
_Chang.jpg)
Quantitative Investigation of the Link between Actin Cytoskeleton Dynamics and Cellular Behavior

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Electric Stimulation
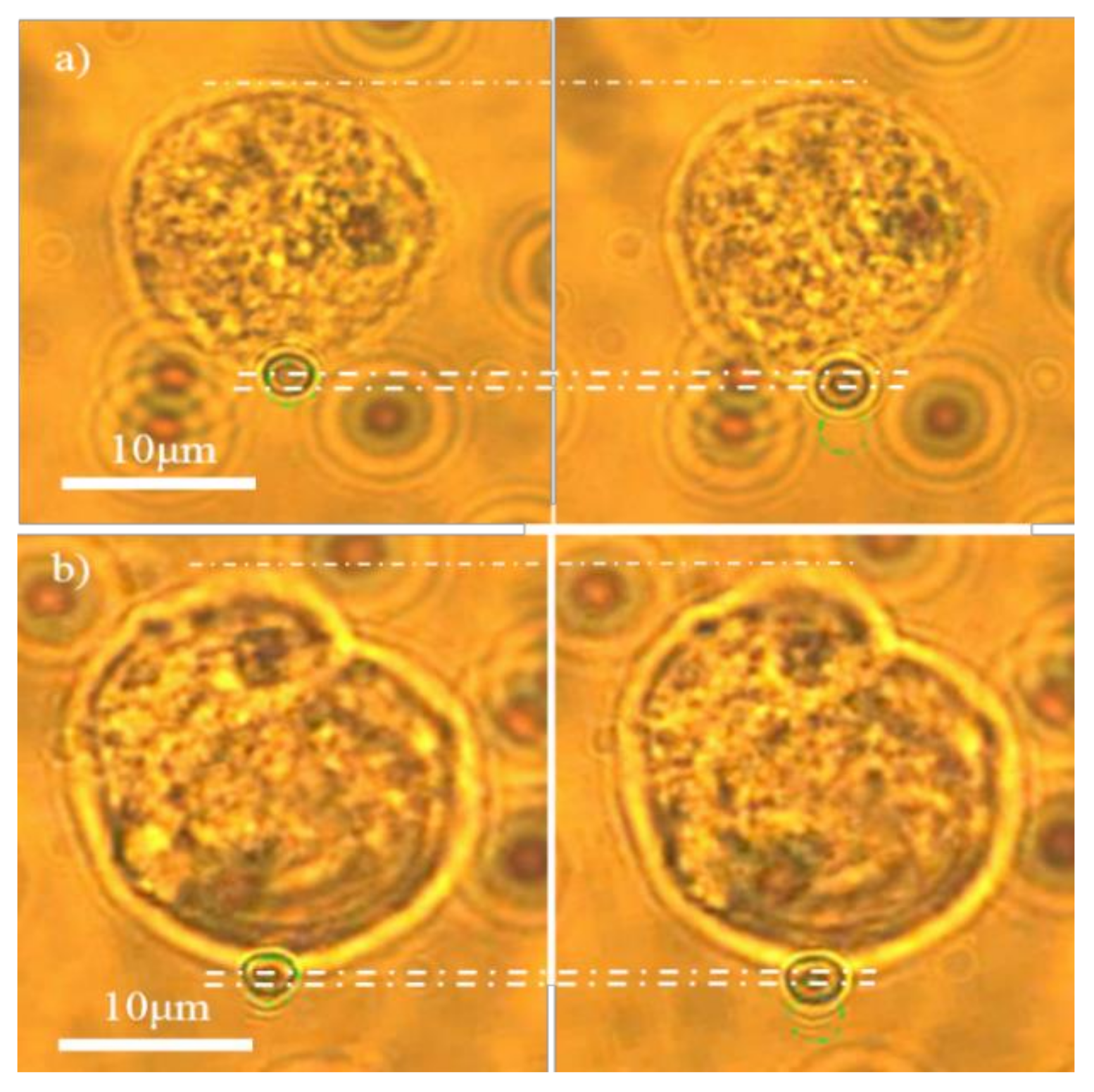
2.3. Optical Stretching
2.4. Actin Cytoskeleton Microstructural Model
2.5. RNA Isolation and RT PCR Analysis
3. Experiments and Results
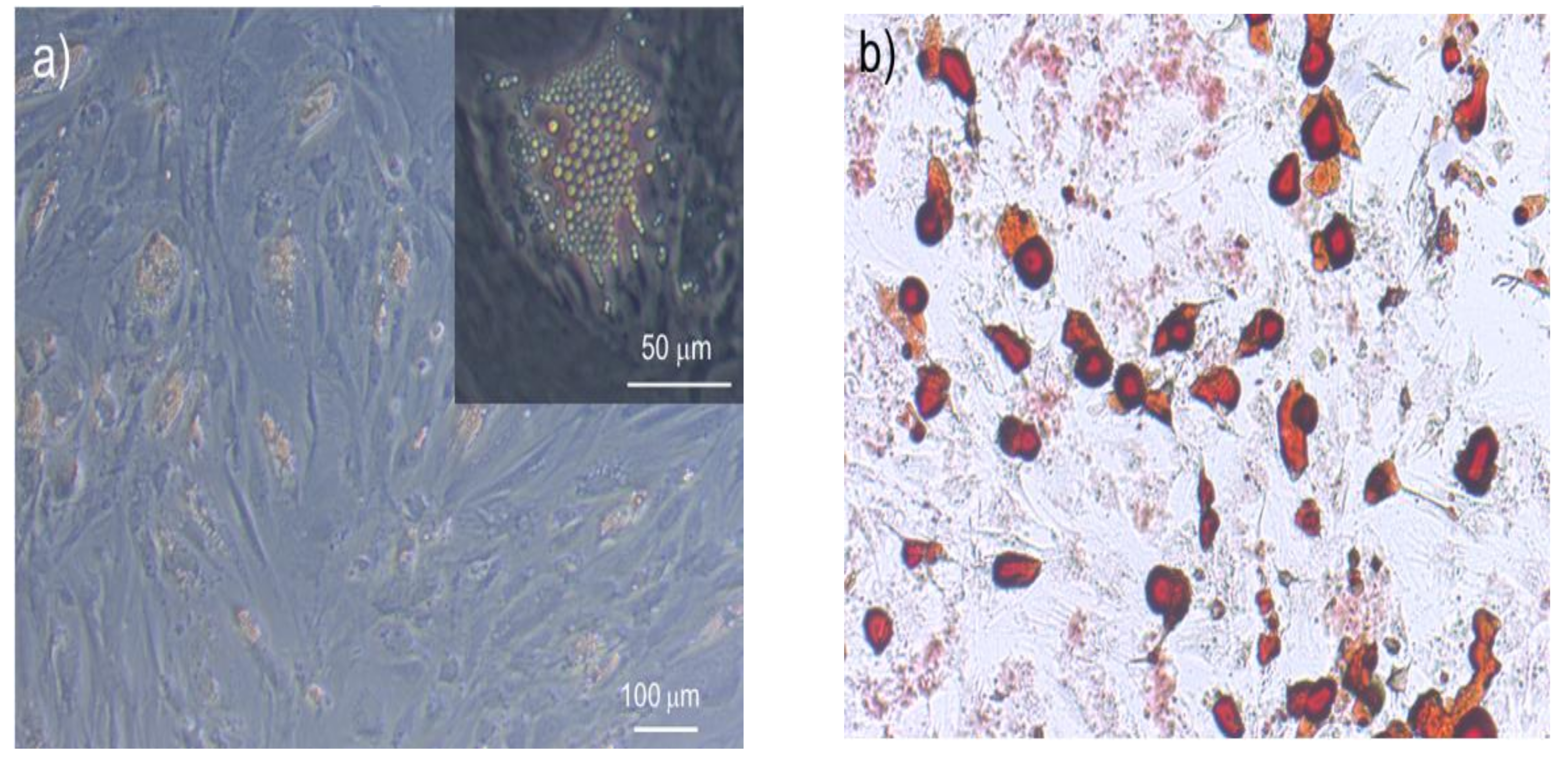
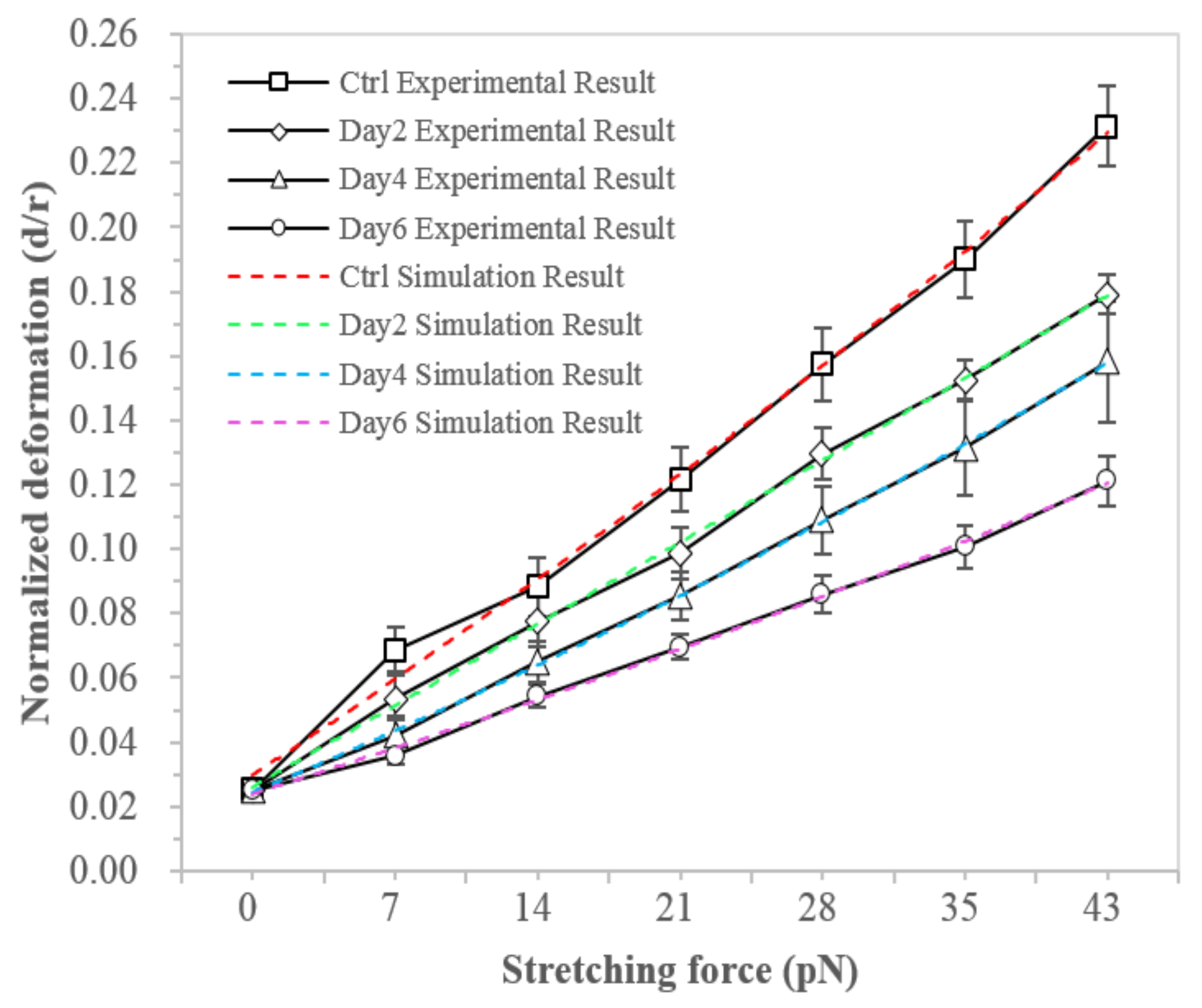
3.1. hMSCs Stretching Experiment
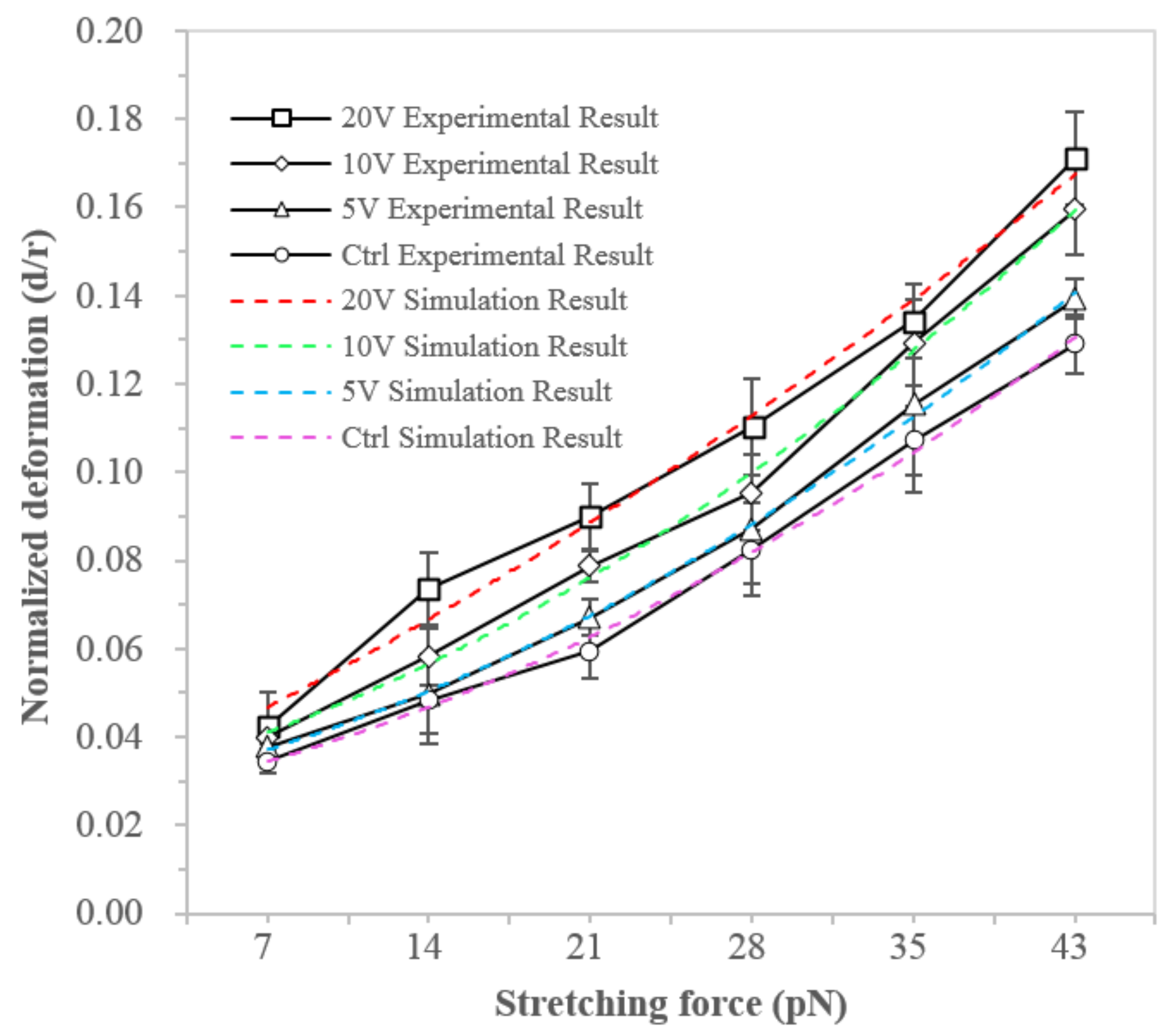
3.2. NB4 Cells Stretching Experiment
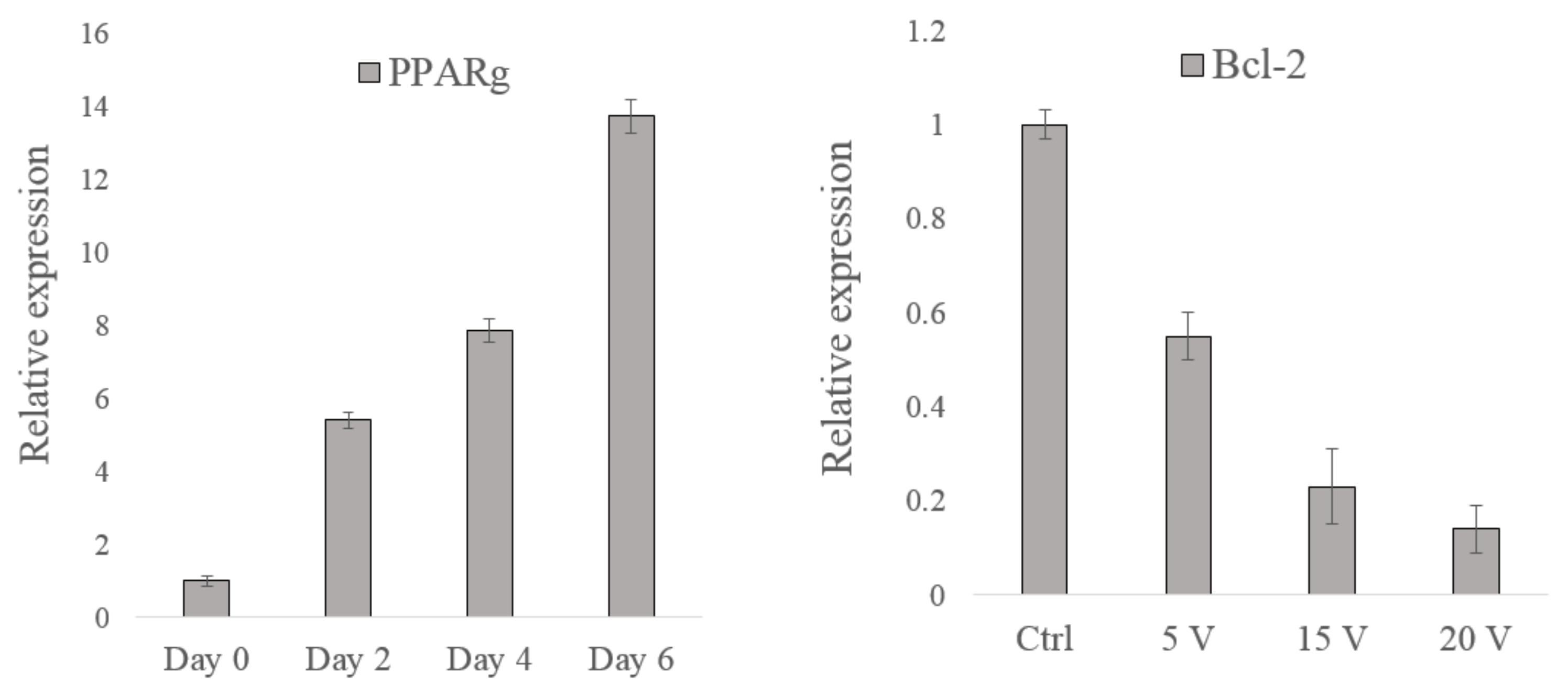
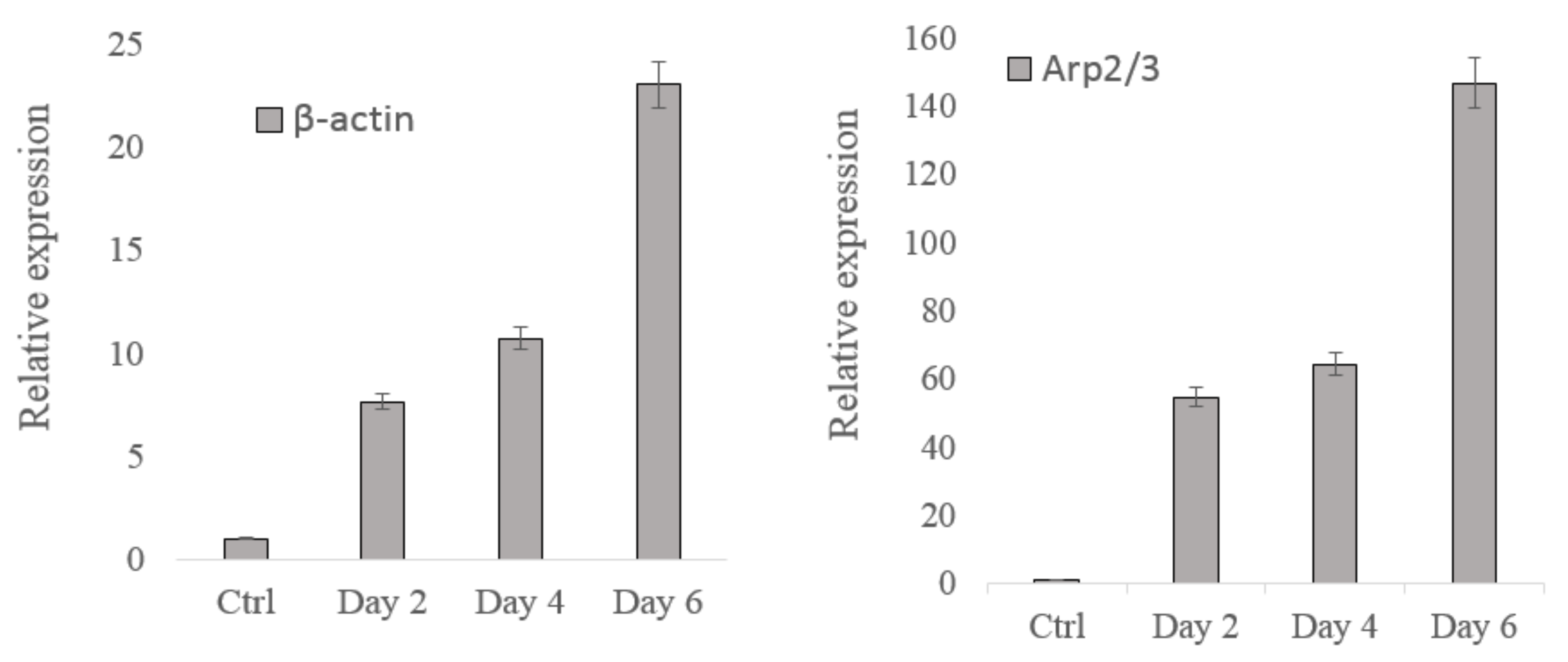
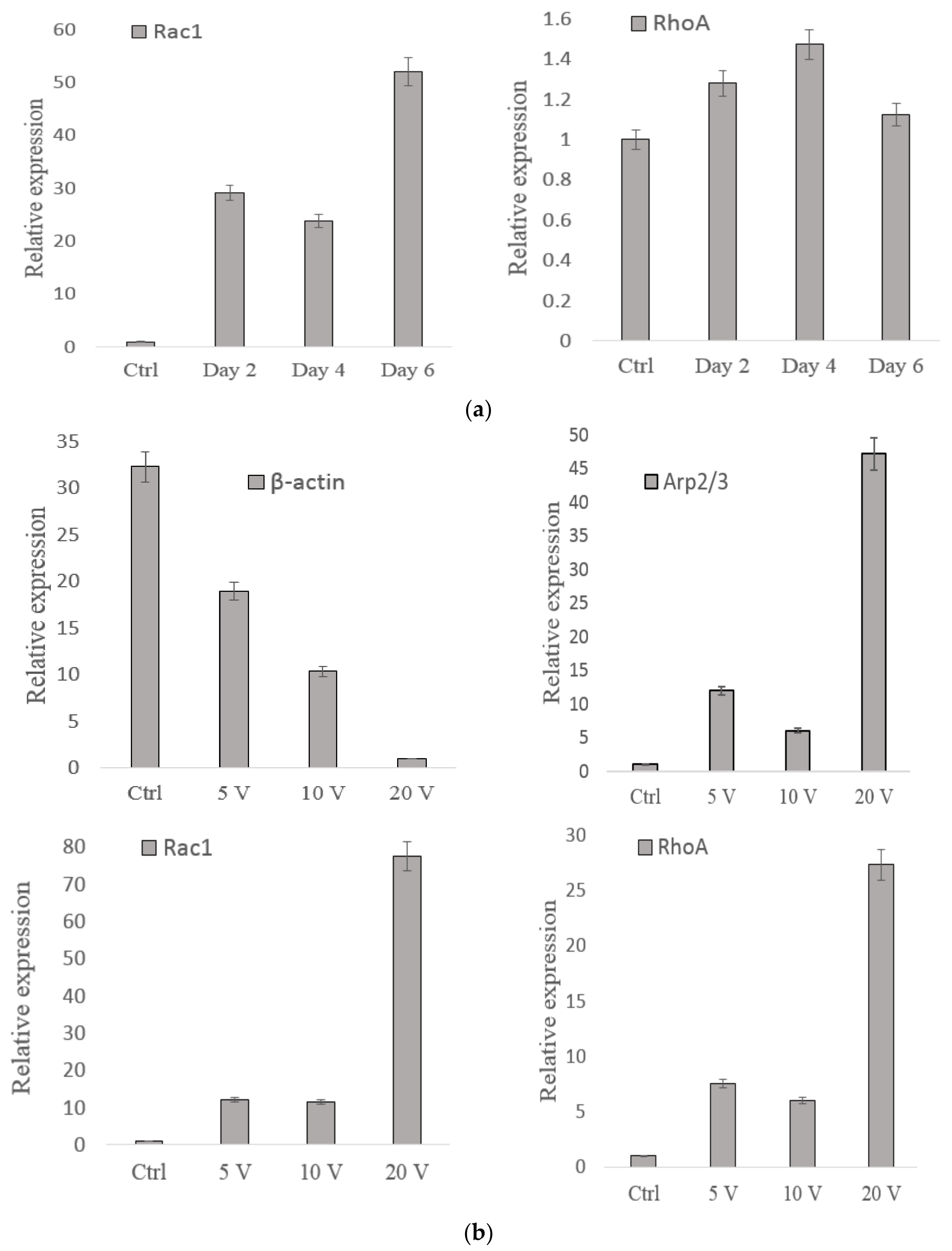
3.3. Gene Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, R.; Lee, D.M.; Jimah, J.R.; Gerassimov, N.; Yang, C.; Kim, S.; Luvsanjav, D.; Winkelman, J.; Mettlen, M.; Abrams, M.E.; et al. Dynamin regulates the dynamics and mechanical strength of the actin cytoskeleton as a multifilament actin-bundling protein. Nat. Cell Biol. 2022, 22, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Dogterom, M.; Koenderink, G.H. Actin–microtubule crosstalk in cell biology. Nat. Rev. Mol. Cell Biol. 2019, 20, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Gardel, M.L.; Schwarz, U.S. The actin cytoskeleton as an active adaptive material. Annu. Rev. Condens. Matter Phys. 2020, 11, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ménasché, G.; Longé, C.; Bratti, M.; Blank, U. Cytoskeletal transport, reorganization, and fusion regulation in mast cell-stimulus secretion coupling. Front. Cell Dev. Biol. 2021, 9, 652077. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, J.; Yao, Y. Extracellular nanofiber-orchestrated cytoskeletal reorganization and mediated directional migration of cancer cells. Nanoscale 2020, 12, 3183–3193. [Google Scholar] [CrossRef]
- Uetaki, M.; Onishi, N.; Oki, Y.; Shimizu, T.; Sugihara, E.; Sampetrean, O.; Watanabe, T.; Yanagi, H.; Suda, K.; Fujii, H.; et al. Regulatory roles of fibronectin and integrin α5 in reorganization of the actin cytoskeleton and completion of adipogenesis. Mol. Biol. Cell 2022, 33, mbc-E21. [Google Scholar] [CrossRef]
- Fan, Y.L.; Zhao, H.C.; Li, B.; Zhao, Z.L.; Feng, X.Q. Mechanical roles of F-actin in the differentiation of stem cells: A review. ACS Biomater. Sci. Eng. 2019, 5, 3788–3801. [Google Scholar] [CrossRef]
