
Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics




Abstract
:1. Introduction
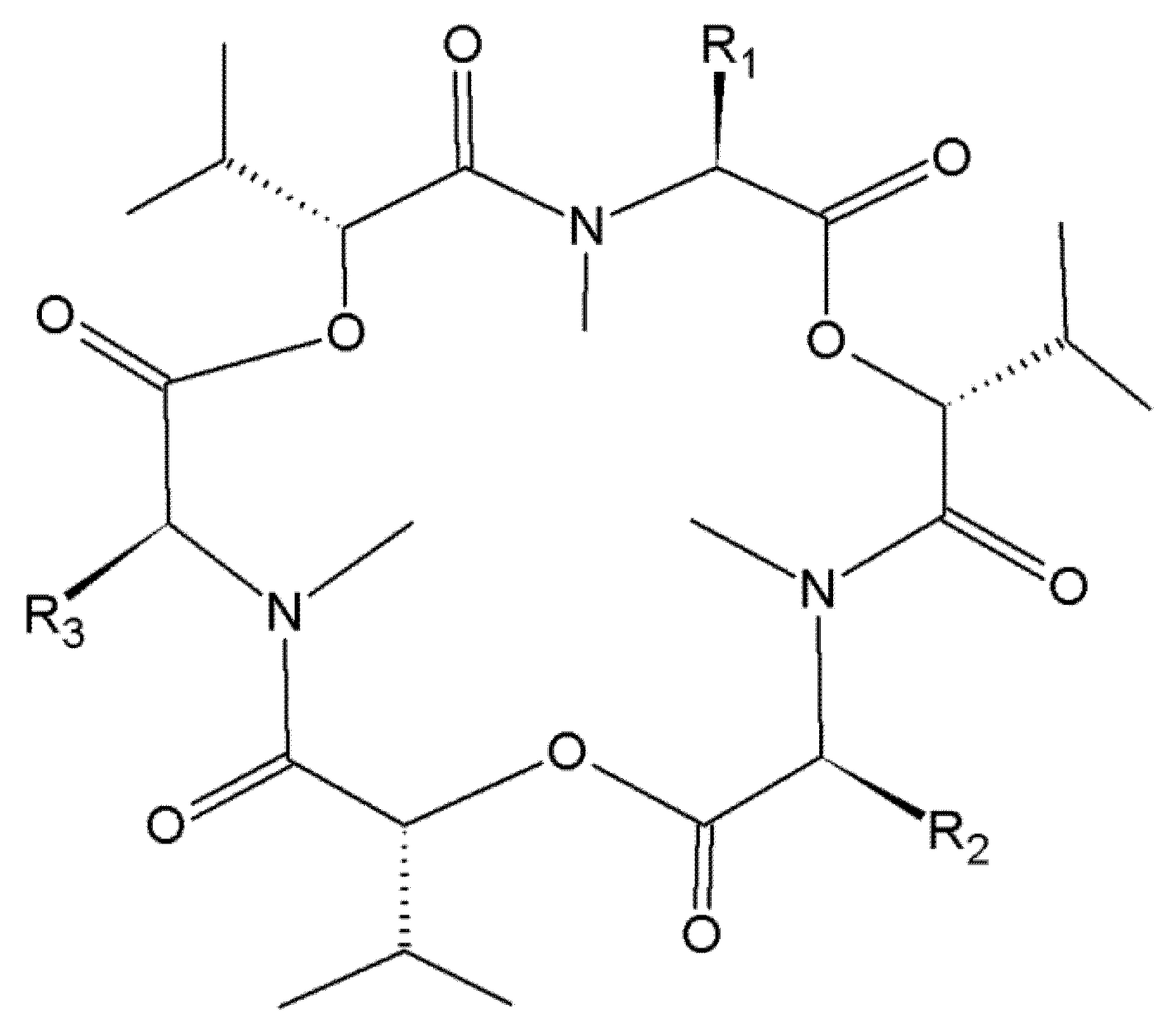
2. Beauvericin and Enniatins
2.1. Occurrence of Beauvericin and Enniatins in Food and Feed
2.2. Toxicity of Beauvericin and Enniatins
2.2.1. In Vitro Toxicity
2.2.2. In Vivo Toxicity
2.3. Toxicokinetics of Beauvericin and Enniatins
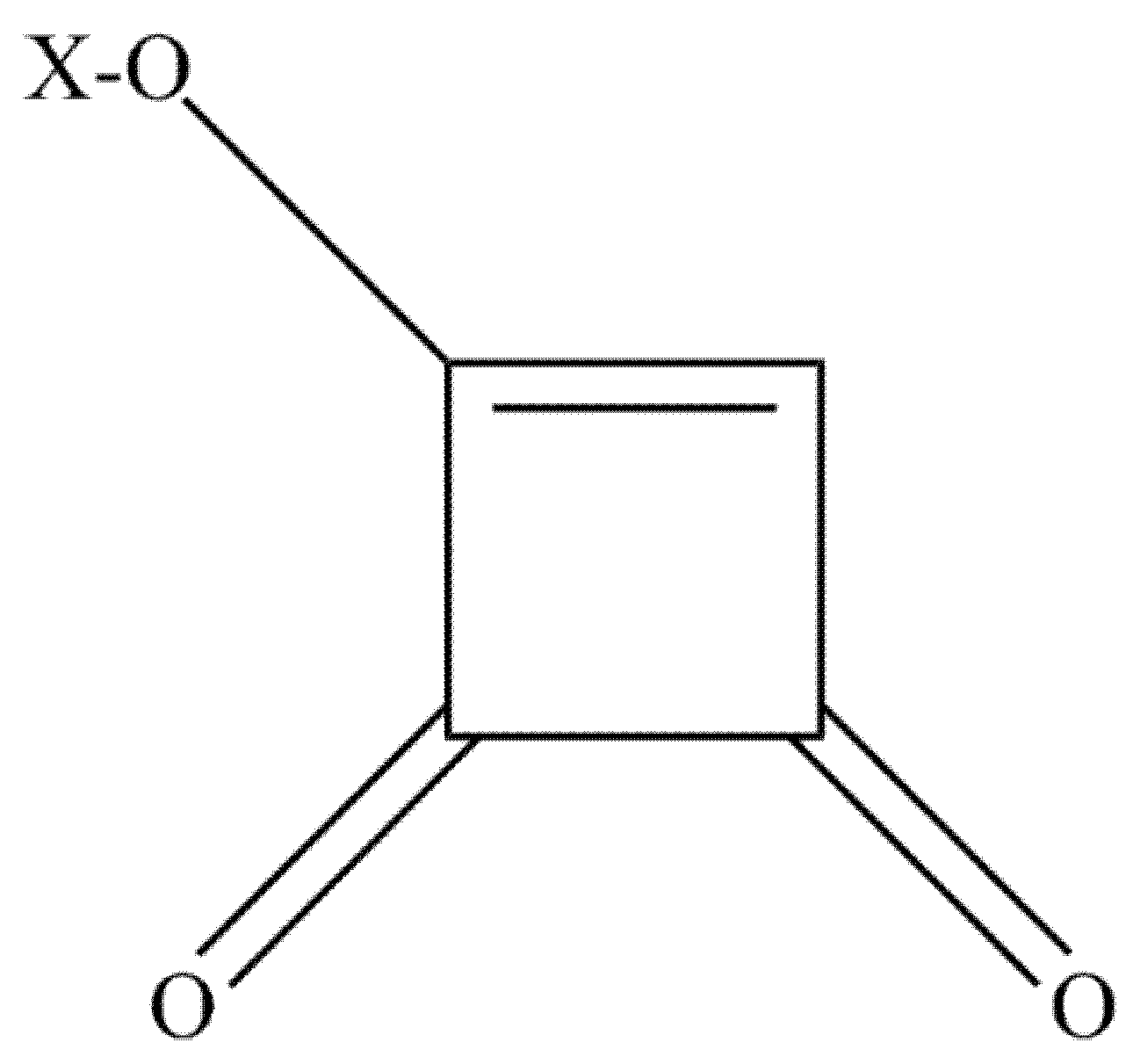
3. Moniliformin
3.1. Occurrence of Moniliformin
3.2. Toxicity of Moniliformin
3.3. Toxicokinetics of Moniliformin
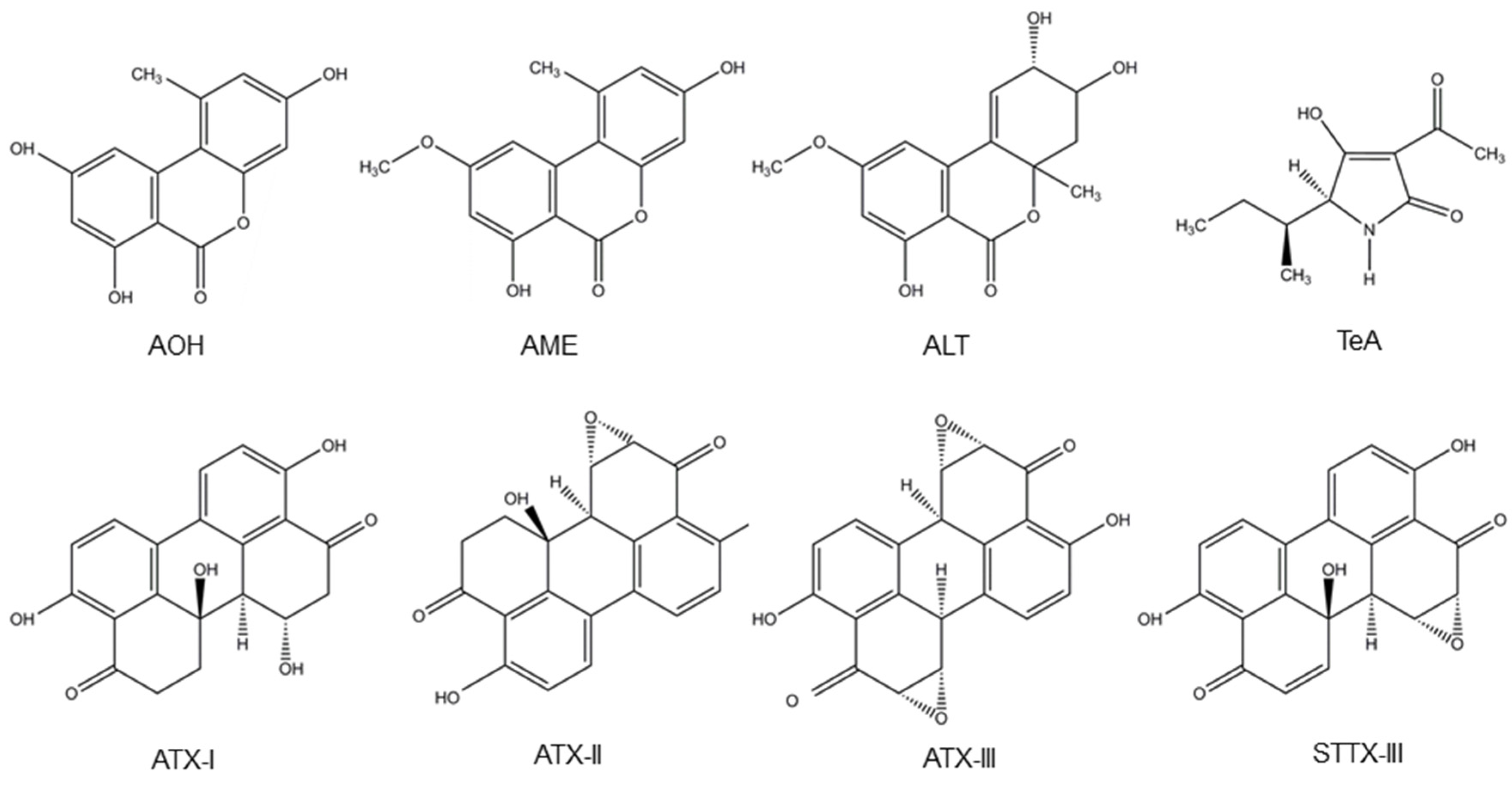
4. Alternaria Mycotoxins
4.1. Occurrence of Alternaria Mycotoxins
4.2. Toxicity of Alternaria Mycotoxins
4.3 Toxicokinetics of Alternaria Mycotoxins
5. Research Gaps
Author Contributions
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Sulyok, M.; Krska, R.; Schuhmacher, R. A liquid chromatography/tandem mass spectrometric multi-mycotoxin method for the quantification of 87 analytes and its application to semi-quantitative screening of moldy food samples. Anal. Bioanal. Chem. 2007, 389, 1505–1523. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M.J. Beauvericin-induced cytotoxicity via ROS production and mitochondrial damage in Caco-2 cells. Toxicol. Lett. 2013, 222, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M.J. Reactive oxygen species involvement in apoptosis and mitochondrial damage in Caco-2 cells induced by enniatins a, a(1), b and b(1). Toxicol. Lett. 2013, 222, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Çelik, M.; Aksoy, H.; Yilmaz, S. Evaluation of beauvericin genotoxicity with the chromosomal aberrations, sister-chromatid exchanges and micronucleus assays. Ecotoxicol. Environ. Saf. 2010, 73, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Çelik, M.; Yilmaz, S.; Aksoy, H.; Unal, F.; Yuzbasioglu, D.; Donbak, L. Evaluation of the genotoxicity of Fusarium mycotoxin moniliformin in human peripheral blood lymphocytes. Environ. Mol. Mutagen. 2009, 50, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Burkhardt, B.; Pfeiffer, E.; Metzler, M. Alternaria toxins: Altertoxin II is a much stronger mutagen and DNA strand breaking mycotoxin than alternariol and its methyl ether in cultured mammalian cells. Toxicol. Lett. 2012, 214, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Sauter, F.; Pfeiffer, E.; Metzler, M.; Hartwig, A.; Köberle, B. DNA damage and repair kinetics of the Alternaria mycotoxins alternariol, altertoxin II and stemphyltoxin III in cultured cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 798–799, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, D.R.; Broomhead, J.N.; Bermudez, A.J.; Rottinghaus, G.E. Individual and combined effects of the Fusarium mycotoxins fumonisin b1 and moniliformin in broiler chicks. Avian Dis. 2003, 47, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Asrani, R.K.; Ledoux, D.R.; Rottinghaus, G.E.; Gupta, V.K. Toxic interaction between fumonisin b-1 and moniliformin for cardiac lesions in Japanese quail. Avian Dis. 2012, 56, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.R.; Fredrickson, T.N.; Hadidian, Z. Toxic effects of the sodium and the n,n′-dibenzylethylenediamine salts of tenuazonic acid (nsc-525816 and nsc-82260). Cancer Chemother. Rep. 1968, 52, 579–585. [Google Scholar] [PubMed]
- Giambrone, J.J.; Davis, N.D.; Diener, U.L. Effect of tenuazonic acid on young chickens. Poult. Sci. 1978, 57, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in food and feed. EFSA J. 2011, 9, 97. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 174. [Google Scholar] [CrossRef]
- Hietaniemi, V.; Rämö, S.; Yli-Mattila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sieviläinen, E.; Koivisto, T.; Parikka, P. Updated survey of Fusarium species and toxins in Finnish cereal grains. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 831–848. [Google Scholar] [CrossRef] [PubMed]
- Hickert, S.; Bergmann, M.; Ersen, S.; Cramer, B.; Humpf, H.U. Survey of Alternaria toxin contamination in food from the German market, using a rapid HPLC-MS/MS approach. Mycotoxin Res. 2016, 32, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asam, S.; Devos, T.; Ediage, E.N.; Di Mavungu, J.D.; Jacxsens, L.; Van Landschoot, A.; Vanhaecke, L.; et al. Validated UPLC-MS/MS methods to quantitate free and conjugated Alternaria toxins in commercially available tomato products and fruit and vegetable juices in Belgium. J. Agric. Food Chem. 2016, 64, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; Broekaert, N.; De Mil, T.; Fraeyman, S.; De Backer, P.; Croubels, S. Pilot toxicokinetic study and absolute oral bioavailability of the Fusarium mycotoxin enniatin b1 in pigs. Food Chem. Toxicol. 2014, 63, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Fraeyman, S.; Devreese, M.; Antonissen, G.; De Baere, S.; Rychlik, M.; Croubels, S. Comparative oral bioavailability, toxicokinetics, and biotransformation of enniatin b1 and enniatin b in broiler chickens. J. Agric. Food Chem. 2016, 64, 7259–7264. [Google Scholar] [CrossRef] [PubMed]
