambient conditions, which are described by 34 variables, i.e., temperature, humidity, atmospheric pressure, oxygen concentration, carbon dioxide concentration, horizontal and vertical wind speeds, and the light duration, intensity, and illuminance of nine different wavelengths/colors (UVA, UVB, UVC, violet, indigo, blue, yellow, green, orange) to which the insect is sensitive (note that our study assumes that the ambient wind speed does not exceed 1.5 m/s; if the wind is too strong, then it is impossible to control the flight path of a moth).
2.2. Pythagorean Fuzzy Deep Denoising Autoencoder
3. Behavior Leaner
3.1. Hierarchical Learning of Cyborg Flight Behaviors
3.1.1. Learning the Species Common Behaviors
3.1.2. Learning Group-Specific Behaviors
3.1.3. Learning Individual-Specific Behaviors
3.2. Pythagorean Fuzzy c-Means Clustering for Moth Grouping
| Algorithm 1: Pythagorean fuzzy c-means clustering algorithm. |
 |
4. Control Leaner
5. Experiments
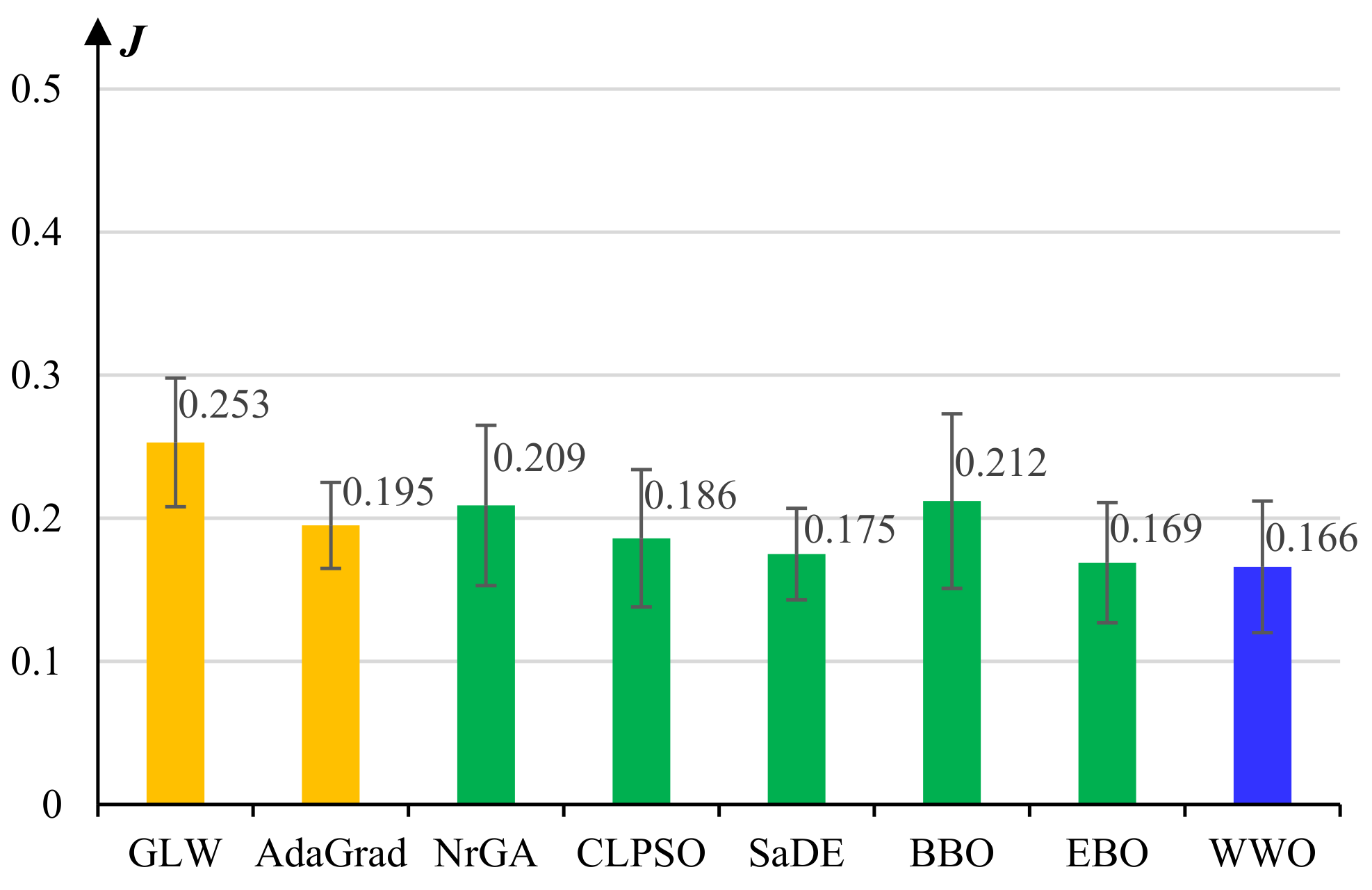
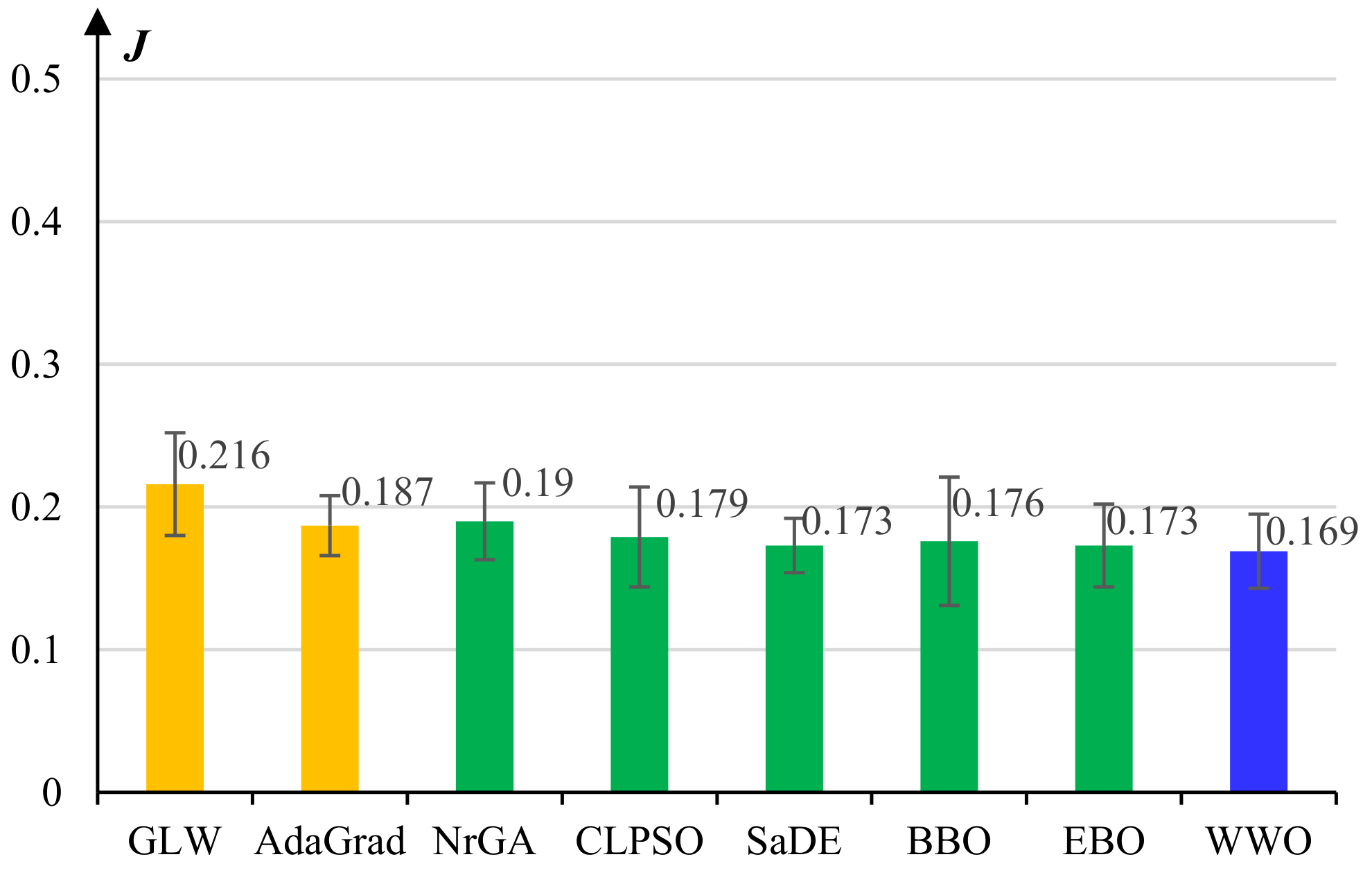
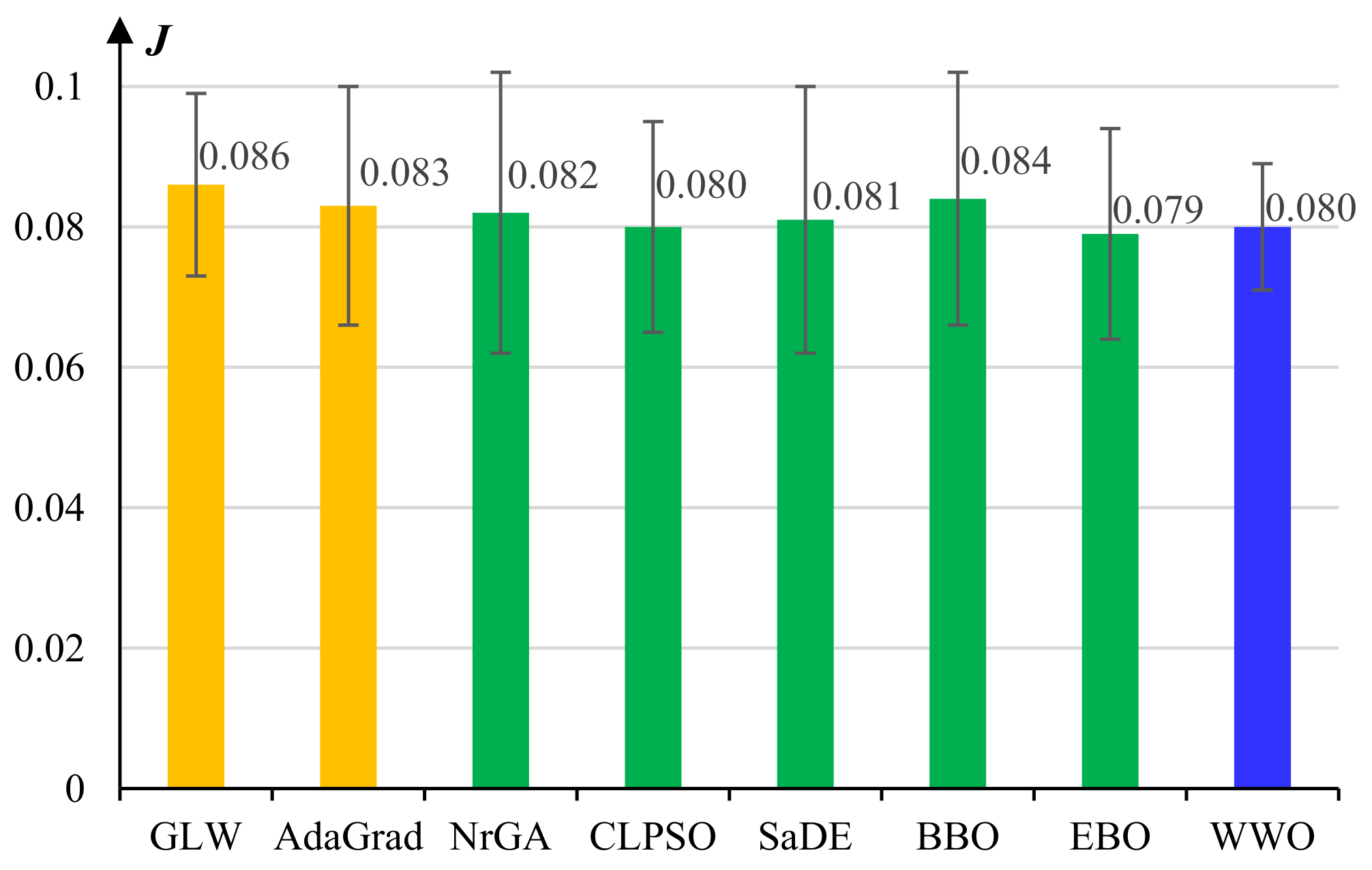
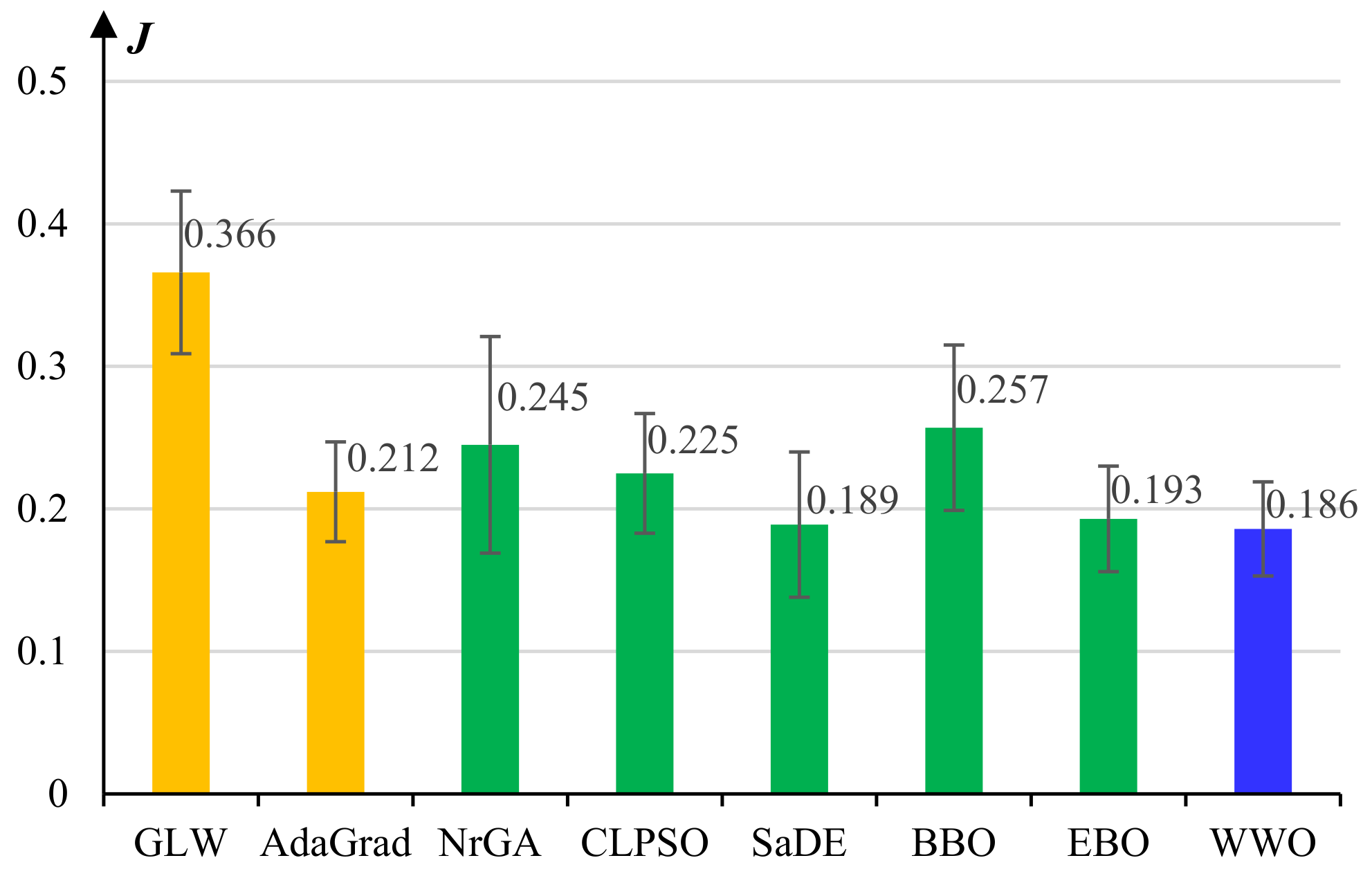
5.1. Experiments on Model Pretraining
- The basic gradient-based layer-wise (GLW) algorithm [28].
- An adaptive gradient (AdaGrad) algorithm [49].
- A non-revisiting genetic algorithm with adaptive mutation (NrGA) [50].
- A comprehensive learning PSO (CLPSO) algorithm [51] where each solution learns from different exemplars at different dimensions.
- A self-adaptive differential evolution (SaDE) algorithm [52], which adaptively chooses more prospective evolution strategies among a set of candidate strategies.
- A biogeography-based optimization (BBO) algorithm [53], which evolves solutions by continuously migrating features from high-fitness individuals to low-fitness ones based on a biogeographical migration model.
- An improved BBO algorithm called ecogeography-based optimization (EBO) [54], which defines two migration operators, namely global migration and local migration, that are adaptively applied according to the maturity of the population.
- The WWO algorithm [42].
- The three layers of the lower common learner have 80, 46, and 26 neurons, respectively;
- The three layers of each group-specific learner have 26, 15, and 9 neurons, respectively;
- The three layers of each individual-specific learner have 9, 6, and 7 neurons, respectively;
- The four layers of the Control Learner have 75, 49, 35, and 32 neurons, respectively.
