Detection of stx1 and stx2 Genes in Pennsylvanian White-Tailed Deer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Processing
2.3. DNA Isolation
2.4. Quantitative PCR
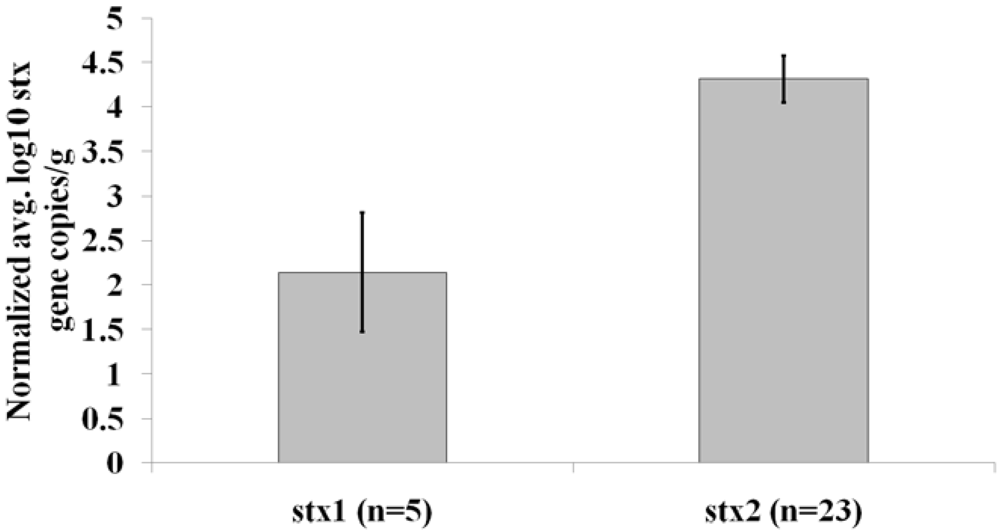
3. Results and Discussion

{kind=link}
| Gene Tested | Fraction (%) Positive for Gene |
|---|---|
| stx1 | 5/50 (10.0) |
| stx2 | 23/50 (46.0) |
| stx1 + stx2 | 5/50 (10.0) |
| stx1 or stx2 + eae | 4/50 (8.0) |

4. Conclusions
Acknowledgments
Conflict of Interest
- The authors declare no conflict of interest.
References
- Tyler, J.S.; Livny, J.; Friedman, D.I. Lambdoid Phages and Shiga Toxin. In Phages: Their Role in Bacterial Pathogenesis and Biotechnology; Waldor, M.K., Friedman, D.I., Adhya, S.L., Eds.; ASM Press: Washington, DC, USA, 2005; pp. 131–164. [Google Scholar]
- Ochoa, T.J.; Cleary, T.G. Epidemiology and spectrum of disease of Escherichia coli O157. Curr. Opin. Infect. Dis. 2003, 16, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Henderson, H. Direct and indirect zoonotic transmission of Shiga toxin-producing Escherichia coli. J. Am. Vet. Med. Assoc. 2008, 232, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States- major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar]
- Keene, W.E.; Sazie, E.; Kok, J.; Rice, D.H.; Hancock, D.D.; Balan, V.K.; Zhao, T.; Doyle, M.P. An outbreak of Escherichia coli O157:H7 infections traced to jerky made from deer meat. J. Am. Med. Assoc. 1997, 277, 1229–1231. [Google Scholar]
- Asakura, H.; Makino, S.; Shirahata, T.; Tsukamoto, T.; Kurazono, H.; Ikeda, T.; Takeshi, K. Detection and genetical characterization of Shiga toxin-producing Escherichia coli from wild deer. Microbiol. Immunol. 1998, 42, 815–822. [Google Scholar] [PubMed]
- Sánchez, S.; García-Sánchez, A.; Martínez, R.; Blanco, J.; Blanco, J.E.; Blanco, M.; Dahbi, G.; Mora, A.; Hermoso de Mendoza, J.; Alonso, J.M.; et al. Detection and characterisation of Shiga toxin-producing Escherichia coli other than Escherichia coli O157:H7 in wild ruminants. Vet. J. 2009, 180, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.R.; Zhao, T.; Doyle, M.P.; Goldberg, M.R.; Brown, C.A.; Sewell, C.T.; Kavanaugh, D.M.; Bauman, C.D. Experimental and field studies of Escherichia coli O157:H7 in white-tailed deer. Appl. Environ. Microbiol. 2001, 67, 1218–1224. [Google Scholar] [PubMed]
- Renter, D.G.; Sargeant, J.M.; Hygnstorm, S.E.; Hoffman, J.D.; Gillespie, J.R. Escherichia coli O157:H7 in free-ranging deer in Nebraska. J. Wildl. Dis. 2001, 37, 755–760. [Google Scholar] [PubMed]
- Dunn, J.R.; Keen, J.E.; Moreland, D.; Thompson, R.A. Prevalance of Escherichia coli O157:H7 in white-tailed deer from Louisiana. J. Wildl. Dis. 2004, 40, 361–365. [Google Scholar] [PubMed]
- Blanco, M.J.; Blanco, J.E.; Mora, A.; Dahbi, G.; Alonso, M.P.; Gonzalez, A.; Bernandez, M.I.; Blanco, J. Serotypes, virulence genes, and intimin types of Shiga toxin (verocytotoxin)-producing Escherichia coli isolates from cattle in Spain and identification of a new intimin gene (eae-xi). J. Clin. Microbiol. 2004, 42, 645–651. [Google Scholar] [PubMed]
- Schmidt, H.; Montag, M.; Bockemuhl, J.; Heesemann, J.; Karch, H. Shiga-like toxin II-related cytotoxins in Citrbacter Freundii strains from humans and beef samples. Infect. Immunol. 1993, 61, 534–543. [Google Scholar]
