Ricin Trafficking in Plant and Mammalian Cells

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Ricin Trafficking In Plant Cells
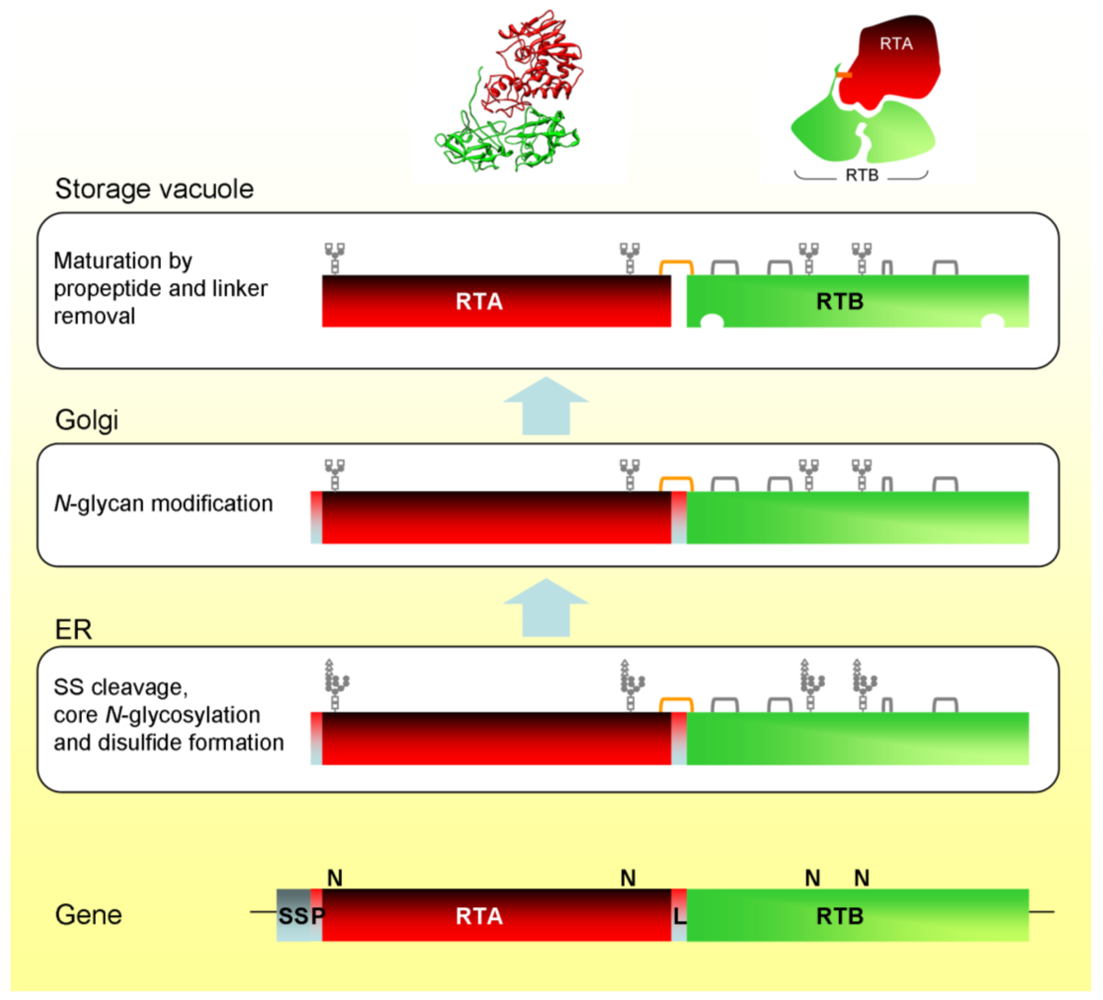
2.1. Ricin Biosynthesis
2.2. Synthesis and Endoplasmic Reticulum Translocation
2.3. Anterograde Trafficking in the Plant Cell

3. Ricin Trafficking in Mammalian Cells
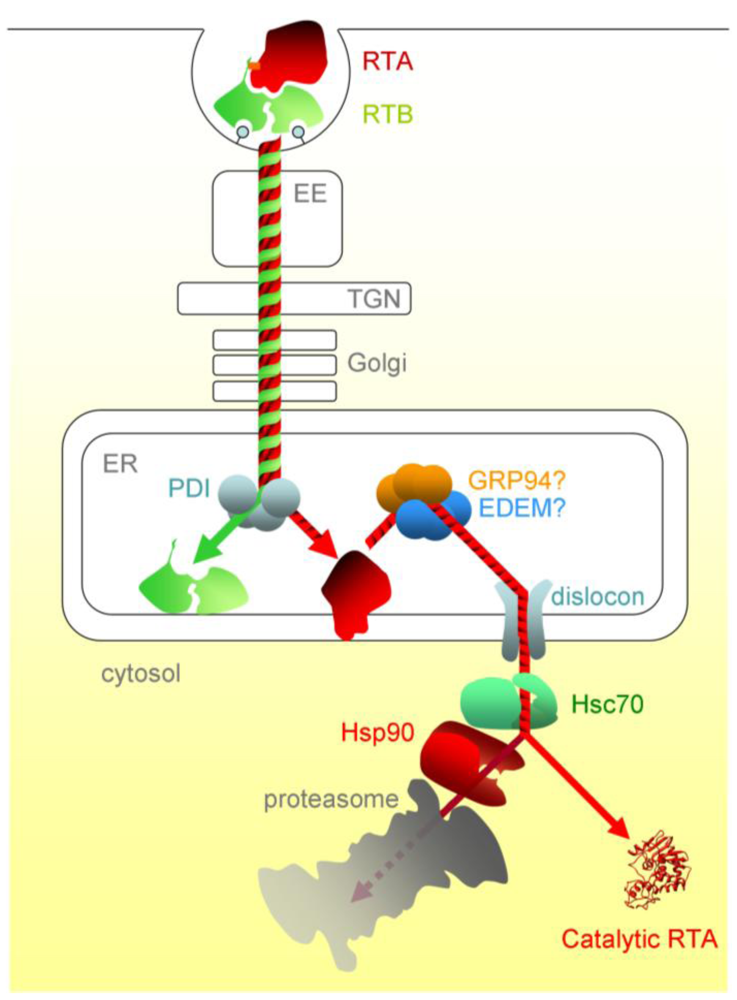
3.1. Ricin Entry into Mammalian Cells
3.2. Pre-Dislocation Events in the ER
3.3. Dislocation from the ER
3.4. Post-Dislocation Events in the Cytosol

4. Conclusions
References
- Stillmark, H. Uber ricin, eines gifiges ferment aus den samen von Ricinus communis L. und anderen euphorbiacen. Ph.D. Thesis, 1888. [Google Scholar]
- Nicolson, G.; Blaustein, J.; Etzler, M. Characterization of two plant lectins from Ricinus communis and their quantitative interaction with a murine lymphoma. Biochemistry 1974, 13, 196–204. [Google Scholar]
- Olsnes, S. Ricin and Ricinus agglutinin, toxic lectins from castor bean. Methods Enzymol. 1978, 50, 330–335. [Google Scholar]
- Endo, Y.; Tsurugi, K. RNA N-glycosidase activity of ricin A-chain. Mechanism of action of the toxic lectin ricin on eukaryotic ribosomes. J. Biol. Chem. 1987, 262, 8128–8130. [Google Scholar] [PubMed]
- Moazed, D.; Robertson, J.M.; Noller, H.F. Interaction of elongation factors EF-G and EF-Tu with a conserved loop in 23S RNA. Nature 1988, 334, 362–364. [Google Scholar]
- Lord, J.M.; Roberts, L.M.; Robertus, J.D. Ricin: Structure, mode of action, and some current applications. Fed. Am. Soc. Exp. Biol. 1994, 8, 201–208. [Google Scholar]
- Olsnes, S. A history of ricin, abrin and related toxins. Toxicon 2004, 44, 361–370. [Google Scholar]
- Audi, J.; Belson, M.; Patel, M.; Schier, J.; Osterloh, J. Ricin poisoning: A comprehensive review. J. Am. Med. Assoc. 2005, 294, 2342–2351. [Google Scholar]
- Lamb, F.I.; Roberts, L.M.; Lord, J.M. Nucleotide sequence of cloned cDNA coding for preproricin. Eur. J. Biochem. 1985, 148, 265–270. [Google Scholar]
- Roberts, L.M.; Lamb, F.I.; Pappin, D.J.; Lord, J.M. The primary sequence of Ricinus communis agglutinin. Comparison with ricin. J. Biol. Chem. 1985, 260, 15682–15686. [Google Scholar] [PubMed]
- Marshall, R.; Frigerio, L.; Roberts, L. Disulfide formation in plant storage vacuoles permits assembly of a mulimeric lectin. Biochem. J. 2010, 427, 513–521. [Google Scholar]