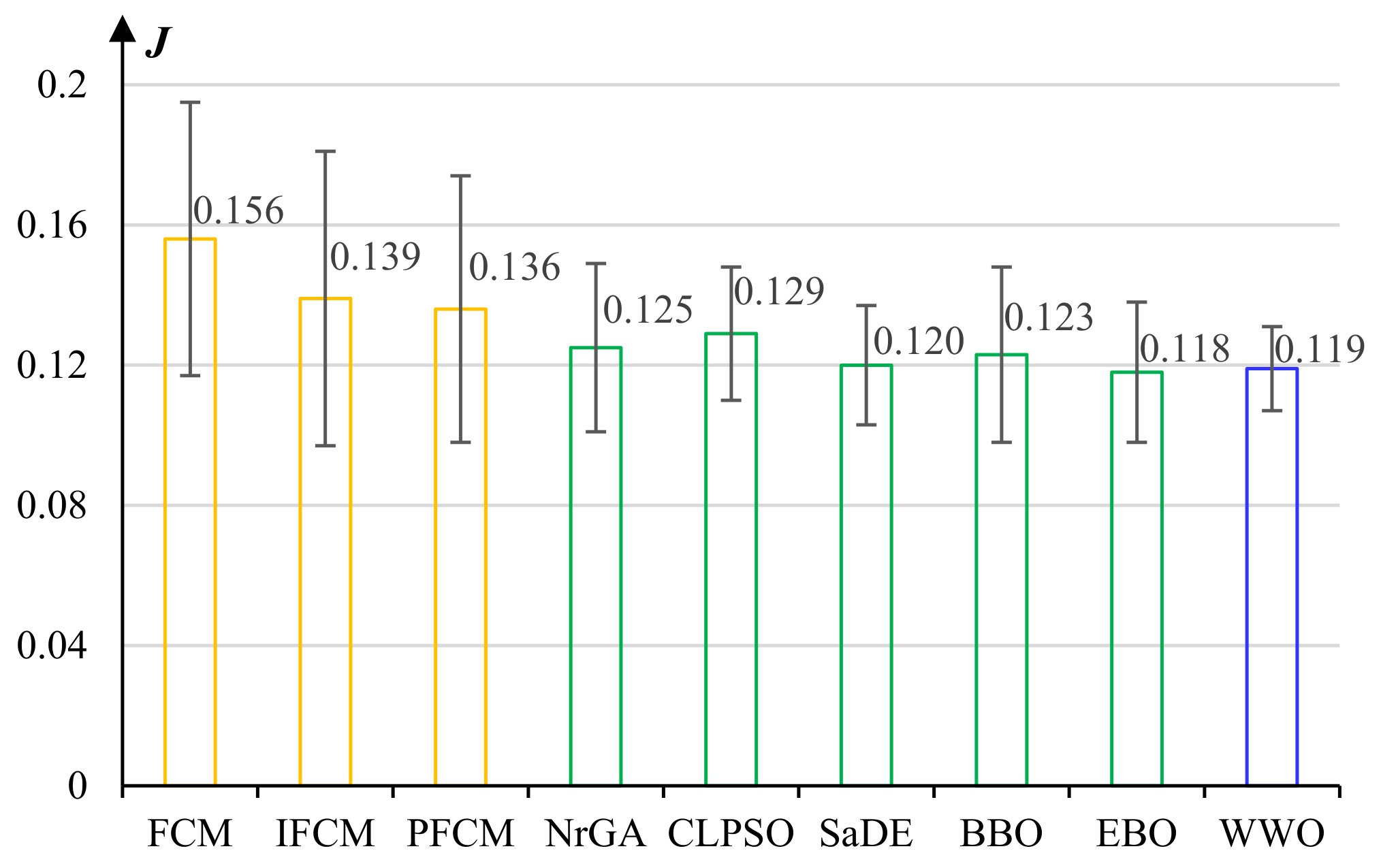
5.2. Experiments on Fuzzy c-Means Clustering
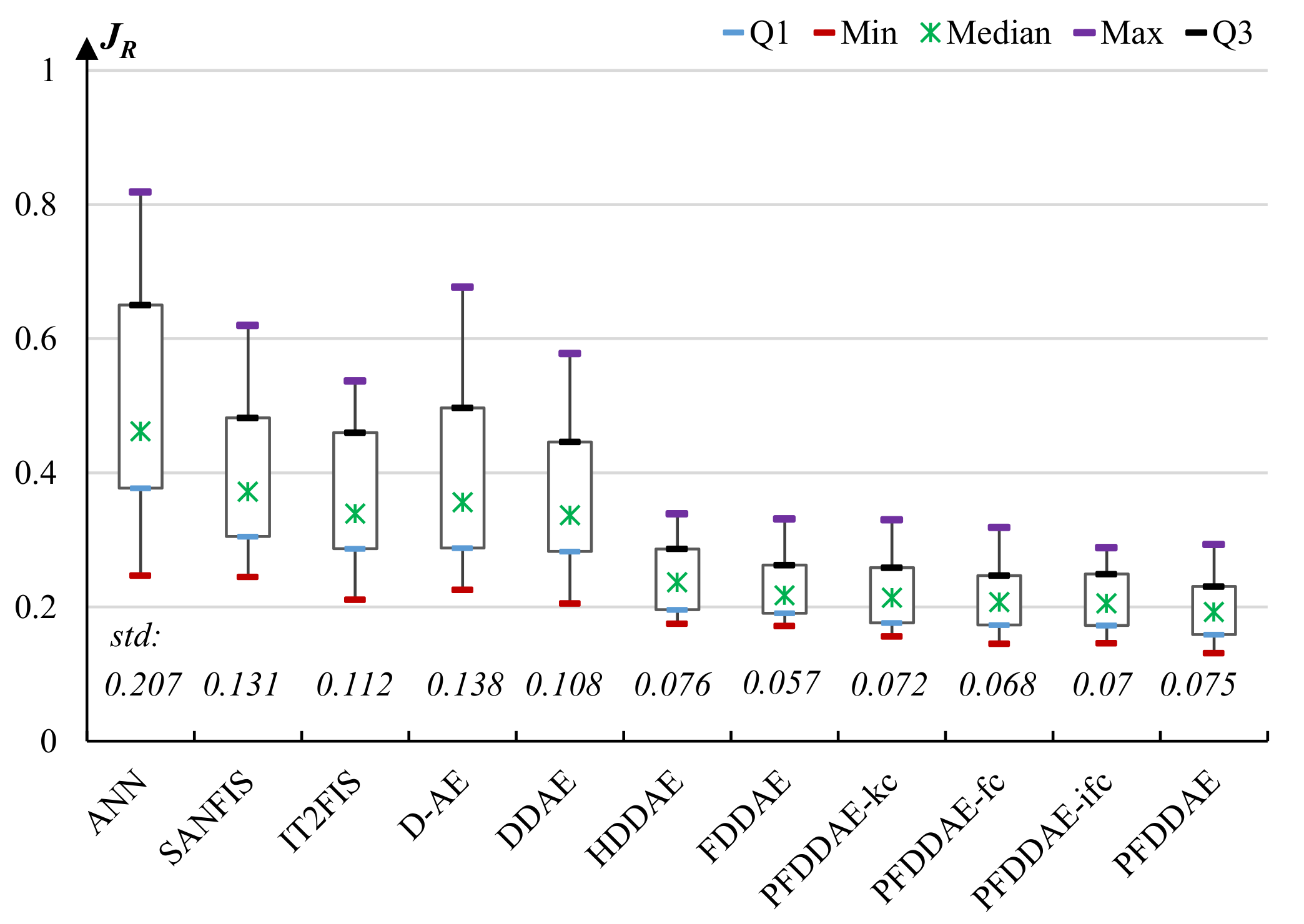
5.3. Experiments on Cyborg Flight Behavior Learning
- A standard three-layer back-propagation artificial neural network (ANN). The numbers of neurons in the three layers are tuned to 80, 25, and 7, respectively.
