

Numerical Simulations of the Effect of the Asymmetrical Bending of the Hindwings of a Hovering C. buqueti Bamboo Weevil with Respect to the Aerodynamic Characteristics
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Biological Scanning Operation
2.3. Definition of Euler Angles
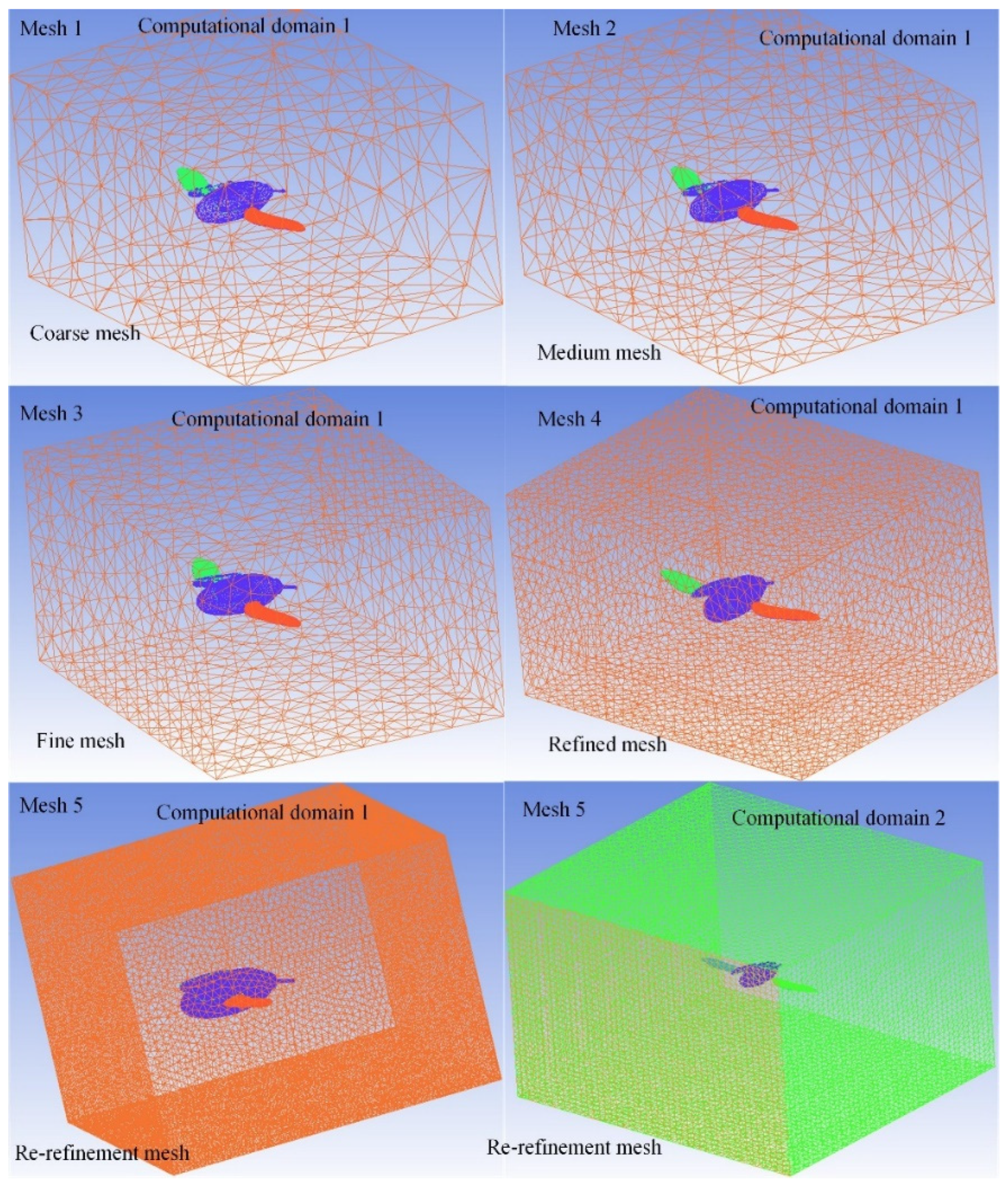
2.4. CFD Numerical Method
2.5. Finite Element Simulation
3. Results
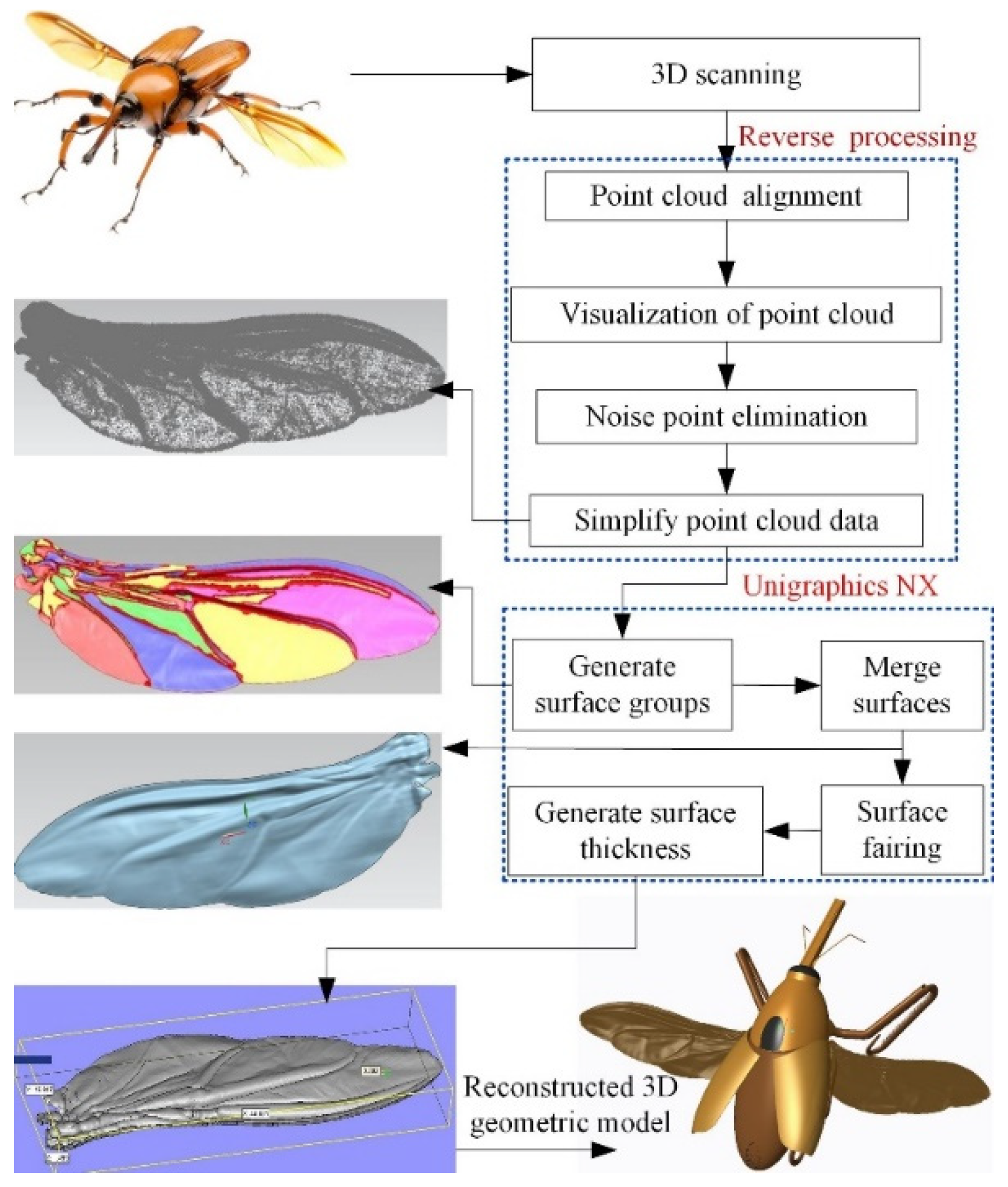
3.1. Reverse Reconstruction
3.2. Flapping Kinematics of the Hindwings
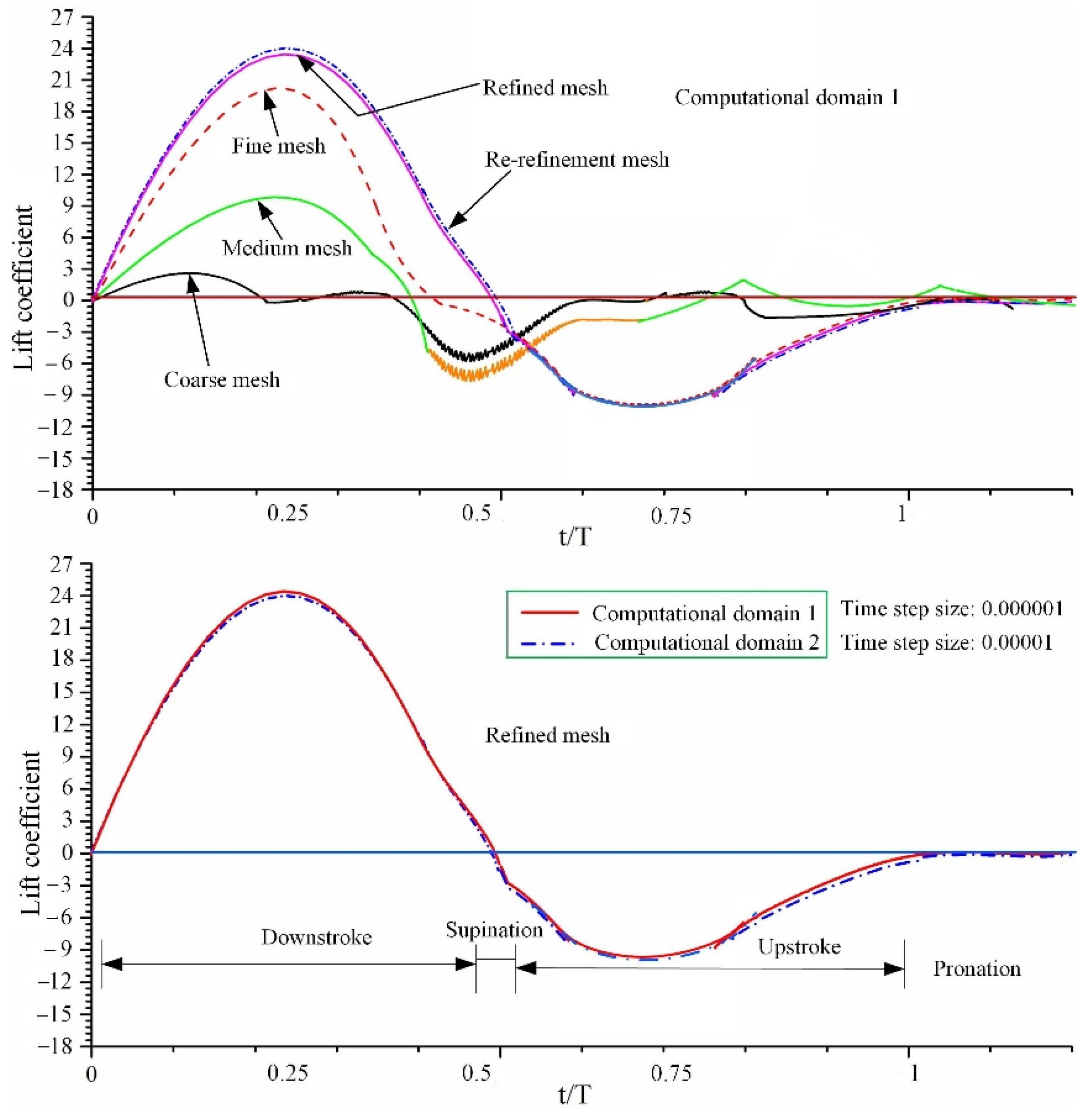
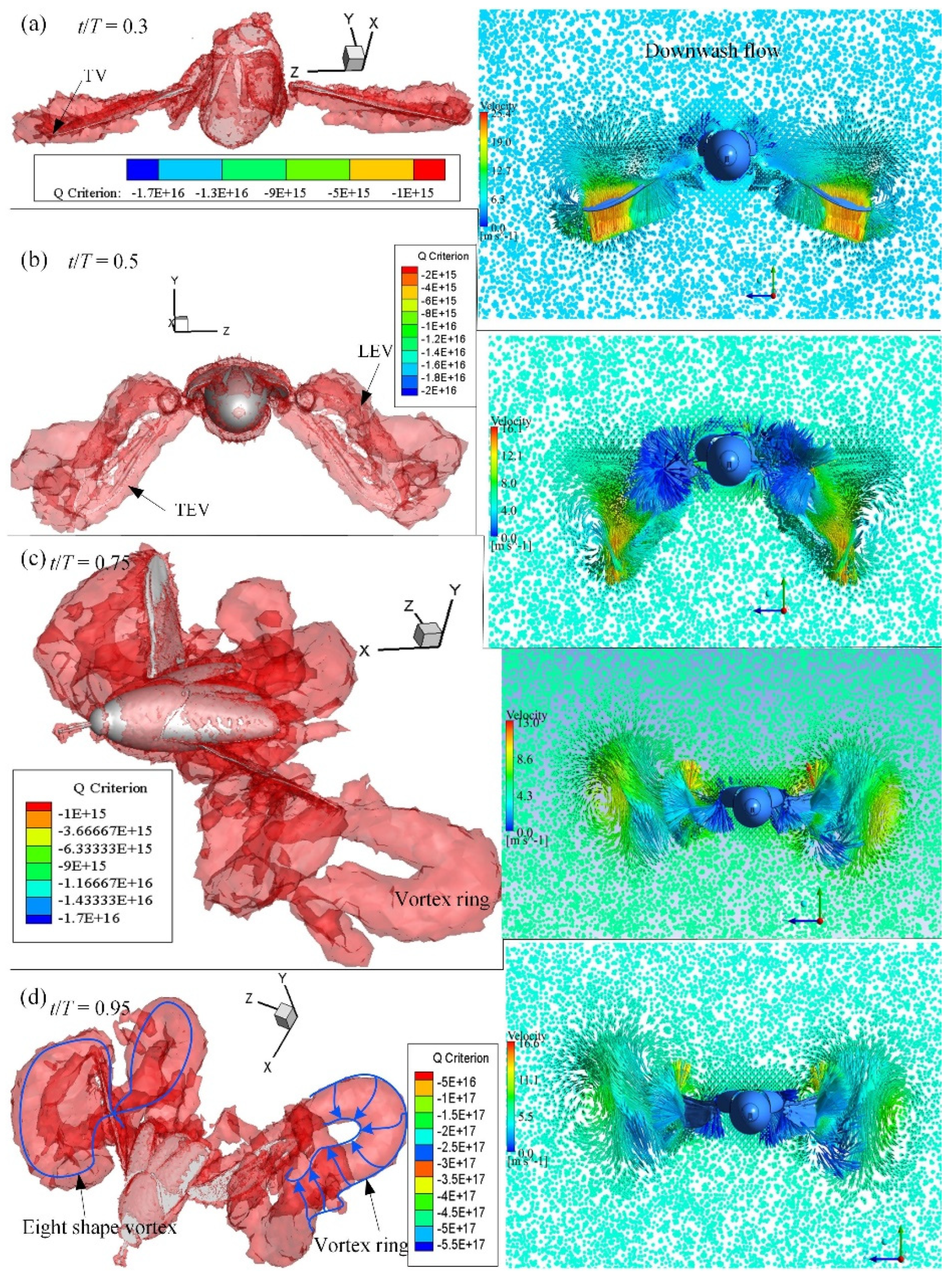
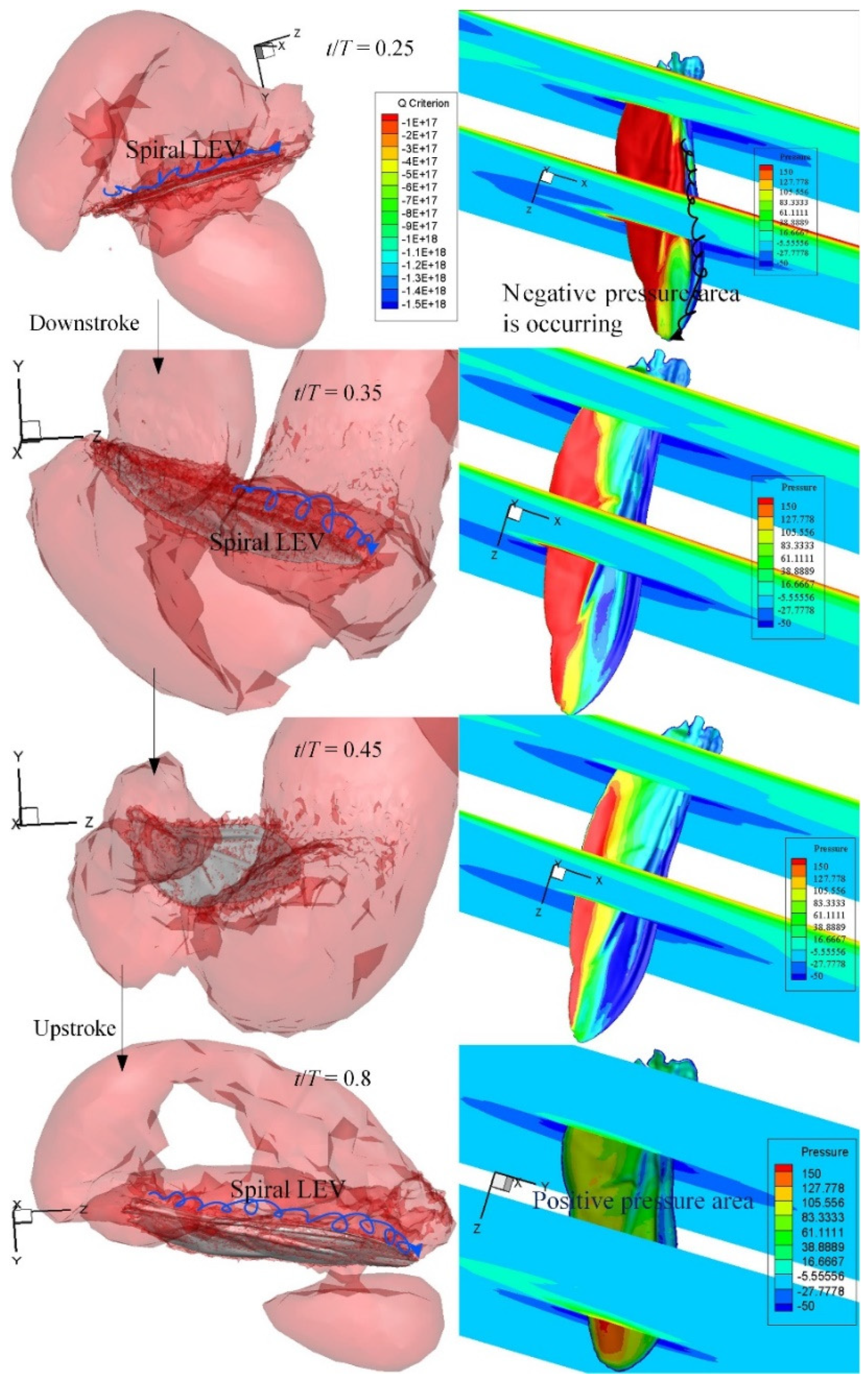
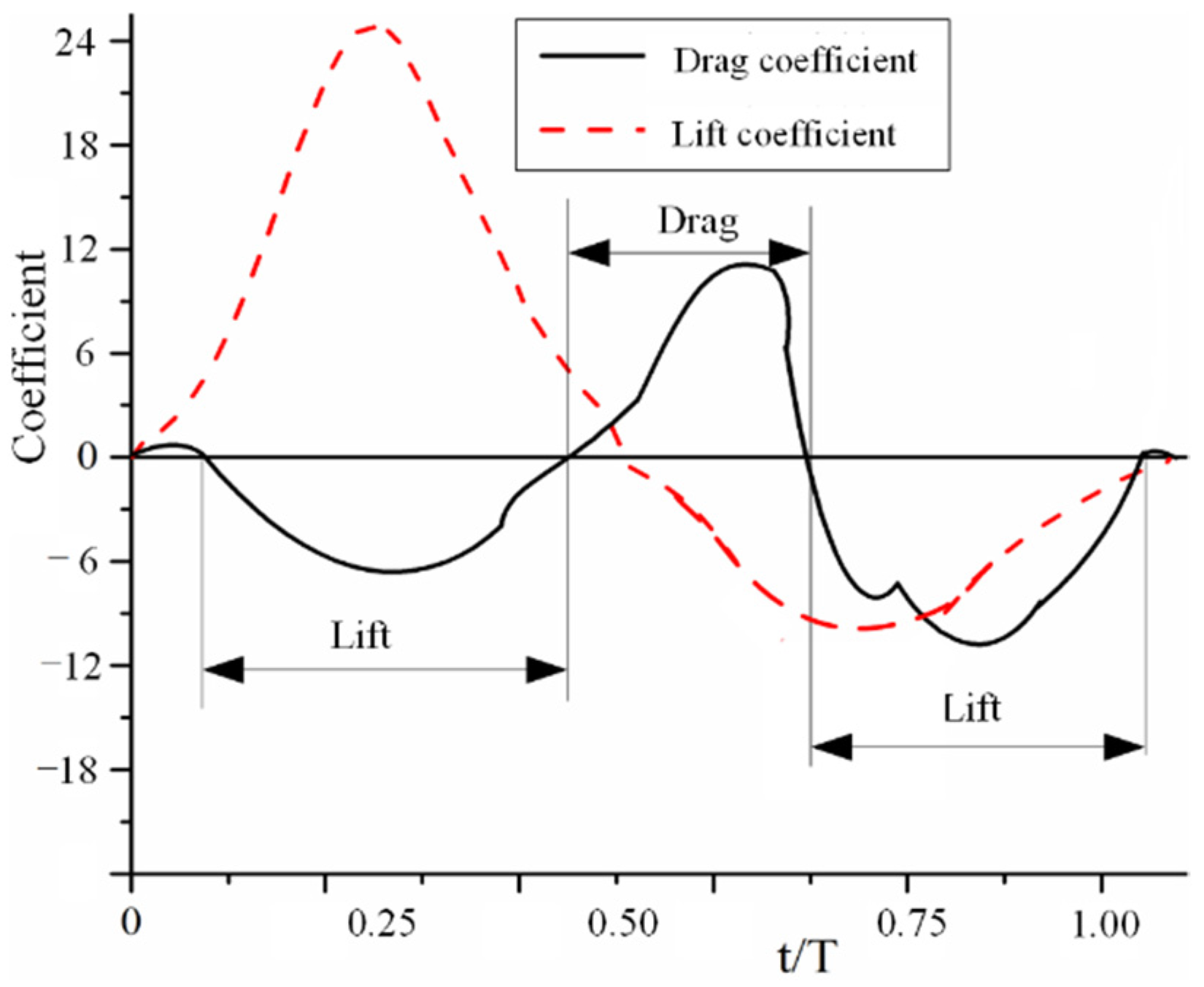
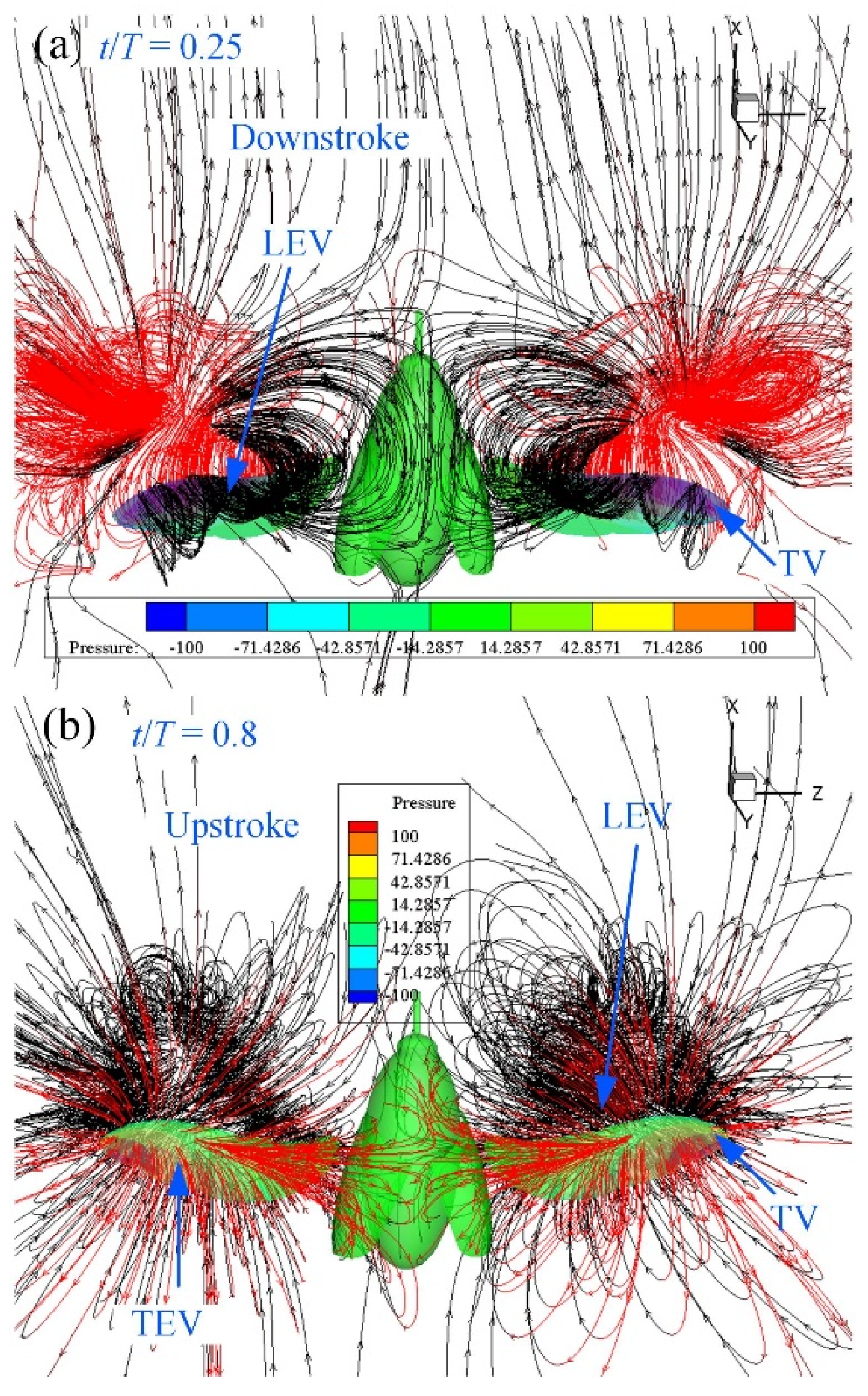
3.3. CFD Simulation Results
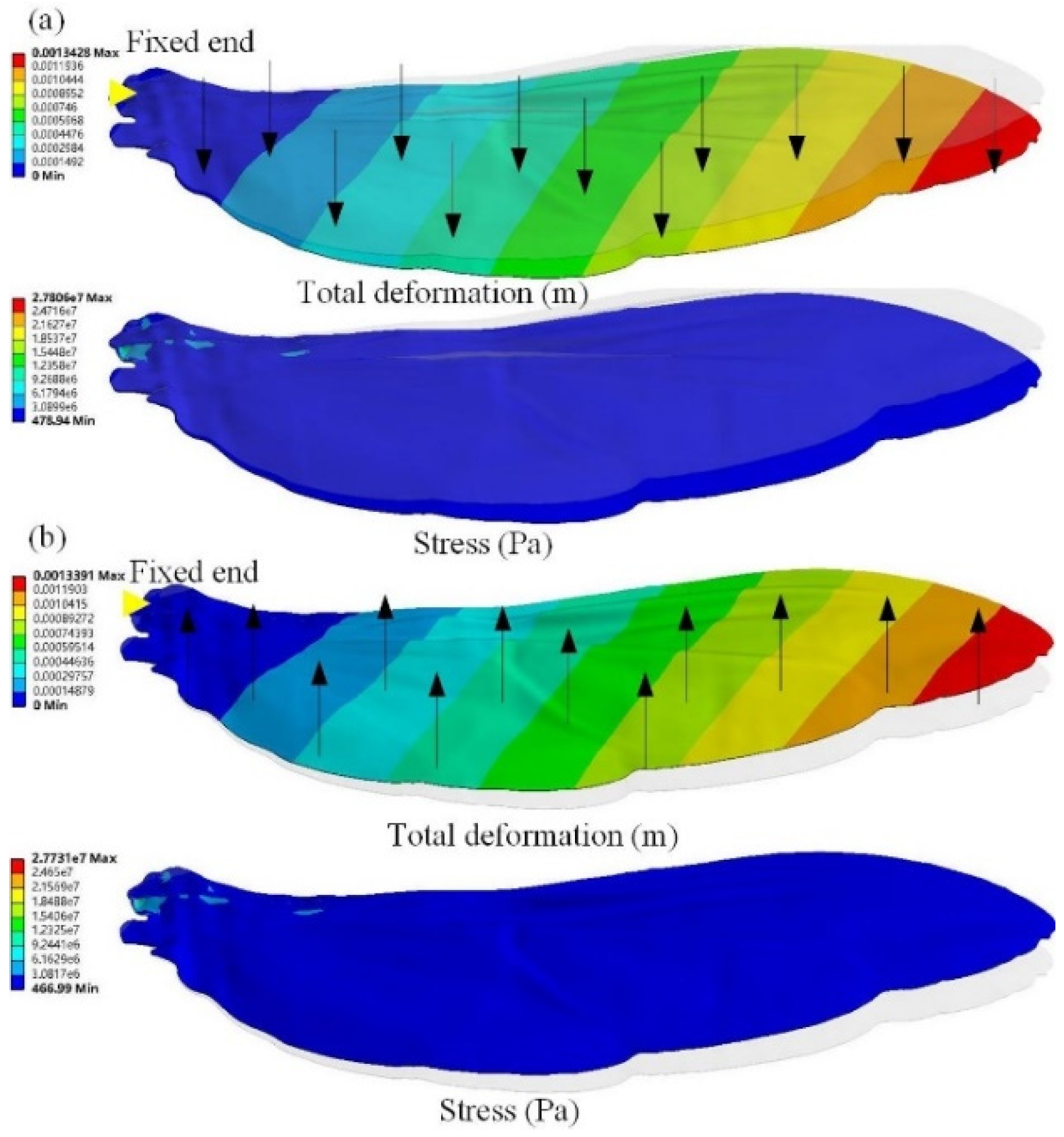
3.4. Asymmetrical Bending
4. Discussion
4.1. Wing Kinematics Analysis
4.2. Flapping Aerodynamic Characteristics
4.3. Asymmetrical Bending Analysis
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, J.; Walker, S.M.; Bomphrey, R.J.; Taylor, G.K.; Thomas, A.L.R. Details of Insect Wing Design and Deformation Enhance Aerodynamic Function and Flight Efficiency. Science 2009, 325, 1549–1552. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, W. Numerical analysis of the three-dimensional aerodynamics of a hovering rufous hummingbird (Selasphorus rufus). Acta Mech. Sin. 2015, 31, 931–943. [Google Scholar] [CrossRef]