- Fraeyman, S.; Devreese, M.; Broekaert, N.; De Mil, T.; Antonissen, G.; De Baere, S.; De Backer, P.; Rychlik, M.; Croubels, S. Quantitative determination of tenuazonic acid in pig and broiler chicken plasma by LC-MS/MS and its comparative toxicokinetics. J. Agric. Food Chem. 2015, 63, 8560–8567. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, L.; Uhlig, S.; Devreese, M.; Croubels, S.; Faeste, C.K. Biotransformation of the mycotoxin enniatin b1 in pigs: A comparative in vitro and in vivo approach. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2017, 105, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging mycotoxins: Beyond traditionally determined food contaminants. J. Agric. Food Chem. 2016. [CrossRef] [PubMed]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. Structure of beauvericin, a new depsipeptide antibiotic toxic to artemia salina. Tetrahedron Lett. 1969, 10, 4255–4258. [Google Scholar] [CrossRef]
- Thrane, U.; Adler, A.; Clasen, P.E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [PubMed]
- Logrieco, A.; Rizzo, A.; Ferracane, R.; Ritieni, A. Occurrence of beauvericin and enniatins in wheat affected by Fusarium avenaceum head blight. Appl. Environ. Microbiol. 2002, 68, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Zocher, R.; Haese, A. Enniatin production by Fusarium strains and its effect on potato tuber tissue. Appl. Environ. Microbiol. 1996, 62, 393–398. [Google Scholar] [PubMed]
- Christ, D.S.; Godecke, R.; von Tiedemann, A.; Varrelmann, M. Pathogenicity, symptom development, and mycotoxin formation in wheat by Fusarium species frequently isolated from sugar beet. Phytopathology 2011, 101, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, M.; Gidlund, A.; Sulyok, M.; Borjesson, T.; Krska, R.; Olsen, M.; Fredlund, E. Deoxynivalenol and other selected Fusarium toxins in Swedish wheat—Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Fredlund, E.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Lindblad, M. Deoxynivalenol and other selected Fusarium toxins in Swedish oats—Occurrence and correlation to specific Fusarium species. Int. J. Food. Microbiol. 2013, 167, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Mahnine, N.; Meca, G.; Elabidi, A.; Fekhaoui, M.; Saoiabi, A.; Font, G.; Manes, J.; Zinedine, A. Further data on the levels of emerging Fusarium mycotoxins enniatins (a, a1, b, b1), beauvericin and fusaproliferin in breakfast and infant cereals from Morocco. Food Chem. 2011, 124, 481–485. [Google Scholar] [CrossRef]
- Sifou, A.; Meca, G.; Serrano, A.B.; Mahnine, N.; El Abidi, A.; Manes, J.; El Azzouzi, M.; Zinedine, A. First report on the presence of emerging Fusarium mycotoxins enniatins (a, a-1, b, b-1), beauvericin and fusaproliferin in rice on the Moroccan retail markets. Food Control 2011, 22, 1826–1830. [Google Scholar] [CrossRef]
- Zinedine, A.; Fernandez-Franzon, M.; Manes, J.; Manyes, L. Multi-mycotoxin contamination of couscous semolina commercialized in Morocco. Food Chem. 2017, 214, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Covarelli, L.; Beccari, G.; Colasante, V.; Manes, J. Simultaneous analysis of twenty-six mycotoxins in durum wheat grain from Italy. Food Control 2016, 62, 322–329. [Google Scholar] [CrossRef]
- Serrano, A.B.; Font, G.; Manes, J.; Ferrer, E. Emerging Fusarium mycotoxins in organic and conventional pasta collected in Spain. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 51, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, N.; Yang, L.; Deng, Y.; Wang, J.; Song, S.; Lin, S.; Wu, A.; Zhou, Z.; Hou, J. Multi-mycotoxin analysis of animal feed and animal-derived food using LC-MS/MS system with timed and highly selective reaction monitoring. Anal. Bioanal. Chem. 2015, 407, 7359–7368. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky, P.; Kos, G.; Nahrer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize-an extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [PubMed]
- De Lourdes Mendes de Souza, M.; Sulyok, M.; Freitas-Silva, O.; Costa, S.S.; Brabet, C.; Machinski Junior, M.; Sekiyama, B.L.; Vargas, E.A.; Krska, R.; Schuhmacher, R. Cooccurrence of mycotoxins in maize and poultry feeds from brazil by liquid chromatography/tandem mass spectrometry. Sci. World J. 2013, 2013, 427369. [Google Scholar] [CrossRef]
- Scarpino, V.; Blandino, M.; Negre, M.; Reyneri, A.; Vanara, F. Moniliformin analysis in maize samples from north-west Italy using multifunctional clean-up columns and the LC-MS/MS detection method. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2013, 30, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.E.H.; Korn, U. Alternaria mycotoxins in wheat—A 10 years survey in the northeast of Germany. Food Control 2013, 34, 191–197. [Google Scholar] [CrossRef]
- Janić Hajnal, E.; Orčić, D.; Torbica, A.; Kos, J.; Mastilović, J.; Škrinjar, M. Alternaria toxins in wheat from the autonomous province of Vojvodina, Serbia: A preliminary survey. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.J.; Han, X.M.; Li, F.Q.; Zhang, L.S. Natural occurrence of alternaria toxins in the 2015 wheat from Anhui province, China. Toxins 2016, 8, 308. [Google Scholar] [CrossRef] [PubMed]
- Hickert, S.; Gerding, J.; Ncube, E.; Hubner, F.; Flett, B.; Cramer, B.; Humpf, H.U. A new approach using micro HPLC-MS/MS for multi-mycotoxin analysis in maize samples. Mycotoxin Res. 2015, 31, 109–115. [Google Scholar] [CrossRef] [PubMed]
- López, P.; Venema, D.; de Rijk, T.; de Kok, A.; Scholten, J.M.; Mol, H.G.J.; de Nijs, M. Occurrence of Alternaria toxins in food products in the Netherlands. Food Control 2016, 60, 196–204. [Google Scholar] [CrossRef]