- Mo, W.; Wu, J.; Qiu, Q.; Zhang, F.; Luo, H.; Xu, N.; Zhu, W.; Liang, M. Platelet-rich plasma inhibits osteoblast apoptosis and actin cytoskeleton disruption induced by gingipains through upregulating integrin β1. Cell Biol. Int. 2020, 44, 2120–2130. [Google Scholar] [CrossRef]
- Titushkin, I.; Cho, M. Regulation of cell cytoskeleton and membrane mechanics by electric field: Role of linker proteins. Biophys. J. 2009, 96, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Azadi, S.; Tafazzoli-Shadpour, M.; Soleimani, M.; Warkiani, M.E. Modulating cancer cell mechanics and actin cytoskeleton structure by chemical and mechanical stimulations. J. Biomed. Mater. Res. Part A 2019, 107, 1569–1581. [Google Scholar] [CrossRef]
- Mack, C.P.; Somlyo, A.V.; Hautmann, M.; Somlyo, A.P.; Owens, G.K. Smooth muscle differentiation marker gene expression is regulated by RhoA-mediated actin polymerization. J. Biol. Chem. 2001, 276, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Martino, J.; Mascalchi, P.; Legros, P.; Lacomme, S.; Gontier, E.; Bioulac-Sage, P.; Balabaud, C.; Moreau, V.; Saltel, F. Actin depolymerization in dedifferentiated liver sinusoidal endothelial cells promotes fenestrae re-formation. Hepatol. Commun. 