- Xiang, J.; Du, J.; Li, D.; Liu, K. Aerodynamic Performance of the Locust Wing in Gliding Mode at Low Reynolds Number. J. Bionic Eng. 2016, 13, 249–260. [Google Scholar] [CrossRef]
- Wan, H.; Dong, H.; Gai, K. Computational investigation of cicada aerodynamics in forward flight. J. R. Soc. Interface 2015, 12, 20141116. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Song, B.; Yang, W.; Xue, D.; Lang, X. Unsteady characteristic research on aerodynamic interaction of slotted wingtip in flapping kinematics. Chin. J. Aeronaut. 2022, 35, 82–101. [Google Scholar] [CrossRef]
- Phan, H.V.; Park, H.C. Mechanisms of collision recovery in flying beetles and flapping-wing robots. Science 2020, 370, 1214–1219. [Google Scholar] [CrossRef]
- Phan, H.V.; Au, T.K.L.; Park, H.C. Clap-and-fling mechanism in a hovering insect-like two-winged flapping-wing micro air vehicle. R. Soc. Open Sci. 2016, 3, 160746. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Dong, H. Wing kinematics measurement and aerodynamics of a dragonfly in turning flight. Bioinspir. Biomim. 2017, 12, 026001. [Google Scholar] [CrossRef]
- Karásek, M.; Muijres, F.T.; De Wagter, C.; Remes, B.D.W.; de Croon, G.C.H.E. A tailless aerial robotic flapper reveals that flies use torque coupling in rapid banked turns. Science 2018, 361, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Debiasi, M.; Nguyen, Q.V.; Chan, W.L. Bioinspired Low-Noise Wing Design for a Two-Winged Flapping-Wing Micro Air Vehicle. AIAA J. 2018, 56, 4697–4705. [Google Scholar] [CrossRef]