- López, P.; Venema, D.; Mol, H.; Spanjer, M.; de Stoppelaar, J.; Pfeiffer, E.; de Nijs, M. Alternaria toxins and conjugates in selected foods in the Netherlands. Food Control 2016, 69, 153–159. [Google Scholar] [CrossRef]
- Zwickel, T.; Klaffke, H.; Richards, K.; Rychlik, M. Development of a high performance liquid chromatography tandem mass spectrometry based analysis for the simultaneous quantification of various Alternaria toxins in wine, vegetable juices and fruit juices. J. Chromatogr. A 2016, 1455, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Asam, S.; Rychlik, M. Potential health hazards due to the occurrence of the mycotoxin tenuazonic acid in infant food. Eur. Food Res. Technol. 2013, 236, 491–497. [Google Scholar] [CrossRef]
- Storm, I.M.; Rasmussen, R.R.; Rasmussen, P.H. Occurrence of pre- and post-harvest mycotoxins and other secondary metabolites in Danish maize silage. Toxins (Basel) 2014, 6, 2256–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monbaliu, S.; Van Poucke, C.; Detavernier, C.; Dumoulin, F.; Van De Velde, M.; Schoeters, E.; Van Dyck, S.; Averkieva, O.; Van Peteghem, C.; De Saeger, S. Occurrence of mycotoxins in feed as analyzed by a multi-mycotoxin LC-MS/MS method. J. Agric. Food Chem. 2010, 58, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Zouaoui, N.; Mallebrera, B.; Berrada, H.; Abid-Essefi, S.; Bacha, H.; Ruiz, M.J. Cytotoxic effects induced by patulin, sterigmatocystin and beauvericin on CHO-K1 cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2016, 89, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Ficheux, A.S.; Sibiril, Y.; Parent-Massin, D. Effects of beauvericin, enniatin b and moniliformin on human dendritic cells and macrophages: An in vitro study. Toxicon 2013, 71, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jow, G.M.; Chou, C.J.; Chen, B.F.; Tsai, J.H. Beauvericin induces cytotoxic effects in human acute lymphoblastic leukemia cells through cytochrome c release, caspase 3 activation: The causative role of calcium. Cancer Lett. 2004, 216, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, A.; Karlsøen, L.M.; Holme, J.A.; Kristoffersen, A.B.; Eriksen, G.S. Immunomodulatory effects of individual and combined mycotoxins in the THP-1 cell line. Toxicol. In Vitro 2016, 36, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Font, G.; Ruiz, M.J. Comparative cytotoxicity study of enniatins a, a(1), a(2), b, b(1), b(4) and j(3) on Caco-2 cells, Hep-G(2) and HT-29. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 2464–2469. [Google Scholar] [CrossRef] [PubMed]
- Kamyar, M.; Rawnduzi, P.; Studenik, C.R.; Kouri, K.; Lemmens-Gruber, R. Investigation of the electrophysiological properties of enniatins. Arch. Biochem. Biophys. 2004, 429, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Kouri, K.; Lemmens, M.; Lemmens-Gruber, R. Beauvericin-induced channels in ventricular myocytes and liposomes. Biochim. Biophys. Acta 2003, 1609, 203–210. [Google Scholar] [CrossRef]
- Springler, A.; Vrubel, G.J.; Mayer, E.; Schatzmayr, G.; Novak, B. Effect of Fusarium-derived metabolites on the barrier integrity of differentiated intestinal porcine epithelial cells (IPEC-J2). Toxins (Basel) 2016, 8, E345. [Google Scholar] [CrossRef] [PubMed]
- Žužek, M.C.; Grandič, M.; Jakovac Strajn, B.; Frangež, R. Beauvericin inhibits neuromuscular transmission and skeletal muscle contractility in mouse hemidiaphragm preparation. Toxicol. Sci. 2016, 150, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Schoevers, E.J.; Santos, R.R.; Fink-Gremmels, J.; Roelen, B.A. Toxicity of beauvericin on porcine oocyte maturation and preimplantation embryo development. Reprod. Toxicol. 2016, 65, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Albonico, M.; Schutz, L.F.; Caloni, F.; Cortinovis, C.; Spicer, L.J. In vitro effects of the Fusarium mycotoxins fumonisin b1 and beauvericin on bovine granulosa cell proliferation and steroid production. Toxicon 2017, 128, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Kalayou, S.; Ndossi, D.; Frizzell, C.; Groseth, P.K.; Connolly, L.; Sørlie, M.; Verhaegen, S.; Ropstad, E. An investigation of the endocrine disrupting potential of enniatin b using in vitro bioassays. Toxicol. Lett. 2015, 233, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Sospedra, I.; Soriano, J.M.; Ritieni, A.; Moretti, A.; Manes, J. Antibacterial effect of the bioactive compound beauvericin produced by Fusarium proliferatum on solid medium of wheat. Toxicon 2010, 56, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Roig, M.; Meca, G.; Marin, R.; Ferrer, E.; Manes, J. Antibacterial activity of the emerging Fusarium mycotoxins enniatins a, a(1), a(2), b, b-1, and b-4 on probiotic microorganisms. Toxicon 2014, 85, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cetin, Y.; Bullerman, L.B. Cytotoxicity of Fusarium mycotoxins to mammalian cell cultures as determined by the mtt bioassay. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2005, 43, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Blanco, C.; Juan-García, A.; Juan, C.; Font, G.; Ruiz, M.J. Alternariol induce toxicity via cell death and mitochondrial damage on Caco-2 cells. Food Chem. Toxicol. 2016, 88, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Bensassi, F.; Gallerne, C.; El Dein, O.S.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Cell death induced by the Alternaria mycotoxin alternariol. Toxicol. In Vitro 2012, 26, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Bensassi, F.; Gallerne, C.; El Dein, O.S.