2019, 3, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izdebska, M.; Zielińska, W.; Grzanka, D.; Gagat, M. The role of actin dynamics and actin-binding proteins expression in epithelial-to-mesenchymal transition and its association with cancer progression and evaluation of possible therapeutic targets. BioMed Res. Int. 2018, 2018, 4578373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahuzier, A.; Shihavuddin, A.; Fournier, C.; Lansade, P.; Faucourt, M.; Menezes, N.; Meunier, A.; Garfa-Traoré, M.; Carlier, M.F.; Voituriez, R.; et al. Ependymal cilia beating induces an actin network to protect centrioles against shear stress. Nat. Commun. 2018, 9, 2279. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Fang, D.N.; Li, C.Q.; Hwang, K.C. Nonlinear electric–mechanical behavior and micromechanics modelling of cferroelectric domain evolution. Acta Mater. 1999, 47, 2913–2926. [Google Scholar] [CrossRef]
- Li, X.; Kolega, J. Effects of direct current electric fields on cell migration and actin filament distribution in bovine vascular endothelial cells. J. Vasc. Res. 2002, 39, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Rassokhin, M.A.; Pakhomov, A.G. Cellular regulation of extension and retraction of pseudopod-like blebs produced by nanosecond pulsed electric field (nsPEF). Cell Biochem. Biophys. 