- Truong, N.T.; Phan, H.V.; Park, H.C. Design and demonstration of a bio-inspired flapping-wing-assisted jumping robot. Bioinspir. Biomim. 2019, 14, 036010. [Google Scholar] [CrossRef] [PubMed]
- Au, L.T.K.; Park, H.C.; Lee, S.T.; Hong, S.K. Clap-and-Fling Mechanism in Non-Zero Inflow of a Tailless Two-Winged Flapping-Wing Micro Air Vehicle. Aerospace 2022, 9, 108. [Google Scholar] [CrossRef]
- Badrya, C.; Sridharan, A.; Baeder, J.D.; Kroninger, C.M. Multi-Fidelity Coupled Trim Analysis of a Flapping-Wing Micro Air Vehicle Flight. J. Aircr. 2017, 54, 1614–1630. [Google Scholar] [CrossRef]
- Xiang, J.; Liu, K.; Li, D.; Du, J. Effects of micro-structure on aerodynamics of Coccinella septempunctata elytra (ladybird) in forward flight as assessed via electron microscopy. Micron 2017, 102, 21–34. [Google Scholar] [CrossRef]
- Johansson, L.C.; Engel, S.; Baird, E.; Dacke, M.; Muijres, F.T.; Hedenström, A. Elytra boost lift, but reduce aerodynamic efficiency in flying beetles. J. R. Soc. Interface 2012, 9, 2745–2748. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chao, L.; Bhushan, B.; Wu, W.; Tong, J. Effect of microtrichia on the interlocking mechanism in the Asian ladybeetle, Harmonia axyridis (Coleoptera: Coccinellidae). Beilstein J. Nanotechnol. 2018, 9, 812–823. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Guo, C.; Ma, Y.; Zheng, Y. Design of Bionic Foldable Wing Mimicking the Hind Wings of the C. Buqueti Bamboo Weevil. J. Mech. Des. 2021, 143, 1–15. [Google Scholar] [CrossRef]
