
Trophoblast Migration with Different Oxygen Levels in a Gel-Patterned Microfluidic System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Gelatin Methacrylate Solution
2.3. Device Fabrication
2.4. Gel Patterning and Cell Seeding
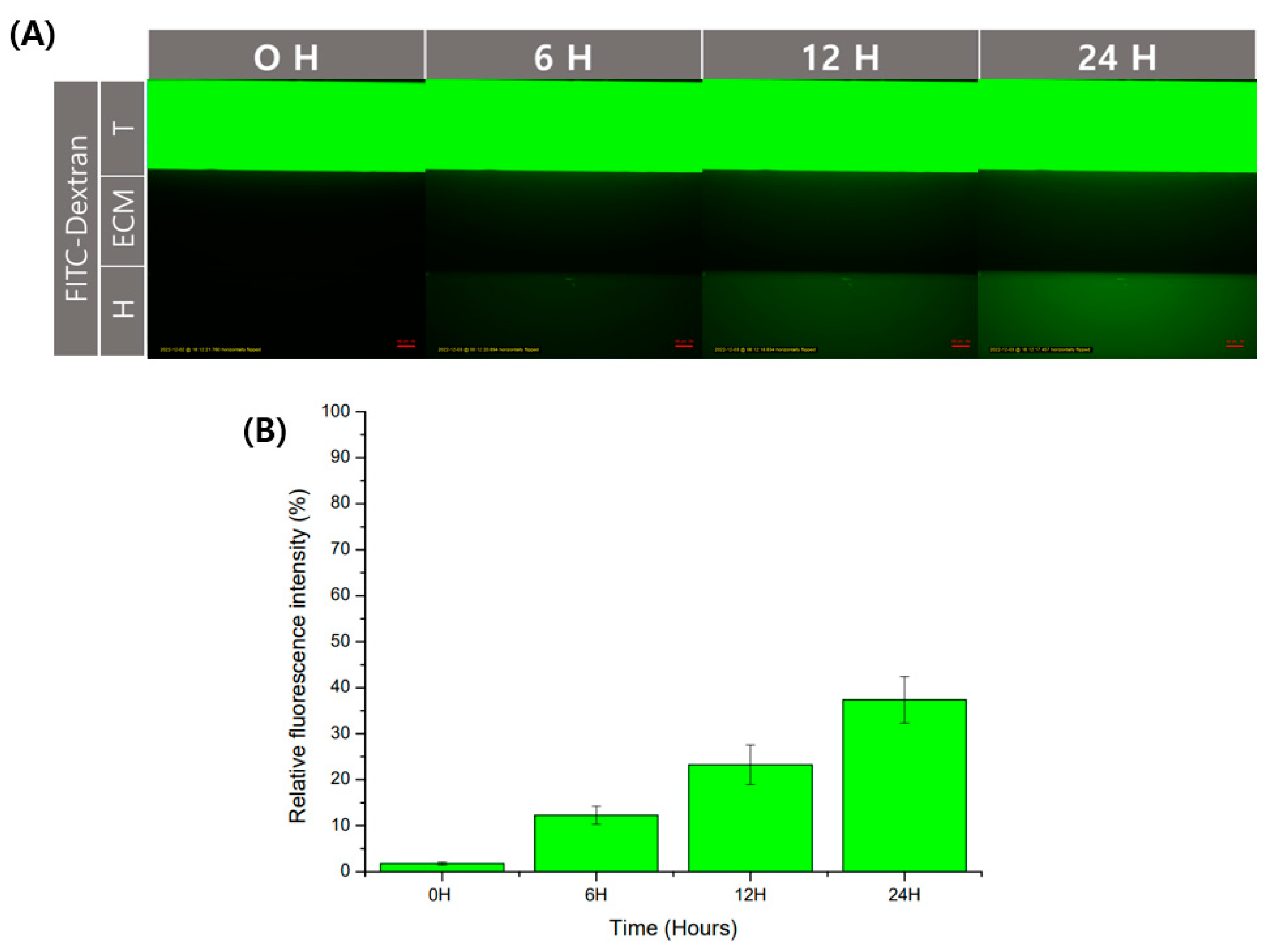
2.5. Molecular Diffusion Analysis
2.6. Cell Tracker Staining
2.7. Quantitative Real-Time PCR (qRT-PCR)
3. Results and Discussion
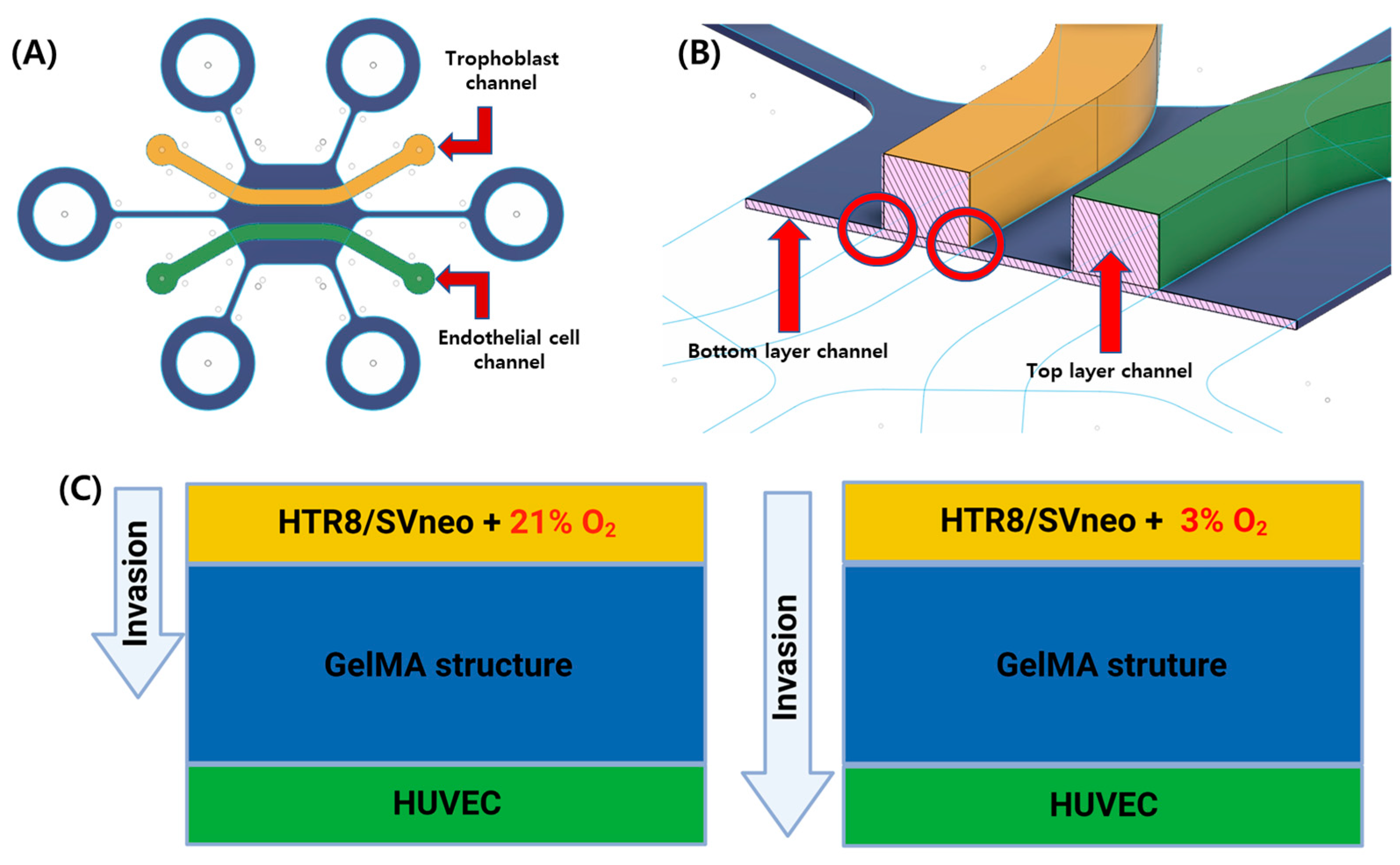
3.1. Gel Patterning in a Microfluidic Chip
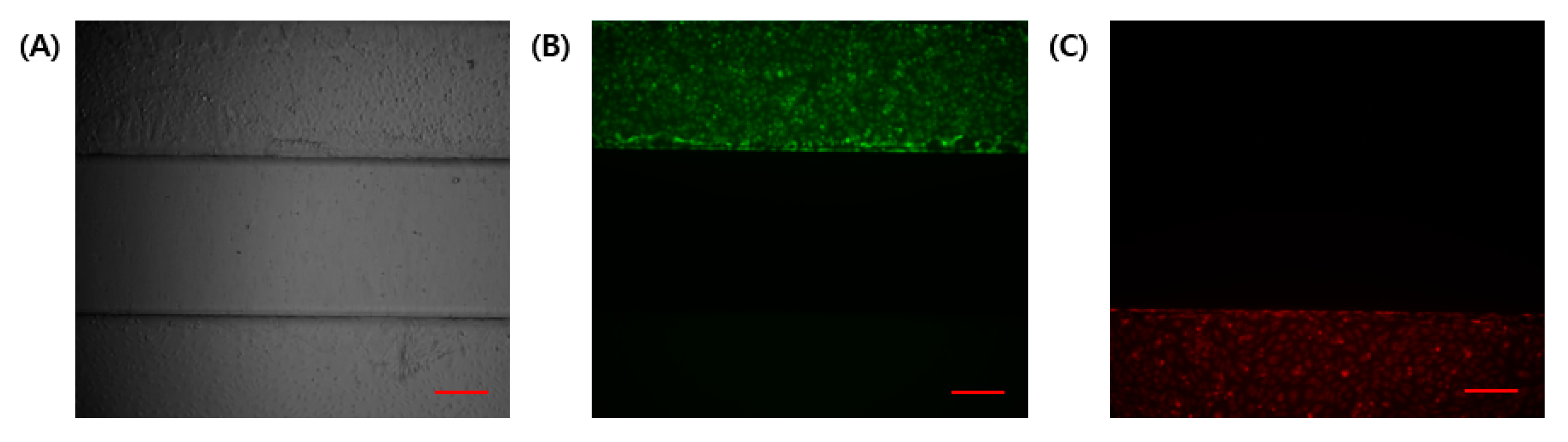
3.2. Cell Separation and Molecular Diffusion of GelMA Structure
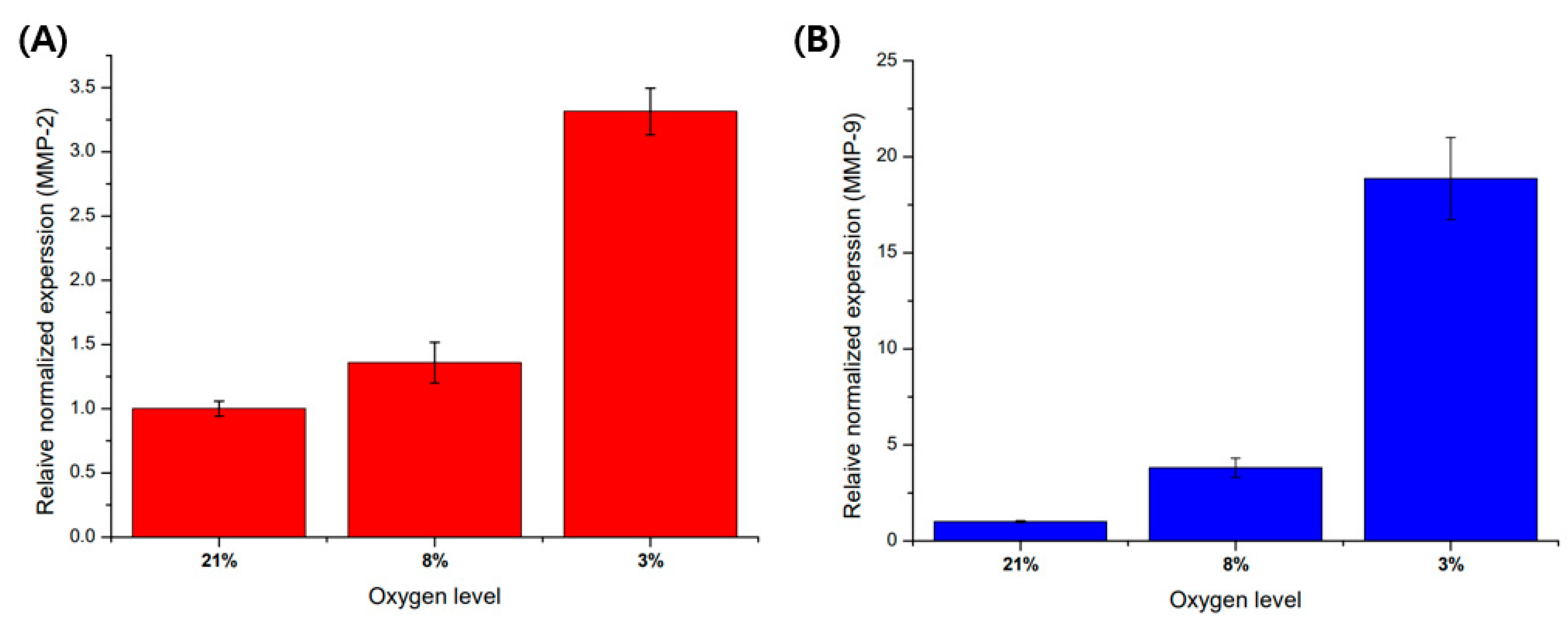
3.3. Hypoxia Promote MMP-2 and MMP-9 mRNA Expression in HTR8/SVneo
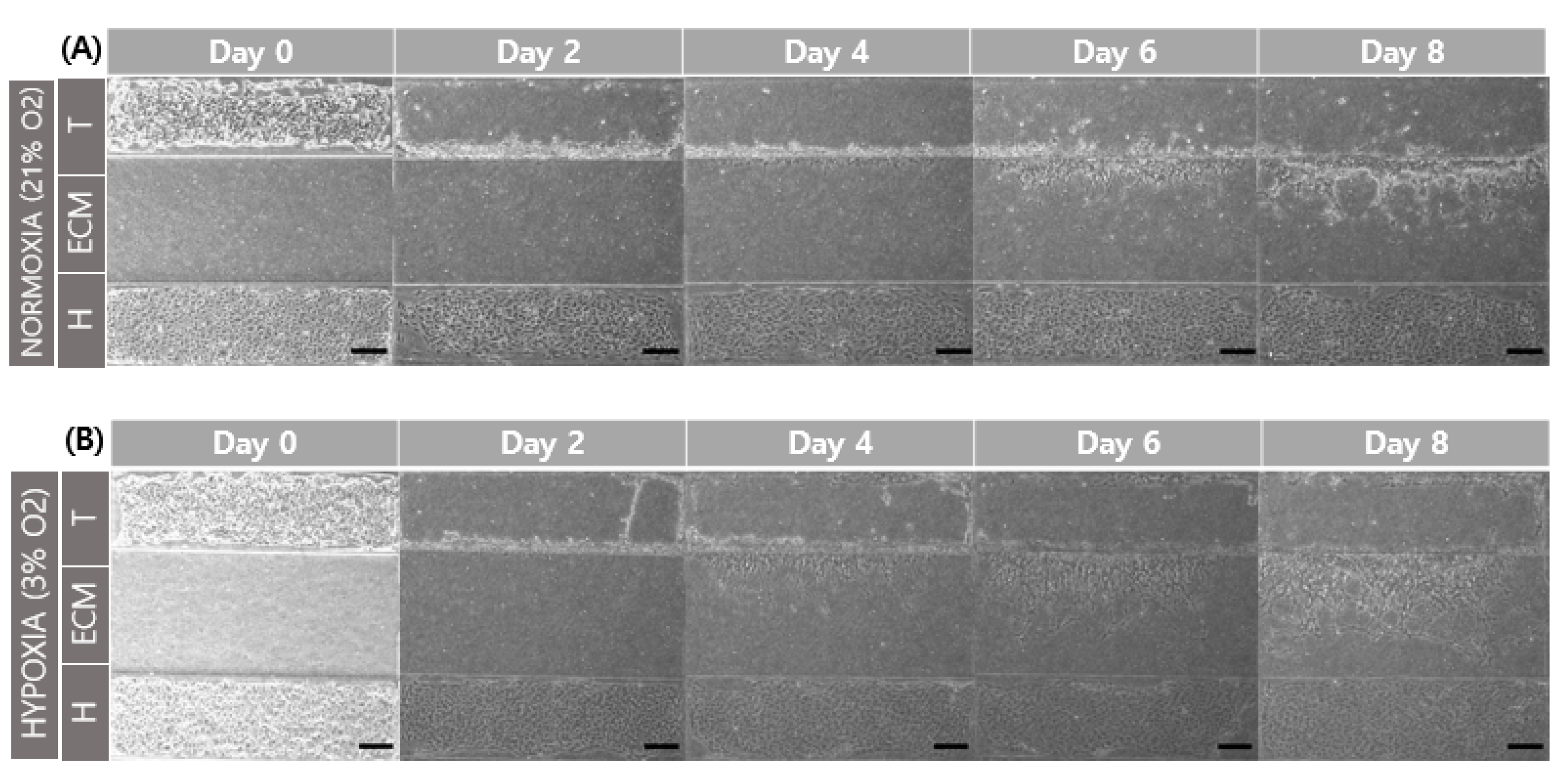
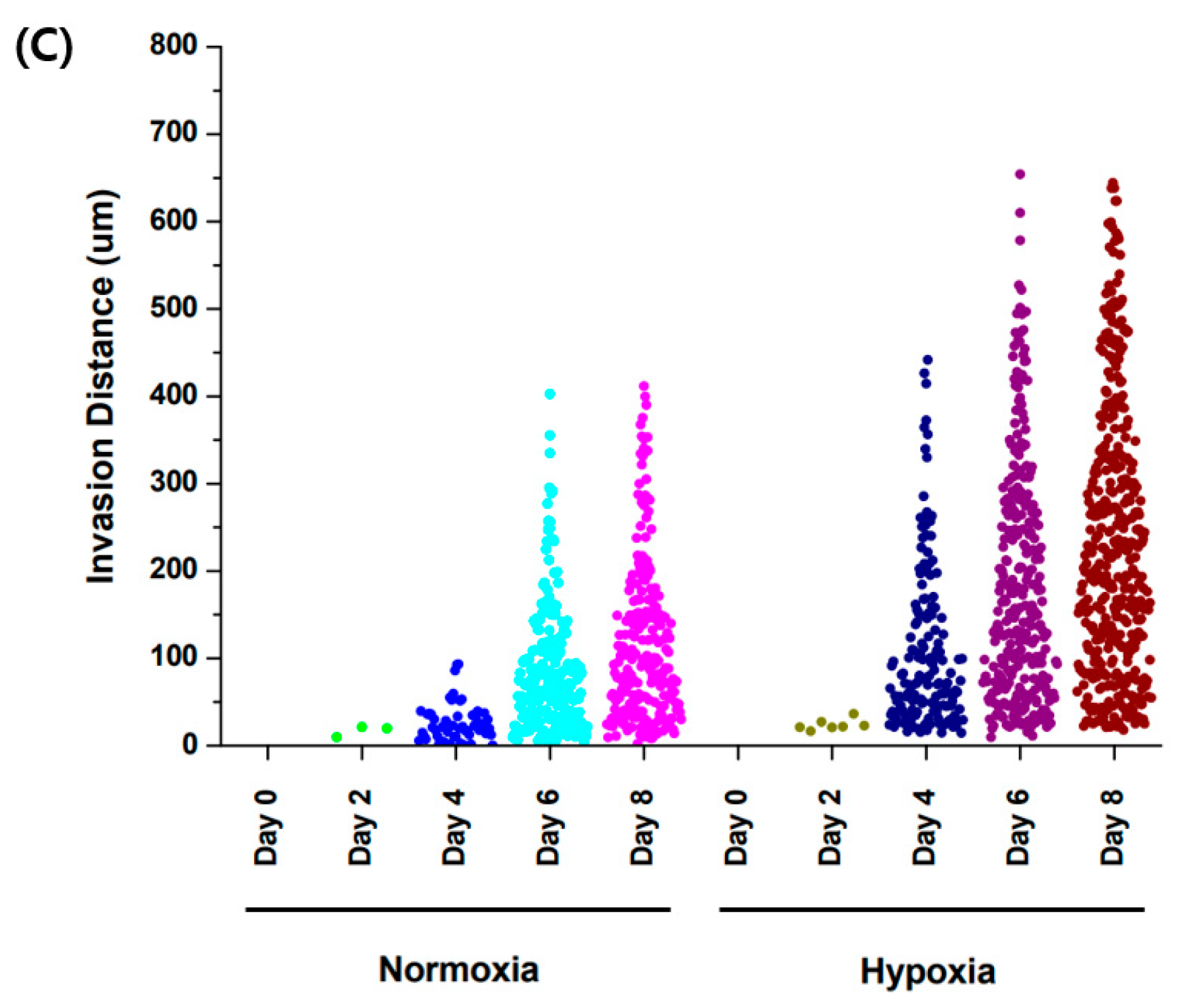
3.4. Comparison of Trophoblast Cell Migration with Different Oxygen Levels