2014, 69, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Hunley, C.; Uribe, D.; Marucho, M. A multi-scale approach to describe electrical impulses propagating along actin filaments in both intracellular and in vitro conditions. RSC Adv. 2018, 8, 12017–12028. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Cheng, J.; Han Cheng, S.; Sun, D. Probing cell biophysical behavior based on actin cytoskeleton modeling and stretching manipulation with optical tweezers. Appl. Phys. Lett. 2013, 103, 083706. [Google Scholar] [CrossRef]
- Bai, G.; Li, Y.; Chu, H.K.; Wang, K.; Tan, Q.; Xiong, J.; Sun, D. Characterization of biomechanical properties of cells through dielectrophoresis-based cell stretching and actin cytoskeleton modeling. Biomed. Eng. Online 2017, 16, 41. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Huan, Z.; Hu, Y. Quantitative characterization of mechano-biological interrelationships of single cells. Int. J. Adv. Manuf. Technol. 2019, 105, 4967–4972. [Google Scholar] [CrossRef]
- Perrin, B.J.; Ervasti, J.M. The actin gene family: Function follows isoform. Cytoskeleton 2010, 67, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Lai, M. Actin, a reliable marker of internal control? Clin. Chim. Acta 2007, 385, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Le, S.; Efremov, A.K.; Zeng, X.; Bershadsky, A.; Yan, J. Effects of mechanical stimuli on profilin-and formin-mediated actin polymerization. Nano Lett. 2018, 18, 5239–5247. [Google Scholar] [CrossRef]
- Pollard, T.D. Regulation of actin filament assembly by Arp2/3 complex and formins. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 451–477. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006, 16, 522–529. [Google Scholar] [CrossRef]
- Guo, F.; Debidda, M.; Yang, L.; Williams, D.A.; Zheng, Y. Genetic deletion of Rac1 GTPase reveals its critical role in actin stress fiber formation and focal adhesion complex assembly. J. Biol. Chem. 2006, 281, 18652–18659. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Sun, D. Influence of semiflexible structural features of actin cytoskeleton on cell stiffness based on actin microstructural modeling. J. Biomech. 2012, 45, 1900–1908. [Google Scholar] [CrossRef]
- Palmer, J.S.; Boyce, M.C. Constitutive modeling of the stress–strain behavior of F-actin filament networks. Acta Biomater. 2008, 4, 597–612. [Google Scholar] [CrossRef]