- Sweeney, E.C.; Tonevitsky, A.G.; Temiakov, D.E.; Agapov, II; Saward, S.; Palmer, R.A. Preliminary crystallographic characterization of ricin agglutinin. Proteins 1997, 28, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Tulley, R.E.; Beevers, H. Protein bodies of castor bean endosperm. Isolation, fractionation, and characterization of protein components. Plant Physiol. 1976, 58, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Huang, A.H. Protein bodies from castor bean endosperm. Subfractionation, protein components, lectins, and changes during germination. Plant Physiol. 1976, 58, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; Hao, Q.; Van Damme, E.J. Ribosome-inactivating proteins from plants: More than RNA N-glycosidases? FASEB J. 2001, 15, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Halling, K.C.; Halling, A.C.; Murray, E.E.; Ladin, B.F.; Houston, L.L.; Weaver, R.F. Genomic cloning and characterization of a ricin gene from Ricinus communis. Nucl. Acids Res. 1985, 13, 8019–8033. [Google Scholar]
- Tregear, J.W.; Roberts, L.M. The lectin gene family of Ricinus communis: Cloning of a functional ricin gene and three lectin pseudogenes. Plant Mol. Biol. 1992, 18, 515–525. [Google Scholar]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus commun. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Ferrini, J.B.; Martin, M.; Taupiac, M.P.; Beaumelle, B. Expression of functional ricin B chain using the baculovirus system. Eur. J. Biochem. 1995, 233, 772–777. [Google Scholar]
- Frigerio, L.; Jolliffe, N.A.; di Cola, A.; Felipe, D.H.; Paris, N.; Neuhaus, J.M.; Lord, J.M.; Ceriotti, A.; Roberts, L.M. The internal propeptide of the ricin precursor carries a sequence-specific determinant for vacuolar sorting. Plant Physiol. 2001, 126, 167–175. [Google Scholar]
- Lord, J.M. Precursors of ricin and Ricinus communis agglutinin. Glycosylation and processing during synthesis and intracellular transport. Eur. J. Biochem. 1985, 146, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Jolliffe, N.A.; di Cola, A.; Marsden, C.J.; Lord, J.M.; Ceriotti, A.; Frigerio, L.; Roberts, L.M. The N-terminal ricin propeptide influences the fate of ricin A chain in tobacco protoplasts. J. Biol. Chem. 2006, 281, 23377–23385. [Google Scholar]
- Harley, S.M.; Beevers, H. Ricin inhibition of in vivo protein synthesis by plant ribosomes. Proc. Natl. Acad. Sci. USA 1982, 79, 5935–5938. [Google Scholar] [CrossRef]
- Richardson, P.T.; Westby, M.; Roberts, L.M.; Gould, J.H.; Colman, A.; Lord, J.M. Recombinant proricin binds galactose but does not depurinate 28 S ribosomal RNA. FEBS Lett. 1989, 255, 15–20. [Google Scholar]
- Wright, H.T.; Robertus, J.D. The intersubunit disulfide bridge of ricin is essential for cytotoxicity. Arch. Biochem. Biophys. 1987, 256, 280–284. [Google Scholar]
- Lord, J.M.; Harley, S.M. Ricinus communis agglutinin B chain contains a fucosylated oligosaccharide side chain not present on ricin B chain. FEBS Lett. 1985, 189, 72–76. [Google Scholar]