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Mechanism of alternariol monomethyl ether-induced mitochondrial apoptosis in human colon carcinoma cells. Toxicology 2011, 290, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Kalayou, S.; Hamre, A.G.; Ndossi, D.; Connolly, L.; Sørlie, M.; Ropstad, E.; Verhaegen, S. Using SILAC proteomics to investigate the effect of the mycotoxin, alternariol, in the human H295R steroidogenesis model. Cell Biol. Toxicol. 2014, 30, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, U.; Tomek, W.; Schneider, F.; Müller, M.; Pöhland, R.; Vanselow, J. The mycotoxins alternariol and alternariol methyl ether negatively affect progesterone synthesis in porcine granulosa cells in vitro. Toxicol. Lett. 2009, 186, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Ndossi, D.; Kalayou, S.; Eriksen, G.S.; Verhaegen, S.; Sørlie, M.; Elliott, C.T.; Ropstad, E.; Connolly, L. An in vitro investigation of endocrine disrupting effects of the mycotoxin alternariol. Toxicol. Appl. Pharmacol. 2013, 271, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, L.; Wagner, J.; Metzler, M. Estrogenic and clastogenic potential of the mycotoxin alternariol in cultured mammalian cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2006, 44, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Fernández-Franzón, M.; Font, G.; Ruiz, M.J. Toxicity evaluation of individual and mixed enniatins using an in vitro method with CHO-K1 cells. Toxicol. In Vitro 2013, 27, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, A.; Wisbech, C.; Christoffersen, T.E.; Hult, L.O.; Lea, T.; Eriksen, G.S.; Holme, J.A. The mycotoxin alternariol induces DNA damage and modify macrophage phenotype and inflammatory responses. Toxicol. Lett. 2015, 239, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Manyes, L.; Font, G.; Juan-Garcia, A. Evaluation of immunologic effect of enniatin a and quantitative determination in feces, urine and serum on treated wistar rats. Toxicon 2014, 87, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.F.; Xu, R.; Ouyang, Z.J.; Qian, C.; Shen, Y.; Wu, X.D.; Gu, Y.H.; Xu, Q.; Sun, Y. Beauvericin ameliorates experimental colitis by inhibiting activated T cells via downregulation of the pi3k/akt signaling pathway. PLoS ONE 2013, 8, e83013. [Google Scholar] [CrossRef] [PubMed]
- Yuca, K.; Cankaya, H.; Bayram, W.; Ozbek, H.; Kiris, M. Local irritant effects of topical oral sprays on oral mucosa in mice. Adv. Ther. 2006, 23, 98–106. [Google Scholar] [CrossRef] [PubMed]
- EMA. Cmdh Endorses Revocation of Authorisations for Fusafungine Sprays Used to Treat Airway Infections; EMA/227560/2016; European Medicine Agency: London, UK, 2016. [Google Scholar]
- Manyes, L.; Escrivá, L.; Serrano, A.B.; Rodríguez-Carrasco, Y.; Tolosa, J.; Meca, G.; Font, G. A preliminary study in wistar rats with enniatin a contaminated feed. Toxicol. Mech. Methods 2014, 24, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Callebaut, F.; Tangni, E.K.; Debongnie, P.; Stals, E.; Huybrechts, B.; Waegeneers, N.; Delezie, E.; Van Pamel, E.; Daeseleire, E. Carry-Over of Mycotoxins to Animal Products: Case Study Poultry; Scientifec Report 211/212 CODA-CERVA (Centrum voor Onderzoek in Diergeneeskunde en Agrochemie-Centre d'Étude et de Recherches Vétérinaires et Agrochemiques): Brussels, Belgium, 2011–2012; pp. 141–144. [Google Scholar]
- Leitgeb, R.; Raffaseder, C.; Ruckenbauer, P.; Lemmens, M.; Böhm, J.; Wagner, E.; Krska, R.; Parich, A. [impact of Fusarium toxins on growth and slaughter performance of broilers and turkeys]. Mycotoxin Res. 2003, 19, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Dorne, J.L.; Fink-Gremmels, J. Human and animal health risk assessments of chemicals in the food chain: Comparative aspects and future perspectives. Toxicol. Appl. Pharmacol. 2013, 270, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, L.; Faeste, C.K.; Delezie, E.; Van Pamel, E.; Daeseleire, E.; Callebaut, A.; Uhlig, S. Presence of enniatin b and its hepatic metabolites in plasma and liver samples from broilers and eggs from laying hens. World Mycotoxin J. 2014, 7, 167–175. [Google Scholar] [CrossRef]
- Barnes, D.M. Expression of p-glycoprotein in the chicken. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2001, 130, 301–310. [Google Scholar] [CrossRef]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Fricker, G.; Miller, D.S. Relevance of multidrug resistance proteins for intestinal drug absorption in vitro and in vivo. Pharmacol. Toxicol. 2002, 90, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Shugarts, S.; Benet, L.Z. The role of transporters in the pharmacokinetics of orally administered drugs. Pharm. Res. 2009, 26, 2039–2054. [Google Scholar] [CrossRef] [PubMed]
- Dornetshuber, R.; Heffeter, P.; Sulyok, M.; Schumacher, R.; Chiba, P.; Kopp, S.; Koellensperger, G.; Micksche, M.; Lemmens-Gruber, R.; Berger, W. Interactions between ABC-transport proteins and the secondary Fusarium metabolites enniatin and beauvericin. Mol. Nutr. Food Res. 2009, 53, 904–920. [Google Scholar] [CrossRef] [PubMed]
- Commission Regulation (EC) No. 495/2011 of 20 may 2011 Amending Regulation (EC) No. 109/2007 as Regards the Composition of the Feed Additive Monensin Sodium; Official Journal of the European Union: Brussels, Belgium, 2011; Volume L 134/6.