- Chowdhury, F.; Na, S.; Li, D.; Poh, Y.C.; Tanaka, T.S.; Wang, F.; Wang, N. Material properties of the cell dictate stress-induced spreading and differentiation in embryonic stem cells. Nat. Mater. 2010, 9, 82–88. [Google Scholar] [CrossRef]
- Shao, M.; Vishvanath, L.; Busbuso, N.C.; Hepler, C.; Shan, B.; Sharma, A.X.; Chen, S.; Yu, X.; An, Y.A.; Zhu, Y.; et al. De novo adipocyte differentiation from Pdgfrβ+ preadipocytes protects against pathologic visceral adipose expansion in obesity. Nat. Commun. 2018, 9, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, T.; Luedtke, D.; Edwards, H.; Taub, J.W.; Ge, Y. A delicate balance–The BCL-2 family and its role in apoptosis, oncogenesis, and cancer therapeutics. Biochem. Pharmacol. 2019, 162, 250–261. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F-Actin Structural Parameter | Control | Day 2 | Day 4 | Day 6 |
|---|---|---|---|---|
| External radius (Rex) | 8.96 μm | 8.89 μm | 8.98 μm | 9.27 μm |
| Internal radius (Rin) | 6.02 μm | 6.13 μm | 6.34 μm | 6.75 μm |
| F-actin density (CAF) | 22.1 μM | 28.6 μM | 33.6 μM | 36.3 μM |
| Crosslink density ratio (R) | 0.02 | 0.02 | 0.02 | 0.02 |
| Number of F-actin in the network (NA) | 28,876 | 33,928 | 39,611 | 44,594 |