- A basic self-adaptive neuro-fuzzy inference system (SANFIS) trained by an agglomerative clustering algorithm and a recursive least-squares algorithm [55].
- An evolving interval type-2 neuro-fuzzy inference system (IT2FIS) trained by a metacognitive sequential learning algorithm [56].
- A basic and integrated deep AE model (denoted by D-AE) [57], which employs the basic AE without a denoising mechanism as the building block and does not hierarchically divide the model into common learner, group-specific learners and individual-specific learners. After fine-tuning, the number of layers of D-AE is set to five, and the numbers of neurons in the three hidden layers are set to 48, 27, and 13, respectively.
- A basic and integrated deep DAE model (denoted by DDAE) [37], which uses the same structure as D-AE but employs DAE as the building block.
- A basic hierarchical deep DAE (denoted by HDDAE) that uses three hierarchies as described in Section 2 but does not employ fuzzy model parameters.
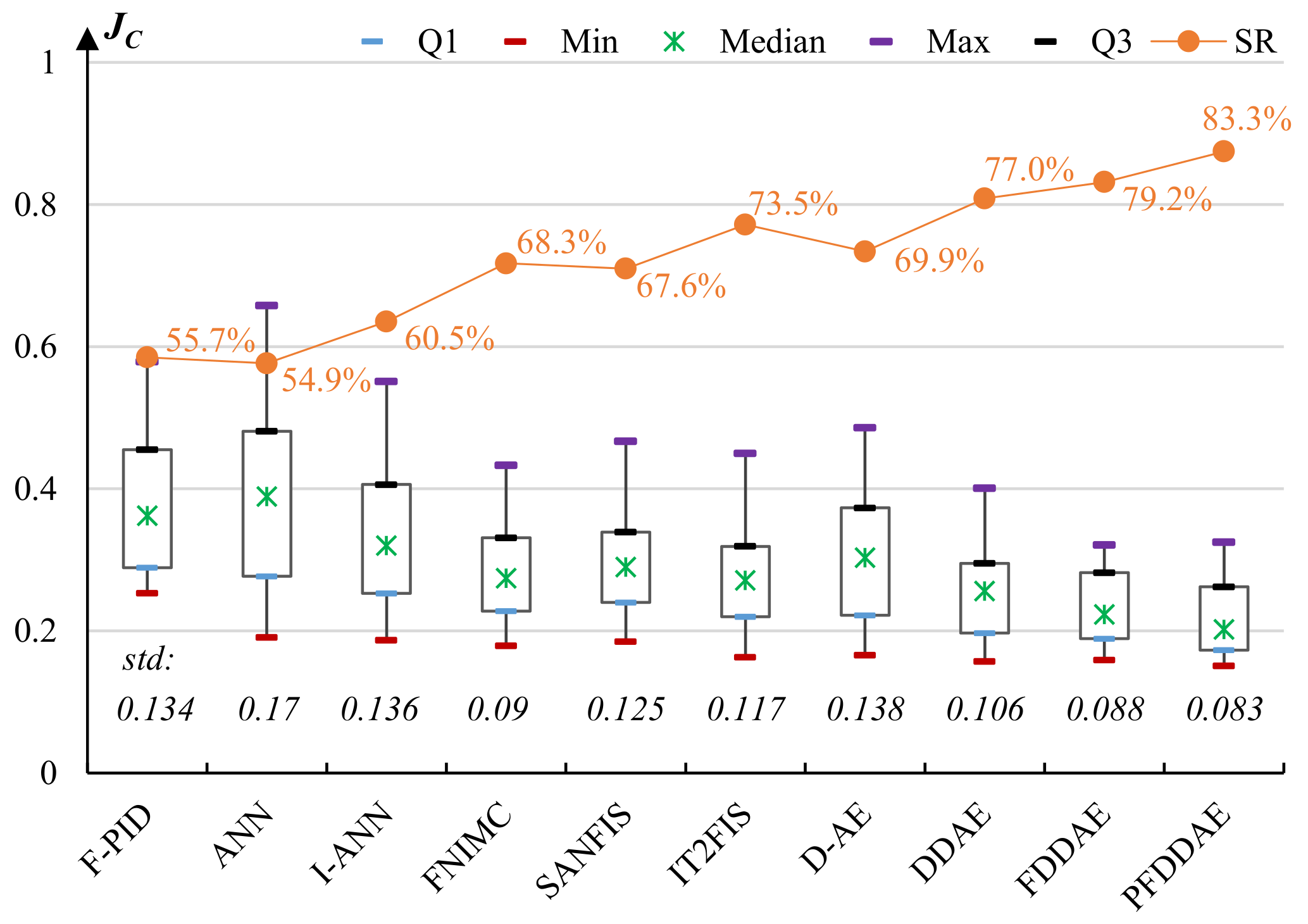
5.4. Experiments on Cyborg Flight Control
6. Conclusions and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| AdaGrad | Adaptive gradient |
| AE | Autoencoder |
| ANN | Artificial neural network |
| BBO | Biogeography-based optimization |
| DAE | Denoising autoencoder |
| DDAE | Deep denoising autoencoder |
| EBO | Ecogeography-based optimization |
| FCM | Fuzzy c-means clustering |
| GLW | Gradient-based layer-wise |
| IT2FIS | Interval type-2 neuro-fuzzy inference system |
| LED | Light-emitting diode |
| MLR | Multivariable linear regression |
| PFDDAE | Pythagorean fuzzy deep denoising autoencoder |
| PFN | Pythagorean-type fuzzy number |
| PSO | Particle swam optimization |
| SANFIS | Self-adaptive neuro-fuzzy inference system |
| UV | Ultraviolet |
| WWO | Water wave optimization |
References
- Dutta, A. Cyborgs: Neuromuscular Control of Insects. In Proceedings of the 9th International IEEE/EMBS Conference on Neural Engineering, San Francisco, CA, USA, 20–23 March 2019; pp. 682–685. [Google Scholar] [CrossRef]
- Fu, Y.; Yu, H.; Zhang, X.; Malgaretti, P.; Kishore, V.; Wang, W. Microscopic Swarms: From Active Matter Physics to Biomedical and Environmental Applications. Micromachines 2022, 13, 295. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.J.; Wang, Y.; Ling, H.F.; Xue, Y.; Chen, S.Y. Integrated civilian-military pre-positioning of emergency supplies: A multiobjective optimization approach. Appl. Soft Comput. 2017, 58, 732–741. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Du, Y.C.; Su, Z.L.; Ling, H.F.; Zhang, M.X.; Chen, S.Y. Evolutionary human-UAV cooperation for transmission network restoration. IEEE Trans. Ind. Inf. 2021, 17, 1648–1657. [Google Scholar] [CrossRef]
- Romano, D.; Donati, E.; Benelli, G.; Stefanini, C. A review on animal–robot interaction: From bio-hybrid organisms to mixed societies. Biol. Cybern. 2019, 113, 201–225. [Google Scholar] [CrossRef]
- Holzer, R.; Shimoyama, I. Locomotion control of a bio-robotic system via electric stimulation. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robot and Systems (IROS’97), Grenoble, France, 11 September 1997; Volume 3, pp. 1514–1519. [Google Scholar] [CrossRef]
- Sanchez, C.J.; Chiu, C.W.; Zhou, Y.; González, J.M.; Vinson, S.B.; Liang, H. Locomotion control of hybrid cockroach robots. J. R. Soc. Interface 2015, 12, 20141363. [Google Scholar] [CrossRef]
- Erickson, J.C.; Herrera, M.; Bustamante, M.; Shingiro, A.; Bowen, T. Effective Stimulus Parameters for Directed Locomotion in Madagascar Hissing Cockroach Biobot. PLoS ONE 2015, 10, e0134348. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zhang, D. Brain-Computer Interface Controlling Cyborg: A Functional Brain-to-Brain Interface Between Human and Cockroach. In Brain-Computer Interface Research; Springer: Cham, Switzerland, 2017; pp. 71–79. [Google Scholar] [CrossRef]