- Jestoi, M.; Rokka, M.; Järvenpää, E.; Peltonen, K. Determination of Fusarium mycotoxins beauvericin and enniatins (a, a1, b, b1) in eggs of laying hens using liquid chromatography-tandem mass spectrometry (LC-MS/MS). Food Chem. 2009, 115, 1120–1127. [Google Scholar] [CrossRef]
- Burmeister, H.R.; Ciegler, A.; Vesonder, R.F. Moniliformin, a metabolite of Fusarium moniliforme nrrl 6322: Purification and toxicity. Appl. Environ. Microbiol. 1979, 37, 11–13. [Google Scholar] [PubMed]
- Springer, J.P.; Clardy, J.; Cole, R.J.; Kirksey, J.W.; Hill, R.K.; Carlson, R.M.; Isidor, J.L. Structure and synthesis of moniliformin, a novel cyclobutane microbial toxin. J. Am. Chem. Soc. 1974, 96, 2267–2268. [Google Scholar] [CrossRef] [PubMed]
- Seifert, K.A.; Aoki, T.; Baayen, R.P.; Brayford, D.; Burgess, L.W.; Chulze, S.; Gams, W.; Geiser, D.; de Gruyter, J.; Leslie, J.F.; et al. The name Fusarium moniliforme should no longer be used. Mycol. Res. 2003, 107, 643–644. [Google Scholar] [CrossRef]
- Fotso, J.; Leslie, J.F.; Smith, J.S. Production of beauvericin, moniliformin, fusaproliferin, and fumonisins b-1, b-2, and b-3 by fifteen ex-type strains of Fusarium species. Appl. Environ. Microbiol. 2002, 68, 5195–5197. [Google Scholar] [CrossRef] [PubMed]
- Schütt, F.; Nirenberg, H.I.; Demi, G. Moniliformin production in the genus Fusarium. Mycotoxin Res. 1998, 14, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Hallas-Møller, M.; Nielsen, K.F.; Frisvad, J.C. Production of the Fusarium mycotoxin moniliformin by Penicillium melanoconidium. J. Agric. Food Chem. 2016, 64, 4505–4510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, M.; Jestoi, M.; Nathanail, A.V.; Kokkonen, U.M.; Anttila, M.; Koivisto, P.; Karhunen, P.; Peltonen, K. Application of OECD guideline 423 in assessing the acute oral toxicity of moniliformin. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 53, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Atosuo, J.; Jestoi, M.; Nathanail, A.V.; Kokkonen, U.M.; Anttila, M.; Koivisto, P.; Lilius, E.M.; Peltonen, K. Repeated dose 28-day oral toxicity study of moniliformin in rats. Toxicol. Lett. 2015, 233, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Pollock, G.A.; Disabatino, C.E.; Heimsch, R.C.; Hilbelink, D.R. The subchronic toxicity and teratogenicity of alternariol monomethyl ether produced by Alternaria solani. Food Chem. Toxicol. 1982, 20, 899–902. [Google Scholar] [CrossRef]
- Griffin, G.F.; Chu, F.S. Toxicity of the Alternaria metabolites alternariol, alternariol methyl-ether, altenuene, and tenuazonic acid in the chicken-embryo assay. Appl. Environ. Microbiol. 1983, 46, 1420–1422. [Google Scholar] [PubMed]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic effects of moniliformin in broilers and turkeys fed dietary treatments to market age. Avian Dis. 2002, 46, 901–908. [Google Scholar] [CrossRef]
- Li, Y.C.; Ledoux, D.R.; Bermudez, A.J.; Fritsche, K.L.; Rottinghaus, G.E. The individual and combined effects of fumonisin b1 and moniliformin on performance and selected immune parameters in turkey poults. Poult. Sci. 2000, 79, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.; Edrington, T.S.; Kubena, L.F.; Rottinghaus, G.E.; Turk, J.R.; Genovese, K.J.; Ziprin, R.L.; Nisbet, D.J. Toxicity of fumonisin from Fusarium verticillioides culture material and moniliformin from Fusarium fujikuroi culture material when fed singly and in combination to growing barrows. J. Food Prot. 2002, 65, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Pirrung, M.C.; Nauhaus, S.K.; Singh, B. Cofactor-directed, time-dependent inhibition of thiamine enzymes by the fungal toxin moniliformin. J. Org. Chem. 1996, 61, 2592–2593. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Huwel, S.; Galla, H.J.; Humpf, H.U. Blood-brain barrier effects of the Fusarium mycotoxins deoxynivalenol, 3 acetyldeoxynivalenol, and moniliformin and their transfer to the brain. PLoS ONE 2015, 10, e0143640. [Google Scholar] [CrossRef] [PubMed]
- Garganese, F.; Schena, L.; Siciliano, I.; Prigigallo, M.I.; Spadaro, D.; De Grassi, A.; Ippolito, A.; Sanzani, S.M. Characterization of citrus-associated Alternaria species in mediterranean areas. PLoS ONE 2016, 11, e0163255. [Google Scholar] [CrossRef] [PubMed]
- Davis, V.M.; Stack, M.E. Mutagenicity of stemphyltoxin iii, a metabolite of Alternaria alternata. Appl. Environ. Microbiol. 1991, 57, 180–182. [Google Scholar] [PubMed]
- Taniwaki, M.H.; Frisvad, J.C.; Ferranti, L.S.; Lopes, A.D.; Larsen, T.O.; Fungaro, M.H.P.; Iamanaka, B.T. Biodiversity of mycobiota throughout the Brazil nut supply chain: From rainforest to consumer. Food Microbiol. 2017, 61, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, A.; Eriksen, G.S.; Holme, J.A. Mechanisms of action and toxicity of the mycotoxin alternariol: A review. Basic Clin. Pharmacol. Toxicol. 2016, 119, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Fehr, M.; Pahlke, G.; Fritz, J.; Christensen, M.O.; Boege, F.; Altemöller, M.; Podlech, J.; Marko, D. Alternariol acts as a topoisomerase poison, preferentially affecting the II alpha isoform. Mol. Nutr. Food Res. 2009, 53, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, A.; Torgersen, M.L.; Holme, J.A.; Lagadic-Gossmann, D.; Eriksen, G.S. Autophagy and senescence, stress responses induced by the DNA-damaging mycotoxin alternariol. Toxicology 2014, 326, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeura, H.T.; Gordon, C.N. Biological activity of tenuazonic acid. Biochemistry 1963, 2, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, M.; Lepper, H.; Weidner, C.; Asam, S. Risk evaluation of the Alternaria mycotoxin tenuazonic acid in foods for adults and infants and subsequent risk management. Food Control 2016, 68, 181–185. [Google Scholar] [CrossRef]