- Paton, A.W.; Paton, J.C. Enterobacter cloacae producing a Shiga-like toxin II-related cytotoxin associated with a case of hemolytic-uremic syndrome. J. Clin. Microbiol. 1996, 34, 463–465. [Google Scholar] [PubMed]
- Smith, C.J.; Olszewski, A.M.; Mauro, S.A. Comparison of Shiga toxin gene frequency to commonly used microbial indicators in recreational waters. Appl. Environ. Microbiol. 2009, 75, 316–321. [Google Scholar]
- Ibekwe, M.A.; Watt, P.M.; Grieve, C.M.; Sharma, V.K.; Lyons, S.R. Multiplex fluorogenic real-time pcr for detection and quantification of Escherichia O157:H7 in dairy wastewater wetlands. Appl. Environ. Microbiol. 2002, 68, 4853–4862. [Google Scholar] [CrossRef] [PubMed]
- Rogerie, F.; Marecat, A.; Gambade, S.; Duponda, F.; Beauboisb, P.; Lange, M. Characterization of Shiga toxin producing E. coli and O157 serotype E. coli isolated in France from healthy domestic cattle. Int. J. Food Microbiol. 2001, 63, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Pearce, M.C.; Chart, H.; Cheasty, T.; Willshaw, G.A.; Gunn, G.J.; Dougan, G.; Smith, H.R.; Synge, B.A.; Frankel, G. An eight-month study of a population of verocytotoxigenic Escherichia coli (VTEC) in a Scottish cattle herd. J. Appl. Microbiol. 2002, 93, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Gilbreath, J.J.; Shields, M.S.; Smith, R.L.; Farrell, L.D.; Sheridan, P.P.; Spiegel, K.M. Shiga toxins, and the genes encoding them, in fecal samples from native Idaho ungulates. Appl. Environ. Microbiol. 2009, 75, 862–865. [Google Scholar] [PubMed]
- Menrath, A.; Wieler, L.H.; Heidemanns, K.; Semmler, T.; Fruth, A.; Kemper, N. Shiga toxin producing Escherichia coli: identification of non-O157:H7-Super-Shedding cows and related risk factors. Gut Pathog. 2010, 2, 7. [Google Scholar] [PubMed]
- Cerqueira, A.M.F.; Guth, B.E.C.; Joaquim, R.M.; Andrade, J.R.C. High occurrence of Shiga toxin-producing Escherichia coli (STEC) in healthy cattle in Rio de Janeiro State, Brazil. Vet. Microbiol. 1999, 70, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.; Rodriguez, E.M.; Arroyo, G.H.; Padola, N.L.; Parma, A.E. Seasonal variation of Shiga toxin-encoding genes (stx) and detection of E. coli O157 in dairy cattle from Argentina. J. Appl. Microbiol. 2009, 106, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.D.; Sullivan, K.L. Wildlife Damage Management Fact Sheet Series White-Tailed Deer. Cornell Cooperative Extension, Wildlife Damage Management Program, Ithaca, NY, USA, 2001. Available online: http://wildlifecontrol.info/pubs/Documents/Deer/Deer_factsheet.pdf (accessed on 14 June 2011).
- Pennsylvania Game Commission Press Release Number 029-08. Available online: http://www.pgc.state.pa.us/pgc/cwp/view.asp?Q=173862&A=11 (accessed on 14 June 2011).
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kistler, W.M.; Mulugeta, S.; Mauro, S.A. Detection of stx1 and stx2 Genes in Pennsylvanian White-Tailed Deer. Toxins 2011, 3, 640-646. https://doi.org/10.3390/toxins3060640
Kistler WM, Mulugeta S, Mauro SA. Detection of stx1 and stx2 Genes in Pennsylvanian White-Tailed Deer. Toxins. 2011; 3(6):640-646. https://doi.org/10.3390/toxins3060640
Chicago/Turabian StyleKistler, Whitney M., Surafel Mulugeta, and Steven A. Mauro. 2011. "Detection of stx1 and stx2 Genes in Pennsylvanian White-Tailed Deer" Toxins 3, no. 6: 640-646. https://doi.org/10.3390/toxins3060640