- Hara-Nishimura, I.; Inoue, K.; Nishimura, M. A unique vacuolar processing enzyme responsible for conversion of several proprotein precursors into the mature forms. FEBS Lett. 1991, 294, 89–93. [Google Scholar]
- Hiraiwa, N.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. An aspartic endopeptidase is involved in the breakdown of propeptides of storage proteins in protein-storage vacuoles of plants. Eur. J. Biochem. 1997, 246, 133–141. [Google Scholar]
- Shimada, T.; Yamada, K.; Kataoka, M.; Nakaune, S.; Koumoto, Y.; Kuroyanagi, M.; Tabata, S.; Kato, T.; Shinozaki, K.; Seki, M.; et al. Vacuolar processing enzymes are essential for proper processing of seed storage proteins in Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 32292–32299. [Google Scholar] [PubMed]
- Hara-Nishimura, I. Vacuolar processing enzymes responsible for regulation of vacuolar function of plants. Tanpakushitsu Kakusan Koso 1997, 42, 2335–2341. [Google Scholar] [PubMed]
- Lord, J.M.; Harley, S.M. In vitro endoproteolytic cleavage of castor bean lectin precursors. Plant Sci. 1985, 41, 111–115. [Google Scholar]
- Frigerio, L.; Vitale, A.; Lord, J.M.; Ceriotti, A.; Roberts, L.M. Free ricin A chain, proricin, and native toxin have different cellular fates when expressed in tobacco protoplasts. J. Biol. Chem. 1998, 273, 14194–14199. [Google Scholar]
- Matsuoka, K.; Nakamura, K. Large alkyl side-chains of isoleucine and leucine in the NPIRL region constitute the core of the vacuolar sorting determinant of sporamin precursor. Plant Mol. Biol. 1991, 41, 825–835. [Google Scholar]
- Holwerda, B.; Padgett, H.; Rogers, J. Proaleurain vacuolar targeting is mediated by short contiguous peptide interactions. Plant Cell. 1992, 4, 307–318. [Google Scholar]
- Lord, J.; Roberts, L.; Lencer, W. Entry of protein toxins into mammalian cells by crossing the endoplasmic reticulum membrane: Co-opting basic mechanisms of endoplasmic reticulum-associated degradation. Curr. Top. Microbiol. Immunol. 2006, 300, 149–168. [Google Scholar]
- Johnson, A.E.; van Maes, M.A. The translocon: A dynamic gateway at the ER membrane. Annu. Rev. Cell Dev. Biol. 1999, 15, 799–842. [Google Scholar]
- Romisch, K. Endoplasmic reticulum-associated degradation. Annu. Rev. Cell Dev. Biol. 2005, 21, 435–456. [Google Scholar]
- Vembar, S.; Brodsky, J. One step at a time: Endoplasmic reticulum-associated degradation. Nat. Rev. Mol. Cell Biol. 2008, 9, 944–957. [Google Scholar]
- Carvalho, P.; Goder, V.; Rapoport, T. Distinct ubiquitin-ligase complexes define convergent pathways for the degradation of ER proteins. Cell 2006, 126, 361–373. [Google Scholar]
- Denic, V.; Quan, E.; Wisemen, J. A luminal surveillance complex that selects misfolded glycoproteins for ER-associated degradation. Cell 2006, 126, 349–359. [Google Scholar]