- Asam, S.; Habler, K.; Rychlik, M. Determination of tenuazonic acid in human urine by means of a stable isotope dilution assay. Anal. Bioanal. Chem. 2013, 405, 4149–4158. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, S.; Ziemann, C.; Hansen, T. External Scientific Report on Combined Toxicokinetic and in vivo Genotoxicity Study on Alternaria Toxins; EN-679; EFSA Supporting Publication: Hannover, Germany, 2014. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Commodity | Mycotoxin | Total # Samples Analyzed | Prevalence Range (%) | Concentration Range (µg/kg) | Origin of Sample with Maximum Concentration | Reference |
|---|---|---|---|---|---|---|
| Cereals (unprocessed) a | BEA | 482 | 12–100 | <10–327 | Sweden | [17,31,32] |
| ENNs b | 482 | 96–100 | Finland | [17,31,32] | ||
| MON | 590 | 0.88–100 | <15–2606 | Italy | [17,31,32,41] | |
| AOH | 1582 | 2.4–47 | 0.75–832 | Germany | [42,43,44,45,46] | |
| AME | 1582 | 3.1–7.1 | 0.3–905 | Germany | [42,43,44,45,46] | |
| TeA | 1526 | 15–68 | 0.1–4224 | Germany | [42,43,44] | |
| ALT | 1106 | 2.63–7.1 | 6–196.6 | Germany | [42,45] | |
| ATX-I | 42 | 2.4 | 43 | South Africa | [45] | |
| TEN | 370 | 77 | 0.4–258.6 | China | [44] | |
| Rice | BEA | 70 | 75.7 | 3800–26,300 | Morocco | [34] |
| ENN A | 70 | 22.8 | 8400–119,500 | Morocco | [34] | |
| ENN A1 | 70 | 5.7 | 56,200–448,700 | Morocco | [34] | |
| ENN B | 70 | 30 | 4400–26,200 | Morocco | [34] | |
| ENN B1 | 70 | 24.3 | 3600–23,700 | Morocco | [34] | |
| Cereal products | BEA | 354 | 0–17.5 | 0.1–10,600 | Morocco | [33,35,36,37] |
| ENN A | 354 | 2.9–77 | 0.5–29,700 | Morocco | [33,35,36,37] | |
| ENN A1 | 354 | 30–100 | 0.25–688,000 | Morocco | [33,35,36,37] | |
| ENN B | 354 | 13.2–100 | 0.5–81,100 | Morocco | [33,35,36,37] | |
| ENN B1 | 354 | 17.6–100 | 0.5–795,000 | Morocco | [33,35,36,37] | |
| AOH | 83 | 31–89 | 8–121 | Italy | [18,36] | |
| AME | 83 | 26–89 | <0.4–48 | Italy | [18,36] | |
| TeA | 9 | 100 | <100–210 | Germany | [18] | |
| TEN | 9 | 100 | <1.6–12 | Germany | [18] | |
| Tomato products c | AOH | 187 | 28–70.6 | <2–41.6 | Belgium | [18,19,46,47] |
| AME | 187 | 20–79 | <0.9–7.8 | The Netherlands | [18,19,46,47] | |
| TeA | 187 | 40–100 | <5–462 | The Netherlands | [18,19,46,47] | |
| ALT | 83 | 32–56 | 6.1–62.0 | Belgium | [19] | |
| TEN | 117 | 21–64 | <3.9–8.9 | Belgium | [18,19] | |
| AOH-3-sulfate | 83 | 11–26 | 2.6–8.7 | Belgium | [19] | |
| AME-3-sulfate | 83 | 32–78 | 1.7–9.9 | Belgium | [19] | |
| Fruit juices d | AOH | 101 | 15–100 | <0.2–16 | Germany | [18,48] |
| AME | 101 | 25–100 | <0.13–4.9 | Germany | [18,48] | |
| TeA | 101 | 8–100 | <1.1–250 | Germany | [18,48] | |
| ALT | 101 | 4.3–100 | 1.18–18.4 | Germany | [18,48] | |
| TEN | 101 | 22–100 | <0.5–10.27 | Germany | [18,48] | |
| altenuic acid | 78 | 100 | 2.71 | Germany | [48] | |
| altenuisol | 78 | 8–50 | <0.74–5.58 | Germany | [48] | |
| Infant food e | TeA | 40 | 100 | 0.8–1200 | Germany | [49] |
| Wines | AOH | 30 | 20–93 | 0.65–11 | The Netherland | [46,48] |
| AME | 25 | 93 | 0.8–1.45 | Germany | [48] | |
| TeA | 25 | 60–100 | <1–60 | Germany | [48] | |
| TEN | 25 | 71 | 1.01–1.47 | Germany | [48] | |
| altenuic acid | 25 | 21–64 | <1–6.1 | Germany | [48] | |
| altenuisol | 25 | 55–71 | <0.74–2.91 | Germany | [48] | |
| Dried figs and olives | AOH | 14 | 7 | 8.7 | The Netherlands | [47] |
| TeA | 19 | 10–100 | 5.3–2345 | The Netherlands | [46,47] | |
| Vegetable oil | AOH | 19 | 47 | ≤6 | Germany | [18] |
| AME | 19 | 84 | <1.1–14 | Germany | [18] | |
| TeA | 19 | 21 | 15 | Germany | [18] | |
| TEN | 19 | 47 | <6.6–11 | Germany | [18] | |
| Sunflower seeds and oils | AOH | 35 | 10–55 | <4.9–39 | Germany | [18,47] |
| AME | 35 | 9–64 | <0.5–17 | The Netherlands | [18,47] | |
| TeA | 40 | 80–100 | <5–1350 | The Netherlands | [18,46,47] | |
| ALT | 11 | 9 | <14 | Germany | [18] | |
| ATX-I | 11 | 9 | <45 | Germany | [18] | |
| TEN | 16 | 20–91 | <3.7–800 | Germany | [18,46] | |
| Feed | BEA | 1345 | 50–98 | <2–2326 | not specified | [4,38,39,40] |
| ENN A | 1315 | 0–87 | <0.1–1745 | not specified | [4,39,40] | |
| ENN A1 | 1315 | 12–95 | <0.15–2216 | not specified | [4,39,40] | |
| ENN B | 1414 | 28–92 | <0.3–1514 | not specified | [4,39,40,50] | |
| ENN B1 | 1315 | 12–92 | <0.2–1846 | not specified | [4,39,40] | |
| MON | 1315 | 3–79 | <2–12,236 | not specified | [4,39,40] | |
| AOH | 264 | 0–80 | 17–221 | not specified | [4,50,51] | |
| AME | 264 | 1.5–82 | <6–733 | not specified | [4,50,51] | |
| TeA | 83 | 65 | not specified-1983 | not specified | [4] |
| Cell Line | Mycotoxin | Exposure Time | Exposure Dose (µM) | Effect | Reference |
|---|---|---|---|---|---|
| Caco-2 a | BEA | 0 min | 1.5 | ROS b generation | [5,6,56,66,67] |
| 24–72 h | IC50: 20.6–3.2 µM (MTT c); IC50: 8.8–1.9 µM (NR d) | ||||
| 24–72 h | 1.5–3.0 | LPO e, ↓ GSH, ↑ GSSG, loss of mitochondrial membrane potential, cell cycle arrest in S and G2/M, apoptosis and necrosis | |||
| 24 h | 12 | DNA damage | |||
| ENN A | <1 h | 1.5–3.0 | ROS generation | ||
| 24–72 h | IC50: 9.3–0.46 µM | ||||
| 24–72 h | 1.5–3.0 | LPO, loss of mitochondrial membrane potential, cell cycle arrest in SubG0/G1 and (Sub)G2/M, DNA damage, apoptosis and necrosis | |||
| ENN A1 | 10 min | 1.5 | ROS generation | ||
| 24–72 h | IC50: 12.3–0.46 µM | ||||
| 24–72 h | 1.5–3.0 | LPO, loss of mitochondrial membrane potential, DNA damage, cell cycle arrest in (Sub)G0/G1 and G2/M, apoptosis, necrosis | |||
| ENN B | 10 min | 3.0 | ROS generation | ||
| 48–72 h | IC50: 10.7–1.4 µM | ||||
| 24–72 h | 1.5–3.0 | LPO, loss of mitochondrial membrane potential, cell cycle arrest in (Sub)G0/G1, and G2/M, apoptosis, necrosis | |||
| ENN B1 | 5–10 min | 1.5–3.0 | ROS generation | ||
| 48–72 h | IC50: 10.8–0.8 µM | ||||
| 24–74 h | 1.5–3.0 | LPO, loss of mitochondrial membrane potential, DNA damage, cell cycle arrest in (Sub) G0/G1, G2/M and S, apoptosis, necrosis | |||
| MON | 72 h | IC50: 30.9 µg/mL | |||
| AOH | 24 h | 15–30 | changes in MMP f, ↓ G1 phase, ↑ S and G2/M phase, apoptosis, necrosis | ||
| HT-29 g | ENN A | 24–48 h | IC50: 9.3–8.2 µM | [56] | |
| ENN A1 | 24–48 h | IC50: 9.1–1.4 µM | |||
| ENN B | 24–48 h | IC50: ≥2.8 µM | |||
| ENN B1 | 24–48 h | IC50: 16.8–3.7 µM | |||
| HCT116 g | AOH | IC50, 24h: 65 µM ↓ early apoptotic and late apoptotic/necrotic cells, ROS generation PTP h-dependent MMP caspase-cascade activation, activation of p53 protein expression | [68,69] | ||