- Sato, H.; Berry, C.W.; Casey, B.E.; Lavella, G.; Yao, Y.; VandenBrooks, J.M.; Maharbiz, M.M. A cyborg beetle: Insect flight control through an implantable, tetherless microsystem. In Proceedings of the IEEE 21st International Conference on Micro Electro Mechanical System, Tucson, AZ, USA, 13–17 January 2008; pp. 164–167. [Google Scholar] [CrossRef]
- Vo Doan, T.T.; Li, Y.; Cao, F.; Sato, H. Cyborg beetle: Thrust control of free flying beetle via a miniature wireless neuromuscular stimulator. In Proceedings of the 28th IEEE International Conference on Micro Electro Mechanical Systems (MEMS), Estoril, Portugal, 18–22 January 2015; pp. 1048–1050. [Google Scholar] [CrossRef]
- Cao, F.; Zhang, C.; Choo, H.Y.; Sato, H. Insect-computer hybrid legged robot with user-adjustable speed, step length and walking gait. J. R. Soc. Interface 2016, 13, 20160060. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.D.; Tan, P.Z.; Sato, H.; Vo-Doan, T.T. Sideways Walking Control of a Cyborg Beetle. IEEE Trans. Med. Robot. Bionics 2020, 2, 331–337. [Google Scholar] [CrossRef]
- Bozkurt, A.; Gilmour, R.F., Jr.; Lal, A. Balloon-Assisted Flight of Radio-Controlled Insect Biobots. IEEE Trans. Bio-Med. Eng. 2009, 56, 2304–2307. [Google Scholar] [CrossRef]
- Tsang, W.M.; Stone, A.; Aldworth, Z.; Otten, D.; Akinwande, A.I.; Daniel, T.; Hildebrand, J.G.; Levine, R.B.; Voldman, J. Remote control of a cyborg moth using carbon nanotube-enhanced flexible neuroprosthetic probe. In Proceedings of the IEEE 23rd International Conference on Micro Electro Mechanical Systems (MEMS 2010), Wanchai, Hong Kong, 24–28 January 2010; pp. 39–42. [Google Scholar] [CrossRef]
- Schwefel, J.; Ritzmann, R.E.; Lee, I.N.; Pollack, A.; Weeman, W.; Garverick, S.; Willis, M.; Rasmussen, M.; Scherson, D. Wireless Communication by an Autonomous Self-Powered Cyborg Insect. J. Electrochem. Soc. 2015, 161, 3113–3116. [Google Scholar] [CrossRef]
- Giampalmo, S.L.; Absher, B.F.; Bourne, W.T.; Steves, L.E.; Vodenski, V.V.; O’Donnell, P.M.; Erickson, J.C. Generation of complex motor patterns in american grasshopper via current-controlled thoracic electrical interfacing. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Boston, MA, USA, 30 August–3 September 2011; pp. 1275–1278. [Google Scholar] [CrossRef]
- Mehta, D.; Altan, E.; Chandak, R.; Raman, B.; Chakrabartty, S. Behaving cyborg locusts for standoff chemical sensing. In Proceedings of the IEEE International Symposium on Circuits and Systems (ISCAS), Baltimore, MD, USA, 28–31 May 2017; pp. 1–4. [Google Scholar] [CrossRef]
- Bao, L.; Zheng, N.; Zhao, H.; Hao, Y.; Zheng, H.; Hu, F.; Zheng, X. Flight control of tethered honeybees using neural electrical stimulation. In Proceedings of the 5th International IEEE/EMBS Conference on Neural Engineering, Cancun, Mexico, 27 April–1 May 2011; pp. 558–561. [Google Scholar] [CrossRef]
- Wang, S.; Shen, L.; Liu, X.; Liao, H. A wearable backpack chip for honeybee biorobot. In Proceedings of the China Semiconductor Technology International Conference (CSTIC), Shanghai, China, 13–14 March 2016; pp. 1–3. [Google Scholar] [CrossRef]
- Jamali, M.; Jamali, Y.; Golshani, M. Theory of cyborg: A new approach to fish locomotion control. arXiv 2019, arXiv:1904.12460. [Google Scholar]
- Montrose, V.; Carroll, G.; Smith, R.; Oxley, J. Cyborg Insects: Use or Abuse? Department of Animal and Agriculture, University Centre Hartpury: Hartpury, UK, 2017. [Google Scholar]
- Zheng, N.; Jin, M.; Hong, H.; Huang, L.; Gu, Z.; Li, H. Real-time and precise insect flight control system based on virtual reality. Electr. Lett. 2017, 53, 387–389. [Google Scholar] [CrossRef]
- Zheng, N.; Ma, Q.; Jin, M.; Zhang, S.; Guan, N.; Yang, Q.; Dai, J. Abdominal-Waving Control of Tethered Bumblebees Based on Sarsa with Transformed Reward. IEEE Trans. Cybern. 2019, 49, 3064–3073. [Google Scholar] [CrossRef]