- Lee, B.; Park, H.; Kim, S.T. Three-dimensional wing behaviors of a rhinoceros beetle during takeoff flights. J. Mech. Sci. Technol. 2015, 29, 5281–5288. [Google Scholar] [CrossRef]
- Le, T.Q.; Truong, T.V.; Tran, H.T.; Park, S.H.; Ko, J.H.; Park, H.C.; Yoon, K.J.; Byun, D. Two-and Three-Dimensional Simulations of Beetle Hind Wing Flapping during Free Forward Flight. J. Bionic Eng. 2013, 10, 316–328. [Google Scholar] [CrossRef]
- Le, T.Q.; Truong, T.V.; Tran, H.T.; Park, S.H.; Ko, J.H.; Park, H.C.; Byun, D. How Could Beetle’s Elytra Support Their Own Weight during Forward Flight? J. Bionic Eng. 2014, 11, 529–540. [Google Scholar] [CrossRef]
- Li, X.; Guo, C. Wing-kinematics measurement and flight modelling of the bamboo weevil C. buqueti. IET Nanobiotechnol. 2020, 14, 53–58. [Google Scholar] [CrossRef]
- Le, T.Q.; Byun, D.; Ko, J.H.; Park, H.C.; Kim, M. Numerical investigation of the aerodynamic characteristics of a hovering Coleopteran insect. J. Theor. Biol. 2010, 266, 485–495. [Google Scholar] [CrossRef]
- Truong, T.V.; Le, T.Q.; Tran, H.T.; Park, H.C.; Yoon, K.J.; Byun, D. Flow visualization of rhinoceros beetle (Trypoxylus dichotomus) in free flight. J. Bionic Eng. 2015, 9, 304–314. [Google Scholar] [CrossRef]