| Persistence length (Lp) | 3 μm | 3 μm | 3 μm | 3 μm |
| F-actin Structural Parameter | Control | Day 2 | Day 4 | Day 6 |
|---|---|---|---|---|
| External radius (Rex) | 7.33 μm | 7.51 μm | 7.45 μm | 7.61 μm |
| Internal radius (Rin) | 6.72 μm | 6.88 μm | 6.90 μm | 6.92 μm |
| F-actin density (CAF) | 74.6 μM | 74.1 μM | 71.2 μM | 70.8 μM |
| Crosslink density ratio (R) | 0.02 | 0.02 | 0.02 | 0.02 |
| Number of F-actin in the network (NA) | 87,656 | 83,231 | 78,943 | 72,881 |
| Persistence length (Lp) | 3 μm | 3 μm | 3 μm | 3 μm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhuang, X.; Niu, F. Quantitative Investigation of the Link between Actin Cytoskeleton Dynamics and Cellular Behavior. Micromachines 2022, 13, 1885. https://doi.org/10.3390/mi13111885
Li Y, Zhuang X, Niu F. Quantitative Investigation of the Link between Actin Cytoskeleton Dynamics and Cellular Behavior. Micromachines. 2022; 13(11):1885. https://doi.org/10.3390/mi13111885
Chicago/Turabian StyleLi, Ying, Xiaoru Zhuang, and Fuzhou Niu. 2022. "Quantitative Investigation of the Link between Actin Cytoskeleton Dynamics and Cellular Behavior" Micromachines 13, no. 11: 1885. https://doi.org/10.3390/mi13111885