- Wu, G.D.; Zhu, Z.W.; Huang, P.H. A TS-Type Maximizing-Discriminability-Based Recurrent Fuzzy Network for Classification Problems. IEEE Trans. Fuzzy Syst. 2011, 19, 339–352. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Ling, H.F.; Chen, S.Y.; Xue, J.Y. A hybrid neuro-fuzzy network based on differential biogeography-based optimization for online population classification in earthquakes. IEEE Trans. Fuzzy Syst. 2015, 23, 1070–1083. [Google Scholar] [CrossRef]
- Vincent, P.; Larochelle, H.; Bengio, Y.; Manzagol, P.A. Extracting and Composing Robust Features with Denoising Autoencoders. In Proceedings of the International Conference on Machine Learning, Helsinki, Finland, 5–9 July 2008; ACM: New York, NY, USA, 2008; pp. 1096–1103. [Google Scholar] [CrossRef] [Green Version]
- Bengio, Y.; Lamblin, P.; Popovici, D.; Larochelle, H. Greedy layer-wise training of deep networks. In Advances in Neural Information Processing Systems (NIPS’06); Bernhard Schölkopf, J.P., Hoffman, T., Eds.; MIT Press: Cambridge, MA, USA, 2007; Volume 19, pp. 153–160. [Google Scholar]
- Chen, C.; Zhang, C.Y.; Chen, L.; Gan, M. Fuzzy Restricted Boltzmann Machine for the Enhancement of Deep Learning. IEEE Trans. Fuzzy Syst. 2015, 23, 2163–2173. [Google Scholar] [CrossRef]
- Yager, R. Pythagorean fuzzy subsets. In Proceedings of the Joint IFSA World Congress and NAFIPS Annual Meeting IFSA/NAFIPS, Edmonton, AB, Canada, 24–28 June 2013; pp. 57–61. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Z. Extension of TOPSIS to multiple criteria decision making with pythagorean fuzzy sets. Int. J. Intell. Syst. 2014, 29, 1061–1078. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Chen, S.Y.; Xue, Y.; Xue, J.Y. A Pythagorean-type fuzzy deep denoising autoencoder for industrial accident early warning. IEEE Trans. Fuzzy Syst. 2017, 25, 1561–1575. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Sheng, W.G.; Sun, X.M.; Chen, S.Y. Airline passenger profiling based on fuzzy deep machine learning. IEEE Trans. Neural Netw. Learn. Syst. 2017, 28, 2911–2923. [Google Scholar] [CrossRef]
- Gou, X.; Xu, Z.; Liao, H. Exponential operations of interval-valued intuitionistic fuzzy numbers. Int. J. Mach. Learn. Cybern. 2016, 7, 501–518. [Google Scholar] [CrossRef]
- Garg, H. New exponential operational laws and their aggregation operators for interval-valued Pythagorean fuzzy multicriteria decision-making. Int. J. Intell. Syst. 2018, 33, 653–683. [Google Scholar] [CrossRef]
- Hung, W.L.; Wu, J.W. Correlation of intuitionistic fuzzy sets by centroid method. Inform. Sci. 2002, 144, 219–225. [Google Scholar] [CrossRef]
- Vincent, P.; Larochelle, H.; Lajoie, I.; Bengio, Y.; Manzagol, P.A. Stacked denoising autoencoders: Learning useful representations in a deep network with a local denoising criterion. J. Mach. Learn. Res. 2010, 11, 3371–3408. [Google Scholar]
- Sharkawy, A.N. Principle of neural network and its main types. J. Adv. Appl. Comput. Math. 2020, 7, 8–19. [Google Scholar] [CrossRef]
- Yao, X. A review of evolutionary artificial neural networks. Int. J. Intell. Syst. 1993, 8, 539–567. [Google Scholar] [CrossRef]
- Wojtowytsch, S.; E, W. Can Shallow Neural Networks Beat the Curse of Dimensionality? A mean field training perspective. IEEE Trans. Artif. Intell. 2021, 1, 121–129. [Google Scholar] [CrossRef]
- Zheng, Y.J. Water wave optimization: A new nature-inspired metaheuristic. Comput. Oper. Res. 2015, 55, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.H.; Zhang, M.X.; Xu, Z.G.; Cai, C.Y.; Huang, Y.J.; Zheng, Y.J. Shallow and deep neural network training by water wave optimization. Swarm Evol. Comput. 2019, 50, 1–13. [Google Scholar] [CrossRef]
- Sarto i Monteys, V.; Acín, P.; Rosell, G.; Quero, C.; Jiménez, M.A.; Guerrero, A. Moths behaving like butterflies. Evolutionary loss of long range attractant pheromones in castniid moths: A Paysandisia archon model. PLoS ONE 2012, 7, e29282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezdek, J. Pattern Recognition with Fuzzy Objective Function Algorithms; Plenum Press: Logan, UT, USA, 1981. [Google Scholar]
- Xu, Z.; Wu, J. Intuitionistic fuzzy C-means clustering algorithms. J. Syst. Eng. Electron. 2010, 20, 580–590. [Google Scholar] [CrossRef]