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Craven, C.M.; Zhao, L.; Ward, K. Lateral Placental Growth Occurs by Trophoblast Cell Invasion of Decidual Veins. Placenta 2000, 21, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.F.; Serakides, R. Intrauterine Trophoblast Migration: A Comparative View of Humans and Rodents. Cell Adhes. Migr. 2016, 10, 88–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.W.; Wakeland, A.K.; Parast, M.M. Trophoblast Lineage Specification, Differentiation and Their Regula-tion by Oxygen Tension. J. Endocrinol. 2018, 236, R43–R56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppertz, B. Oxygenation of the Placenta and Its Role in Pre-Eclampsia. Pregnancy Hypertens. Int. J. Women’s Cardiovasc. Health 2014, 4, 244–245. [Google Scholar] [CrossRef]
- Al-Jameil, N.; Khan, F.A.; Khan, M.F.; Tabassum, H. A Brief Overview of Preeclampsia. J. Clin. Med. Res. 2014, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Steegers, E.A.P.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-Eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Pennington, K.A.; Schlitt, J.M.; Jackson, D.L.; Schulz, L.C.; Schust, D.J. Preeclampsia: Multiple Approaches for a Multifactorial Disease. Dis. Model Mech. 2012, 5, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Lee, H.J.; Yang, T.H.; Kim, G.J. Effects of Hypoxia Inducible Factors-1α on Autophagy and Invasion of Trophoblasts. Clin. Exp. Reprod. Med. 2012, 39, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, P.; Babu, G.S.; Sobti, R.C.; Gupta, S.K. HGF Regulate HTR-8/SVneo Trophoblastic Cells Migra-tion/Invasion under Hypoxic Conditions through Increased HIF-1α Expression via MAPK and PI3K Pathways. J. Cell Commun. Signal. 2019, 13, 503–521. [Google Scholar] [CrossRef]
- Goteri, G.; Lucarini, G.; Montik, N.; Zizzi, A.; Stramazzotti, D.; Fabris, G.; Tranquilli, A.L.; Ciavattini, A. Expres-sion of Vascular Endothelial Growth Factor (VEGF), Hypoxia Inducible Factor-1α (HIF-1α), and Microvessel Density in Endometrial Tissue in Women with Adenomyosis. Int. J. Gynecol. Pathol. 