- Sitorus, P.E.; Park, H.C.; Byun, D.; Goo, N.S.; Han, C.H. The role of elytra in beetle flight: I. generation of quasi-static aerodynamic forces. J. Bionic Eng. 2010, 7, 354–363. [Google Scholar] [CrossRef]
- Prasath, N.G.; Rangaraj, V.; Mathaiyan, V.; Kishore, G.; Mohanraj, M.; Kumar, V.S. Numerical Simulation of Biology-Inspired Beetle Wings at Various Flying Conditions. In Proceedings of the 12th International Energy Conversion Engineering Conference, Cleveland, OH, USA, 28–30 July 2014. [Google Scholar]
- Song, Z.; Tong, J.; Yan, Y.; Tian, L.; Sun, J. The rigidizable behavior of the deployable hindwings of the asian ladybeetle during flight. J. Mater. Sci. 2021, 56, 5670–5683. [Google Scholar] [CrossRef]
- Liu, C.; Li, P.; Song, F.; Stamhuis, E.J.; Sun, J. Design optimization and wind tunnel investigation of a flapping system based on the flapping wing trajectories of a beetle’s hindwings. Comput. Biol. Med. 2022, 140, 105085. [Google Scholar] [CrossRef]
- Lehmann, F.O.; Gorb, S.; Nasir, N.; Schuützner, P. Elastic deformation and energy loss of flapping fly wings. J. Exp. Biol. 2011, 214, 2949–2961. [Google Scholar] [CrossRef] [Green Version]
- Steppan, S.J. Flexural stiffness patterns of butterfly wings (Papilionoidea). J. Res. Lepid. 2000, 35, 61–77. [Google Scholar] [CrossRef]
- Combes, S.A. Flexural stiffness in insect wings II. Spatial distribution and dynamic wing bending. J. Exp. Biol. 2003, 206, 2989–2997. [Google Scholar] [CrossRef] [Green Version]