- Ren, P.; Xu, Z.; Gou, X. Pythagorean fuzzy TODIM approach to multi-criteria decision making. Appl. Soft Comput. 2016, 42, 246–259. [Google Scholar] [CrossRef]
- Gong, M.; Liang, Y.; Shi, J.; Ma, W.; Ma, J. Fuzzy C-Means Clustering with Local Information and Kernel Metric for Image Segmentation. IEEE Trans. Image Process. 2013, 22, 573–584. [Google Scholar] [CrossRef]
- Zhang, M.; Jiang, W.; Zhou, X.; Xue, Y.; Chen, S. A hybrid biogeography-based optimization and fuzzy C-means algorithm for image segmentation. Soft Comput. 2019, 23, 2033–2046. [Google Scholar] [CrossRef]
- Duchi, J.; Hazan, E.; Singer, Y. Adaptive subgradient methods for online learning and stochastic optimization. J. Mach. Learn. Res. 2011, 12, 2121–2159. [Google Scholar]
- Yuen, S.Y.; Chow, C.K. A Genetic Algorithm That Adaptively Mutates and Never Revisits. IEEE Trans. Evol. Comput. 2009, 13, 454–472. [Google Scholar] [CrossRef]
- Liang, J.J.; Qin, A.K.; Suganthan, P.; Baskar, S. Comprehensive learning particle swarm optimizer for global optimization of multimodal functions. IEEE Trans. Evol. Comput. 2006, 10, 281–295. [Google Scholar] [CrossRef]
- Qin, A.K.; Huang, V.L.; Suganthan, P. Differential Evolution Algorithm with Strategy Adaptation for Global Numerical Optimization. IEEE Trans. Evol. Comput. 2009, 13, 398–417. [Google Scholar] [CrossRef]
- Simon, D. Biogeography-Based Optimization. IEEE Trans. Evol. Comput. 2008, 12, 702–713. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.J.; Ling, H.F.; Xue, J.Y. Ecogeography-Based Optimization: Enhancing Biogeography-Based Optimization with Ecogeographic Barriers and Differentiations. Comput. Oper. Res. 2014, 50, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Lee, C.S.G. Self-adaptive neuro-fuzzy inference systems for classification applications. IEEE Trans. Fuzzy Syst. 2002, 10, 790–802. [Google Scholar] [CrossRef]
- Das, A.K.; Subramanian, K.; Sundaram, S. An Evolving Interval Type-2 Neurofuzzy Inference System and Its Metacognitive Sequential Learning Algorithm. IEEE Trans. Fuzzy Syst. 2015, 23, 2080–2093. [Google Scholar] [CrossRef]
- Hinton, G.E.; Salakhutdinov, R.R. Reducing the dimensionality of data with neural networks. Science 2006, 313, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.S.; Man, K.F.; Chen, G.; Kwong, S. An optimal fuzzy PID controller. IEEE Trans. Ind. Electron. 2001, 48, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Hunt, K.; Sbarbaro, D. Neural networks for nonlinear internal model control. IEE Proc. D Control Theory Appl. 1991, 138, 431–438. [Google Scholar] [CrossRef]
- Boukezzoula, R.; Galichet, S.; Foulloy, L. Nonlinear internal model control: Application of inverse model based fuzzy control. IEEE Trans. Fuzzy Syst. 2003, 11, 814–829. [Google Scholar] [CrossRef]
- Le Thi Thuy, N.; Nguyen Trong, T. The Multitasking System of Swarm Robot based on Null-Space-Behavioral Control Combined with Fuzzy Logic. Micromachines 2017, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Du, Y.; Ling, H.; Sheng, W.; Chen, S. Evolutionary collaborative human-UAV search for escaped criminals. IEEE Trans. Evol. Comput. 2020, 24, 217–231. [Google Scholar] [CrossRef]
- Song, Q.; Zheng, Y.J.; Sheng, W.G.; Yang, J. Tridirectional transfer learning for predicting gastric cancer morbidity. IEEE Trans. Neural Netw. Learn. Syst. 2021, 32, 561–574. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Jiang, X.-L.; Su, Z.-L.; Wang, B. Cyborg Moth Flight Control Based on Fuzzy Deep Learning. Micromachines 2022, 13, 611. https://doi.org/10.3390/mi13040611
Yang X, Jiang X-L, Su Z-L, Wang B. Cyborg Moth Flight Control Based on Fuzzy Deep Learning. Micromachines. 2022; 13(4):611. https://doi.org/10.3390/mi13040611
Chicago/Turabian StyleYang, Xiao, Xun-Lin Jiang, Zheng-Lian Su, and Ben Wang. 2022. "Cyborg Moth Flight Control Based on Fuzzy Deep Learning" Micromachines 13, no. 4: 611. https://doi.org/10.3390/mi13040611