| AME | IC50, 24h: 120 µM apoptotic cell death, PTP-opening, induction of MMP, cytochrome c release caspase-cascade activation, ↑ p53 protein, ROS generation | ||||
| IPEC-J2 i | BEA | 24–72 h | 5–10 | TEERj reduction (between −59% and −80%), no reduction of cell viability | [59] |
| ENN A | 72 h | 5 | TEER reduction (−70%), no reduction of cell viability | ||
| ENN A1 | 24–72 h | 10 | TEER reduction (between −29% and −74%), no reduction of cell viability | ||
| ENN B | 48–72 h | 2.5 | TEER reduction (between −55% and −68%), no reduction of cell viability | ||
| ENN B1 | 48–72 h | 5 | TEER reduction (between −44% and −58%), no reduction of cell viability | ||
| ENN combinations | 1.5 | additive effect on TEER reduction | |||
| MON | 72 h | 5–10 | no effect on TEER or viability | [59] | |
| Hep-G2 k | ENN A | 24–48 h | IC50: 26.2–11.4 µM | [56,66] | |
| ENN A1 | 24–48 h | IC50: 11.6–2.6 µM | |||
| ENN B | 24–48 h | IC50: >30 µM | |||
| ENN B1 | 24–48 h | IC50: 24.3–8.5 µM | |||
| MON | 48–72 h | IC50: 39.5–24.1 µg/mL | |||
| H295R l | ENN B | 72 h | 10–100 | ↓ viability by 37%, ↑ S-phase, ↓ G0/G1phase, ↑ apoptosis ↓ HMGR, STAR, CYP11A, HSD3B2, CYP17A1 ↑ CYP1A1, MC2R, NR0B1, CYP21A2, CYP11B1, CYP19 ↓ progesterone, testosterone and cortisol; estradiol unaffected | [63,70,71] |
| AOH | 3.87 | no influence on viability ↑ 7 proteins (FDX1, HSD3B, CYP21A2, SCAMP3, SOAT1, ARF6, RRP15) ↓ 15 proteins (ACTBL2, NUCKS1, EIF2B5, COX2, CRMP1, ABHD14A-ACY1, ATP5J, ACSF2, HN1, ETHE1, HIST1H1E, ACBD5, NPC1, NR5A1, TOMM7) upregulation mRNA for CYP21A2 and HSD3B ↑ G0/G1 and ↑ G2/M phase | |||
| H29R l | AOH | no effect on testosterone and cortisol levels ↑ progesterone and estradiol levels ↓ NR0B1 gene ↑ CYP1A1, MC2R, HSD3B2, CYP17, CYP21, CYP11B2, CYP19 | [72] | ||
| neonatal Leydig cells | ENN B | 10–100 | ↓ viability by 20%, ↓ estradiol in unstimulated cells ↓ estradiol and testosterone in LH stimulated cells, probably due to cytotoxicity | [63] | |
| human breast adenocarcinoma RGA cell line | AOH | agonistic estrogen response, relative estrogenic potential: 0.0004% and equivalent estrogenic quantity of 17β-estradiol: 2.9 fg/mL | [72] | ||
| cell free buffer | AOH | binding affinity to ERα: 10,000× lower compared to 17β-estradiol binding affinity to ERβ: 2500× lower compared to 17β-estradiol similar EC50 | [73] | ||
| Ishikawa human endometrial adenocarcinoma cell line | AOH | 2.5–10 | ↑ alkaline phosphatase mRNA and activity ↓ G1 phase and ↑ S and G2/M phase ↓ cell number due to inhibition of proliferation | [73] | |
| porcine oocytes and embryos | BEA | >0.5 | ↓ rate of development of maturing oocyte and 2–4 cell stage embryo, activated oocytes and 2–4 cell stage embryos more sensitive than maturing oocytes, compromised cytoplasmic maturation and abnormal meiosis in oocytes, ↓ cumulus viability and progesterone synthesis, cumulus cells control intracellular BEA through MDR1 activity, in oocytes mitochondrial function was altered, altered gene expression in cumulus cells and oocytes, altered MDR1 activity in activated oocytes, ↓ viability embryo | [61] | |
| pig granulosa cells | AOH | 0.8–1.6 | ↓ cell viability, ↓ progesterone levels, ↓ P450scc ↓ α-tubulin, actin and EIF4a | [71] | |
| AME | 0.8–1.6 | ↓ cell viability, ↓ progesterone levels, ↓ P450scc | |||
| TeA | 6.4–100 | no influence on viability no influence on progesterone concentrations | |||
| bovine granulosa cells | BEA | 3 | ↓ estradiol and progesterone production ↓ CYP11A1 and CYP19A1 mRNA | [62] | |
| 6–10 | ↓ (fetal calf serum-induced) proliferation | ||||
| CHO-K1 m | BEA | 24–72 h | IC50: 10.7–2.2 µM combination of BEA + PAT n, BEA + STG o, BEA + PAT + STG: synergistic effect at low (IC < 1), additive effect at higher (IC 0.6–5.9) doses | [52,66,74] | |
| ENN A | 24–72 h | >7.5–2.83 µM | |||
| ENN A1 | 24–72 h | 8.8–1.65 µM | |||
| ENN B | 24–72 h | 11.0–2.44 µM | |||
| ENN B1 | 24–72 h | 4.53–2.47 µM | |||
| ENN combinations | 24 h | additive effects: A + B1, A1 + B, B + B1 synergistic effects: A + A1, A + B, A1 + B1, A1 + B1, A + A1 + B, A + A1 + B1, A1 + B + B1 (higher concentrations) antagonistic effects: A + A1 + B1, A1 + B +B1 (lower concentrations) | |||
| MON | IC50: >100 µg/mL | ||||
| THP-1 p monocyte | AOH | 24–48 h | 7.5–15 | cell cycle arrest in S- and G2/M-phase | [55] |
| ↓ CD14 and CD11b upregulation during macrophage differentiation | |||||
| ↓ downregulation of CD71 during macrophage differentiation, ↓ TNF-α secretion due to ↓ gene expression | |||||
| +DON: additive effect +ZEA: synergistic effect on macrophage differentiation | |||||
| CCRF-CEM q | BEA | 24 h | 1 | cytotoxicity, apoptosis | [54] |
| human lymphocytes | MON | 48 h | 10–25 15–25 | chromosome breaks, chromatid breaks and exchanges, polyploidy, increase in sister chromatid exchanges and micronuclei frequency all effects were dose-dependent | [8] |
| human immature dendritic cells | BEA | IC50: 1.0 µM | [53] | ||
| ENN B | IC50: 1.6 µM | ||||
| MON | 80 | 20% mortality, ↓ endocytosis, ↓ CD1a expression | |||
| human mature dendritic cells | BEA | IC50: 2.9 µM, ↓ CCR7 expression, ↑ IL-10 concentration | [53] | ||
| ENN B | IC50: 2.6 µM, ↓ CD80, CD86 and CCR7 expression, ↑ IL-10 | ||||
| MON | 80 | 20% mortality | |||
| human macrophages | BEA | ≥0.5 | IC50: 2.5 µM, ↓ endocytosis | [53,75] | |
| ENN B | IC50: 2.5 µM, ↓ endocytosis, ↑ CD71 | ||||
| MON | ↓ endocytosis, ↓ CD71, ↓ HLA-DR | ||||
| AOH | 24 h | 30 | changed morphology: from round to elongated with dendrite-like protrusions ↑ CD83 and CD86 ↓ HLA-DR and CD68 ↑ secretion of TNFα and IL-6 ↓ endocytosis and ↓ autophagy double DNA strand breaks | ||
| RAW 2654.7 mouse macrophage | AOH | 24–48 h | 30 | changed morphology: from round to flattened, star-shaped or elongated spindle-shaped cells micronuclei, polyploidy, ↑ CD86, CD80, MHCII (T cell activation), ↑ CD11b ↑ mRNA of TNFα and IL-6, but only ↑ TNFα secretion, ↑ endocytosis | [75] |