2009, 28, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Wan, R.; Mo, Y.; Chien, S.; Li, Y.; Li, Y.; Tollerud, D.J.; Zhang, Q. The Role of Hypoxia Inducible Factor-1α in the Increased MMP-2 and MMP-9 Production by Human Monocytes Exposed to Nickel Nanoparticles. Nanotoxicology 2011, 5, 568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Ma, Y.; Jiang, L.; Mu, Z.; Jiang, Z.; Chen, X.; Wang, Y.; Yang, G.Y.; Zhang, Z. Hypoxia Response Ele-ment-Regulated MMP-9 Promotes Neurological Recovery via Glial Scar Degradation and Angiogenesis in De-layed Stroke. Mol. Ther. 2017, 25, 1448–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Li, P.; Wang, Y.; Yan, H. Hypoxia-Induced Expression of CXCR4 Favors Trophoblast Cell Migration and Invasion via the Activation of HIF-1α. Int. J. Mol. Med. 2018, 42, 1508–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.-Y.; Pang, Z.-J.; Yu, Y. Regulation of Trophoblast Invasion: The Role of Matrix Metalloproteinases. Rev. Obstet. Gynecol. 2012, 5, e137. [Google Scholar] [CrossRef] [PubMed]
- Pollheimer, J.; Fock, V.; Knöfler, M. Review: The ADAM Metalloproteinases—Novel Regulators of Trophoblast Invasion? Placenta 2014, 35, S57–S63. [Google Scholar] [CrossRef]
- Staun-Ram, E.; Goldman, S.; Gabarin, D.; Shalev, E. Expression and Importance of Matrix Metalloproteinase 2 and 9 (MMP-2 and -9) in Human Trophoblast Invasion. Reprod. Biol. Endocrinol. 2004, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Abbas, Y.; Turco, M.Y.; Burton, G.J.; Moffett, A. Investigation of Human Trophoblast Invasion in Vitro. Hum. Re-Prod Update 2020, 26, 501–513. [Google Scholar] [CrossRef]
- Pu, Y.; Gingrich, J.; Veiga-Lopez, A. A 3-Dimensional Microfluidic Platform for Modeling Human Extravillous Trophoblast Invasion and Toxicological Screening. Lab Chip 2021, 21, 546–557. [Google Scholar] [CrossRef]