- Bays, N.W.; Gardner, R.G.; Seelig, L.P.; Joazeiro, C.A.; Hampton, R.Y. Hrd1p/Der3p is a membrane-anchored ubiquitin ligase required for ER-associated degradation. Nat. Cell. Biol. 2001, 3, 24–29. [Google Scholar]
- Jarosch, E.; Geiss-Friedlander, R.; Meusser, B.; Walter, J.; Sommer, T. Protein dislocation from the endoplasmic reticulum—pulling out the suspect. Traffic 2002, 3, 530–536. [Google Scholar]
- Rabinovich, E.; Kerem, A.; Frohlich, K.-U.; Diamant, N.; Bar-Nun, S. AAA-ATPase p97/Cdc48p, a cytosolic chaperone required of endoplasmic reticulum-associated protein degradation. Mol. Cell. Biol. 2002, 22, 626–634. [Google Scholar]
- Ye, Y.; Meyer, H.H.; Rapoport, T.A. The AAA ATPase Cdc48/p97 and its partners transport proteins from the ER into the cytosol. Nature 2001, 414, 652–656. [Google Scholar]
- Elkabetz, Y.; Shapira, H.; Rabinovich, E.; Bar-Nun, S. Distinct steps in the dislocation of luminal endoplasmic reticulum-associated degradation substrates. Roles of endoplasmic reticulum-boundp97/Cdc48p and proteasome. J. Biol. Chem. 2004, 279, 3980–3989. [Google Scholar] [PubMed]
- Lipson, C.; Alalouf, G.; Bajorek, M.; Rabinovich, E.; Atir-Lande, A.; Glickman, M.; Bar-Nun, S. A proteasomal ATPase contributes to dislocation of endoplasmic reticulum-associated degradation ERAD, substrates. J. Biol. Chem. 2008, 283, 7166–7175. [Google Scholar]
- Surjit, M.; Jameel, S.; Lal, S. Cytoplasmic location of the ORF2 protein of hepatitis E virus is dependent on its ability to undergo retrotranslocation from the endoplasmic reticulum. J. Virol. 2007, 81, 3339–3345. [Google Scholar]
- Giodini, A.; Cresswell, P. Hsp90-mediated cytosolic refoldingof exogenous proteins internalised by dendritic cells. EMBO J. 2008, 27, 201–211. [Google Scholar]
- Afshar, N.; Black, B.E.; Paschal, B.M. Retrotranslocation of the chaperone calreticulin from the endoplasmic reticulum lumen to the cytosol. Mol. Cell. Biol. 2005, 25, 8844–8853. [Google Scholar]
- Sandvig, K.; Olsnes, S.; Pihl, A. Kinetics of binding of the toxic lectins abrin and ricin to surface receptors of human cells. J. Biol. Chem. 1976, 251, 3977–3984. [Google Scholar]
- van Deurs, B.; Sandvig, K.; Petersen, O.W.; Olsnes, S.; Simons, K.; Griffiths, G. Estimation of the amount of internalized ricin that reaches the trans-Golgi network. J. Cell. Biol. 1988, 106, 253–267. [Google Scholar]
- Spooner, R.A.; Smith, D.C.; Easton, A.J.; Roberts, L.M.; Lord, J.M. Retrograde transport pathways utilised by viruses and protein toxins. Virol. J. 2006, 7, 26. [Google Scholar]
- Johannes, L.; Popoff, V. Tracing the retrograde route in protein trafficking. Cell 2008, 135, 1175–1187. [Google Scholar]
- Johannes, L.; Romer, W. Shiga toxins-from cell biology to medical applications. Nat. Rev. Microbiol. 2010, 8, 105–116. [Google Scholar]