- Wootton, R.J.; Evans, K.E.; Herbert, R.; Smith, C.W. The hind wing of the desert locust (Schistocerca gregaria Forskal). I. Functional morphology and mode of operation. J. Exp. Biol. 2000, 203, 2921–2931. [Google Scholar] [CrossRef]
- Truong, V.T.; Le, Q.T.; Byun, D.Y.; Park, H.C.; Kim, M.J. Flexible Wing Kinematics of a Free-Flying Beetle (Rhinoceros Beetle Trypoxylus dichotomus). J. Bionic Eng. 2012, 9, 177–184. [Google Scholar] [CrossRef]
- Ha, N.S.; Truong, Q.T.; Goo, N.S.; Park, H.C. Biomechanical Properties of Insect Wings: The Stress Stiffening Effects on the Asymmetric Bending of the Allomyrina dichotoma Beetle’s Hind Wing. PLoS ONE 2013, 8, e80689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.B.; Luo, H.; Song, J.; Lu, X.Y. Force production and asymmetric deformation of a flexible flapping wing in forward flight. J. Fluids Struct. 2013, 36, 149–161. [Google Scholar] [CrossRef]
- Nian, P.; Song, B.; Xuan, J.; Zhou, W.; Xue, D. Study on flexible flapping wings with three dimensional asymmetric passive deformation in a flapping cycle. Aerosp. Sci. Technol. 2020, 104, 105944. [Google Scholar] [CrossRef]
- Li, X.; Guo, C. Structural characteristics analysis of the hind wings in a bamboo weevil (Cyrtotrachelus buqueti). IET Nanobiotechnol. 2019, 13, 850–856. [Google Scholar] [CrossRef]
- Li, X.; Zheng, Y. Functional characteristics of the rigid elytra in a bamboo weevil beetle Cyrtotrachelus buqueti. IET Nanobiotechnol. 2022, 16, 273–283. [Google Scholar] [CrossRef]