| mouse hemidiaphragm preparation | BEA | 5 | inhibition (in) directly elicited tetanic muscle contraction; inhibition nerve-evoked and directly elicited muscle twitches, reduction amplitude and frequency of miniature endplate potentials | [60] | |
| 1 h | 7.5 | inhibition directly elicited twitches, induction contracture, decrease resting membrane potential | |||
| 1 h | 10 | complete block of (in) directly elicited isometric muscle contraction, amplitude reduction of directly elicited muscle twitch, decrease resting membrane potential | |||
| C5-O r | MON | 72 h | IC50: 34.2 µg/mL | [66] | |
| V79 s | MON | 72 h | IC50: >100 µg/mL | [62,66,73] | |
| AOH | 5–50 | induction of micronuclei cell cycle arrest in G2 and S phase |
| Animal Species | Mycotoxin | Route of Exposure | Exposure Time | Exposure Dose | Effect | Reference |
|---|---|---|---|---|---|---|
| mouse | MON | po-ip | 1x | LD50: 20.9 (♀) 29.1 (♂) mg/kg bw (ip), survivors clinically healthy | [13,92] | |
| TeA | iv-po | 1x | 0–398 mg/kg bw | LD50: 76–162 (iv) and 81–209 (po), vomiting, diarrhea, hemorrhages, death | ||
| Sprague-Dawley rats | MON | po | 1x | 5 mg/kg bw | no clinical signs | [13,98,99] |
| 1x | 10 mg/kg bw | ↓ activity for 24 h, respiratory changes, trembling, piloerection, complete recovery within 48 h | ||||
| 1x | 25–50 mg/kg bw | respiratory and cardiovascular changes, collapse, convulsion and death within 48–83 min | ||||
| 28 days | 3–6 mg/kg bw | no clinical symptoms, no effect on leucocyte and red blood cell counts, food and water consumption or organ and body weights, ↓ phagocytic activity of neutrophils | ||||
| TeA | iv-po | 1x | 0–398 mg/kg bw | LD50: 83–157 (iv) and LD50: 168–240 (po), vomiting, diarrhea, hemorrhages, death | ||
| Syrian golden hamster | AME | ip | 1x | 200 mg/kg bw | severe necrosis and coalescence of visceral organs lethargy, breathing difficulties, flaccid hind limbs↑ resorptions and ↓ fetal weight | [100] |
| chicken embryo | MON | injection | 1x | LD50: 2.8 µg/egg, no gross teratogenic effects in survivors | [92] | |
| TeA | injection | 1x | 150–1500 µg/egg | dose-related mortality, LD50: 548 µg/egg | [101] | |
| 1-day old chicken | MON | po | 1x | 0–16 mg/kg bw | LD50: 5.4 mg/kg bw (crop intubation), survivors clinically healthy | [92] |
| broiler chickens | MON | feed | 21 days | 200 mg/kg feed | death (56%) ↑ kidney, heart and liver weight ↑ serum albumin, total protein and aspartate aminotransferase | [11,14,102] |
| 100 mg/kg feed | ↓ feed intake and body weight gain ↑ hearth weight ↑ kidney weight if feed also contained 200 mg FB1/kg ↑ incidence of large pleomorphic cardiomyocyte nuclei loss of cardiomyocyte cross striations mild focal renal tubular mineralization | |||||
| feed | 42 days (day 7–49) | 50 mg/kg feed | mortality (13.3%) ↑ feed consumption, ↓ body weight gain, ↓ feed conversion ↑ relative heart and proventriculus weight, ↓ mean corpuscular volume ↑ serum gamma glutamyltransferase activity loss of cardiomyocyte cross striations ↑ cardiomyocyte nuclear size | |||
| 25 mg/kg feed | mortality (7.8%) ↑ serum gamma glutamyltransferase activity | |||||
| TeA | po | 21 days | 1.25–2.5 mg/kg bw | ↓ weight gain and feed efficiency hemorrhages, erosions of the gizzard, pale mottled spleens, edema of the myocardium, microscopic congestions of blood vessels and hemorrhages | ||
| White leghorn chicken | TeA | po | 1x | LD50: 37.5 mg/kg bw with hemorrhages of the musculature of the thigh, breast, heart and subcutaneous tissues | [14] | |
| 21 days | 0.63 mg/kg bw | pathological changes in spleen and gizzard but no extensive hemorrhages microscopic congestions of blood vessels and hemorrhages | ||||
| 21 days | 1.25–2.5 mg/kg bw | ↓ weight gain and feed efficiency hemorrhages, erosions of the gizzard, pale mottled spleens, edema of the myocardium, microscopic congestions of blood vessels and hemorrhages | ||||
| turkeys | MON | feed | 91 days (day 7–98) | 25, 37.5, 50 mg/kg feed | ↑ relative heart weight | [102] |
| 37.5, 50 mg/kg feed | ↑ relative liver weight | |||||
| 50 mg/kg feed | loss of cardiomyocyte cross striations ↑ cardiomyocyte nuclear size, ↑ number of cardiomyocyte mitotic figures | |||||
| turkey poults | MON | feed | 21–28 days | 100 mg/kg feed | ↓ feed intake, body weight gain, feed efficiency, ↓ relative thymus, bursa and spleen weights, ↓ primary and secondary antibody response to inactivated Newcastle disease virus, ↓ systemic clearance of E. coli | [103] |
| Japanese quail | MON | feed | 35 days | 100 mg/kg feed | cardiomegaly, myocardial congestion, hypertrophy, myocardial disarray, ↑ mitochondria, resulting in separation of muscle fibers, swollen and deformed mitochondria with degenerative changes. Congestion, hemorrhages and degenerative changes more pronounced and extensive disruption of muscle fibers and destruction of Z-bands when feed contained both MON and fumonisin B1. Death. | [12] |
| barrow | MON | feed | 28 days | 100 mg/kg feed | acute mortality due to apparent cardiac failure ↓ body weight gain ↓ body weight gain, feed consumption and feed efficiency when feed also contained FB1 | [104] |
| dog | TeA | iv-po | 1x | 25–50 mg/kg bw | severe tachycardia, massive diffuse hemorrhages, (bloody) diarrhea | [13] |
| iv | 3x | 20 mg/kg bw | severe hemorrhagic gastro-enteropathy, death | |||
| iv-po | 6–30 days | 0.0625–11.2 mg/kg bw | salivation, emesis, tachycardia, hemorrhagic gastro-enteropathy, death | |||
| monkey | TeA | iv | 3x | 20 mg/kg bw | severe hemorrhagic gastro-enteropathy, death | [13] |
| iv-po | 7–45 days | 11.2–89.6 mg/kg | salivation, emesis, hemorrhagic gastro-enteropathy, death |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins 2017, 9, 228. https://doi.org/10.3390/toxins9070228
Fraeyman S, Croubels S, Devreese M, Antonissen G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins. 2017; 9(7):228. https://doi.org/10.3390/toxins9070228
Chicago/Turabian StyleFraeyman, Sophie, Siska Croubels, Mathias Devreese, and Gunther Antonissen. 2017. "Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics" Toxins 9, no. 7: 228. https://doi.org/10.3390/toxins9070228