- Boal, F.; Geutzoyan, L.; Session, R.; Zeghouf, M.; Spooner, R.; Lord, J.; Cherfils, J.; Clarkson, G.; Roberts, L.; Stephens, D. LG186: An inhibitor of GBF1 function that causes Golgi disassembly in human and canine cells. Traffic 2010, 11, 1537–1551. [Google Scholar]
- Spooner, R.A.; Watson, P.; Smith, D.C.; Boal, F.; Amessou, M.; Johannes, L.; Clarkson, G.J.; Lord, J.M.; Stephens, D.J.; Roberts, L.M. The secretion inhibitor Exo2 perturbs trafficking of Shiga toxin between endosomes and the trans-Golgi network. Biochem. J. 2008, 414, 471–484. [Google Scholar]
- Feng, Y.; Jadhav, A.; Rodighiero, C.; Fujinaga, Y.; Kirchausen, T.; Lencer, W. Retrograde transport of cholera toxin from the plasma membrane to the endoplasmic reticulum requires the trans-Golgi network but not the Golgi apparatus in Exo2-treated cells. EMBO Rep. 2004, 5, 596–601. [Google Scholar]
- Popoff, V.; Mardones, G.; Bai, S.; Chambon, V.; Tenza, D.; Burgos, P.; Shi, A.; Benaroch, P.; Urbe, S.; Lamaze, C.; et al. Analysis of articulation between clathrin and retromer in retrograde sorting on early endosomes. Traffic 2009, 10, 1868–1880. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.; Poupon, V.; Blondeau, F.; McPherson, P. The DnaJ-domain protein RME-8 functions in endosomal trafficking. J. Biol. Chem. 2005, 280, 40135–40143. [Google Scholar]
- Spooner, R.A.; Watson, P.D.; Marsden, C.J.; Smith, D.C.; Moore, K.A.; Cook, J.P.; Lord, J.M.; Roberts, L.M. Protein disulphide isomerase reduces ricin to its A and B chains in the endoplasmic reticulum. Biochem J. 2004, 383, 285–293. [Google Scholar]
- Bellisola, G.; Fracasso, G.; Ippoliti, R.; Menestrina, G.; Rosen, A.; Solda, S.; Udali, S.; Tomazzolli, R.; Tridente, G.; Colombatti, M. Reductive activation of ricin and ricin A-chain immunotoxins by protein disulfide isomerase and thioredoxin reductase. Biochem. Pharmacol. 2004, 67, 1721–1731. [Google Scholar]
- Majoul, I.; Ferrari, D.; Soling, H.D. Reduction of protein disulfide bonds in an oxidizing environment. The disulfide bridge of cholera toxin A-subunit is reduced in the endoplasmic reticulum. FEBS Lett. 1997, 401, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Hazes, B.; Read, R.J. Accumulating evidence suggests that several AB-toxins subvert the endoplasmic reticulum-associated protein degradation pathway to enter target cells. Biochemistry 1997, 36, 11051–11054. [Google Scholar]
- Deeks, E.D.; Cook, J.P.; Day, P.J.; Smith, D.C.; Roberts, L.M.; Lord, J.M. The low lysine content of ricin A chain reduces the risk of proteolytic degradation after translocation from the endoplasmic reticulum to the cytosol. Biochemistry 2002, 41, 3405–3413. [Google Scholar]