- Sun, J.; Du, R.; Liu, X.; Bechkoum, K.; Tong, J.; Chen, D. A simulation of the flight characteristics of the deployable hindwings of beetle. J. Bionic Eng. 2017, 14, 296–306. [Google Scholar] [CrossRef]
- Li, X.; Guo, C. Microstructure and material properties of hind wings of a bamboo weevil Cyrtotrachelus buqueti (Coleoptera: Curculionidae). Microsc. Res. Tech. 2019, 82, 1102–1113. [Google Scholar] [CrossRef]
- Chen, Y.H.; Skote, M.; Zhao, Y.; Huang, W.M. Dragonfly (Sympetrum flaveolum) flight: Kinematic measurement and modelling. J. Fluids Struct. 2013, 40, 115–126. [Google Scholar] [CrossRef]
- Srygley, R.B.; Thomas, A.L. Unconventional lift-generating mechanisms in free-flying butterflies. Nature 2002, 420, 660–664. [Google Scholar] [CrossRef]
- Hasan, J.; Roy, A.; Chatterjee, K.; Prasad, K.D.V.Y. Mimicking Insect Wings: The Roadmap to Bio-inspiration. ACS Biomater. Sci. Eng. 2019, 5, 3139–3160. [Google Scholar] [CrossRef]
- Dickinson, M.H.; Lehmann, F.O.; Sane, S.P. Wing rotation and the aerodynamic basis of insect flight. Science 1999, 284, 1954–1960. [Google Scholar] [CrossRef]
- Sun, M.; Wu, J.H. Aerodynamic force generation and power requirements in forward flight in a fruit fly with modeled wing motion. J. Exp. Biol. 2003, 206, 3065–3083. [Google Scholar] [CrossRef] [Green Version]
- Muijres, F.T.; Elzinga, M.J.; Melis, J.M.; Dickinson, M.H. Flies evade looming targets by executing rapid visually directed banked turns. Science 2014, 344, 172–177. [Google Scholar] [CrossRef]