- Park, J.Y.; Mani, S.; Clair, G.; Olson, H.M.; Paurus, V.L.; Ansong, C.K.; Blundell, C.; Young, R.; Kanter, J.; Gordon, S.; et al. A Microphysiological Model of Human Trophoblast Invasion during Implantation. Nat. Commun. 2022, 13, 1252. [Google Scholar] [CrossRef]
- Blundell, C.; Tess, E.R.; Schanzer, A.S.R.; Coutifaris, C.; Su, E.J.; Parry, S.; Huh, D. A Microphysiological Model of the Human Placental Barrier. Lab Chip 2016, 16, 3065–3073. [Google Scholar] [CrossRef]
- Lee, J.; Song, B.; Subbiah, R.; Chung, J.J.; Choi, U.H.; Park, K.; Kim, S.H.; Oh, S.J. Effect of Chain Flexibility on Cell Adhesion: Semi-Flexible Model-Based Analysis of Cell Adhesion to Hydrogels. Sci. Rep. 2019, 9, 2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambuto, S.G.; Clancy, K.B.H.; Harley, B.A.C. A Gelatin Hydrogel to Study Endometrial Angiogenesis and Trophoblast Invasion. Interface Focus 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambuto, S.G.; Rattila, S.; Dveksler, G.; Harley, B.A.C.; Harley, B.A.C. The Role of Pregnancy-Specific Glycopro-teins on Trophoblast Motility in Three-Dimensional Gelatin Hydrogels. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ibañez, R.I.R.; do Amaral, R.J.F.C.; Reis, R.L.; Marques, A.P.; Murphy, C.M.; O’brien, F.J. 3D-Printed Gelatin Meth-acrylate Scaffolds with Controlled Architecture and Stiffness Modulate the Fibroblast Phenotype towards Dermal Regeneration. Polymers 2021, 13, 2510. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, G.; Jeon, T.-J.; Kim, S.M. Trophoblast Migration with Different Oxygen Levels in a Gel-Patterned Microfluidic System. Micromachines 2022, 13, 2216. https://doi.org/10.3390/mi13122216
Ko G, Jeon T-J, Kim SM. Trophoblast Migration with Different Oxygen Levels in a Gel-Patterned Microfluidic System. Micromachines. 2022; 13(12):2216. https://doi.org/10.3390/mi13122216
Chicago/Turabian StyleKo, Gun, Tae-Joon Jeon, and Sun Min Kim. 2022. "Trophoblast Migration with Different Oxygen Levels in a Gel-Patterned Microfluidic System" Micromachines 13, no. 12: 2216. https://doi.org/10.3390/mi13122216