- Rodighiero, C.; Tsai, B.; Rapoport, T.A.; Lencer, W.I. Role of ubiquitination in retro-translocation of cholera toxin and escape of cytosolic degradation. EMBO Rep. 2002, 3, 1222–1227. [Google Scholar]
- Day, P.J.; Pinheiro, T.J.; Roberts, L.M.; Lord, J.M. Binding of ricin A-chain to negatively charged phospholipid vesicles leads to protein structural changes and destabilizes the lipid bilayer. Biochemistry 2002, 41, 2836–2843. [Google Scholar]
- Mayerhofer, P.U.; Cook, J.C.; Wahlman, J.; Pinheiro, T.T.; Moore, K.A.H.; Lord, J.M.; Johnson, A.E.; Roberts, L.M. Ricin A chain insertion into endoplasmic reticulum membranes is triggered by a temperature increase to 37 °C. J. Biol. Chem. 2009, 284, 10232–10242. [Google Scholar]
- Argent, R.H.; Parrott, A.M.; Day, P.J.; Roberts, L.M.; Stockley, P.G.; Lord, J.M.; Radford, S.E. Ribosome-mediated folding of partially unfolded ricin A-chain. J. Biol. Chem. 2000, 275, 9263–9269. [Google Scholar]
- Spooner, R.A.; Hart, P.J.; Cook, J.C.; Pietroni, P.; Rogon, C.; Hoehfeld, J.; Roberts, L.M.; Lord, J.M. Cytosolic chaperones influence the fate of a toxin dislocated from the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2008, 105, 17408–17413. [Google Scholar]
- Li, S.; Spooner, R.A.; Allen, S.C.H.; Guise, C.P.; Ladds, G.; Schnoeder, T.; Schmitt, M.J.; Lord, J.M.; Roberts, L.M. Folding-competent and folding-defective forms of ricin A chain have different fates after retrotranslocation from the endoplasmic reticulum. Mol. Biol. Cell. 2010, 21, 2543–2554. [Google Scholar]
- Slominska-Wojewodzka, M.; Gregers, T.; Walchli, S.; Sandvig, K. EDEM is involved in retrotranslocation of ricin from the endoplasmic reticulum to the cytosol. Mol. Biol. Cell. 2006, 17, 1664–1675. [Google Scholar]
- LaPointe, P.; Wei, X.; Gariepy, J. A role for the protease-sensitive loop region of Shiga-like toxin 1 in the retrotranslocation of its A1 domain from the endoplasmic reticulum lumen. J. Biol. Chem. 2005, 280, 23310–23318. [Google Scholar]
- Menikh, A.; Saleh, M.; Gariepy, J.; Boggs, J. Orientation in lipid bilayers of a synthetic peptide representing the C-terminus of the A1 domain of Shiga toxin. Biochemistry 1997, 36, 15865–15872. [Google Scholar]
- Saleh, M.T.; Ferguson, J.; Boggs, J.M.; Gariepy, J. Insertion and orientation of a synthetic peptide representing the C-terminus of the A1 domain of Shiga toxin into phospholipid membranes. Biochemistry 1996, 35, 9325–9334. [Google Scholar]
- Marshall, R.S.; Jolliffe, N.A.; Ceriotti, A.; Snowden, C.J.; Lord, J.M.; Frigerio, L.; Roberts, L.M. The role of CDC48 in the retro-translocation of non-ubiquitinated toxin substrates in plant cells. J. Biol. Chem. 2008, 283, 15869–15877. [Google Scholar]