- Sane, S.P.; Dickinson, M.H. The control of flight force by a flapping wing: Lift and drag production. J. Exp. Biol. 2001, 204, 2607–2626. [Google Scholar] [CrossRef]
- Birch, J.M.; Dickinson, M.H. Spanwise flow and the attachment of the leading-edge vortex on insect wings. Nature 2001, 412, 729–733. [Google Scholar] [CrossRef]
- Li, X.; Guo, C.; Li, L. Functional morphology and structural characteristics of the hind wings of the bamboo weevil Cyrtotrachelus buqueti (Coleoptera, Curculionidae). Anim. Cells Syst. 2019, 23, 143–153. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X. Numerical Simulations of the Effect of the Asymmetrical Bending of the Hindwings of a Hovering C. buqueti Bamboo Weevil with Respect to the Aerodynamic Characteristics. Micromachines 2022, 13, 1995. https://doi.org/10.3390/mi13111995
Li X. Numerical Simulations of the Effect of the Asymmetrical Bending of the Hindwings of a Hovering C. buqueti Bamboo Weevil with Respect to the Aerodynamic Characteristics. Micromachines. 2022; 13(11):1995. https://doi.org/10.3390/mi13111995
Chicago/Turabian StyleLi, Xin. 2022. "Numerical Simulations of the Effect of the Asymmetrical Bending of the Hindwings of a Hovering C. buqueti Bamboo Weevil with Respect to the Aerodynamic Characteristics" Micromachines 13, no. 11: 1995. https://doi.org/10.3390/mi13111995