- Suzuki, T.; Park, H.; Hollingsworth, N.M.; Sternglanz, R.; Lennarz, W.J. PNG1, a yeast gene encoding a highly conserved peptide: N-glycanase. J. Cell. Biol. 2000, 149, 1039–1052. [Google Scholar]
- Hosomi, A.; Tanabe, K.; Hirayama, H.; Kim, I.; Rao, H.; Suzuki, T. Identification of an Htm1 EDEM,-dependent, Mns1-independent Endoplasmic Reticulum-associated Degradation ERAD, pathway in Saccharomyces cerevisiae: Application of a novel assay for glycoprotein ERAD. J. Biol. Chem. 2010, 285, 2424–2434. [Google Scholar]
- Di Cola, A.; Frigerio, L.; Lord, J.; Roberts, L.; Ceriotti, A. Endoplasmic reticulum-associated degradation of ricin A chain has unique and plant-specific features. Plant Physiol. 2005, 137, 287–296. [Google Scholar]
- Simpson, J.C.; Roberts, L.M.; Romisch, K.; Davey, J.; Wolf, D.H.; Lord, J.M. Ricin A chain utilises the endoplasmic reticulum-associated protein degradation pathway to enter the cytosol of yeast. FEBS Lett. 1999, 459, 80–84. [Google Scholar]
- Wesche, J.; Rapak, A.; Olsnes, S. Dependence of ricin toxicity on translocation of the toxin A-chain from the endoplasmic reticulum to the cytosol. J. Biol. Chem. 1999, 274, 34443–34449. [Google Scholar] [CrossRef] [PubMed]
- Redman, V.; Oresic, K.; Tortorella, L.L.; Cook, J.P.; Lord, M.; Tortorella, D. Dislocation of ricin toxin A chains in human cells utilizes selective cellular factors. J. Biol. Chem. 2011, 286, 21231–21238. [Google Scholar]
- Lilley, B.N.; Ploegh, H.L. A membrane protein required for dislocation of misfolded proteins from the ER. Nature 2004, 429, 834–840. [Google Scholar]
- Ye, Y.; Shibata, Y.; Yun, C.; Ron, D.; Rapoport, T.A. A membrane protein complex mediates retro-translocation from the ER lumen into the cytosol. Nature 2004, 429, 841–847. [Google Scholar]
- Saslowsky, D.; Cho, J.; Chinnapen, H.; Massol, R.; Chinnapen, D.-F.; Wagner, J.; De Luca, H.; Kam, W.; Paw, B.; Lencer, W. Intoxication of zebrafish and mammalian cells by cholera toxin depends on the flotillin/reggie protein but not Derlin-1 or -2. J. Clin. Invest. 2010, 120, 4399–4409. [Google Scholar]
- AbuJarour, R.J.; Dalal, S.; Hanson, P.I.; Draper, R.K. p97 s in a complex with cholera toxin and influences the transport of cholera toxin and related toxins to the cytoplasm. J. Biol. Chem. 2005, 280, 15865–15871. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lord, J.M.; Spooner, R.A. Ricin Trafficking in Plant and Mammalian Cells. Toxins 2011, 3, 787-801. https://doi.org/10.3390/toxins3070787
Lord JM, Spooner RA. Ricin Trafficking in Plant and Mammalian Cells. Toxins. 2011; 3(7):787-801. https://doi.org/10.3390/toxins3070787
Chicago/Turabian StyleLord, J. Michael, and Robert A. Spooner. 2011. "Ricin Trafficking in Plant and Mammalian Cells" Toxins 3, no. 7: 787-801. https://doi.org/10.3390/toxins3070787