New Analytical Approach to Quinolizidine Alkaloids and Their Assumed Biosynthesis Pathways in Lupin Seeds


 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Analytical Method
2.2. Quinolizidine Alkaloid Profiles
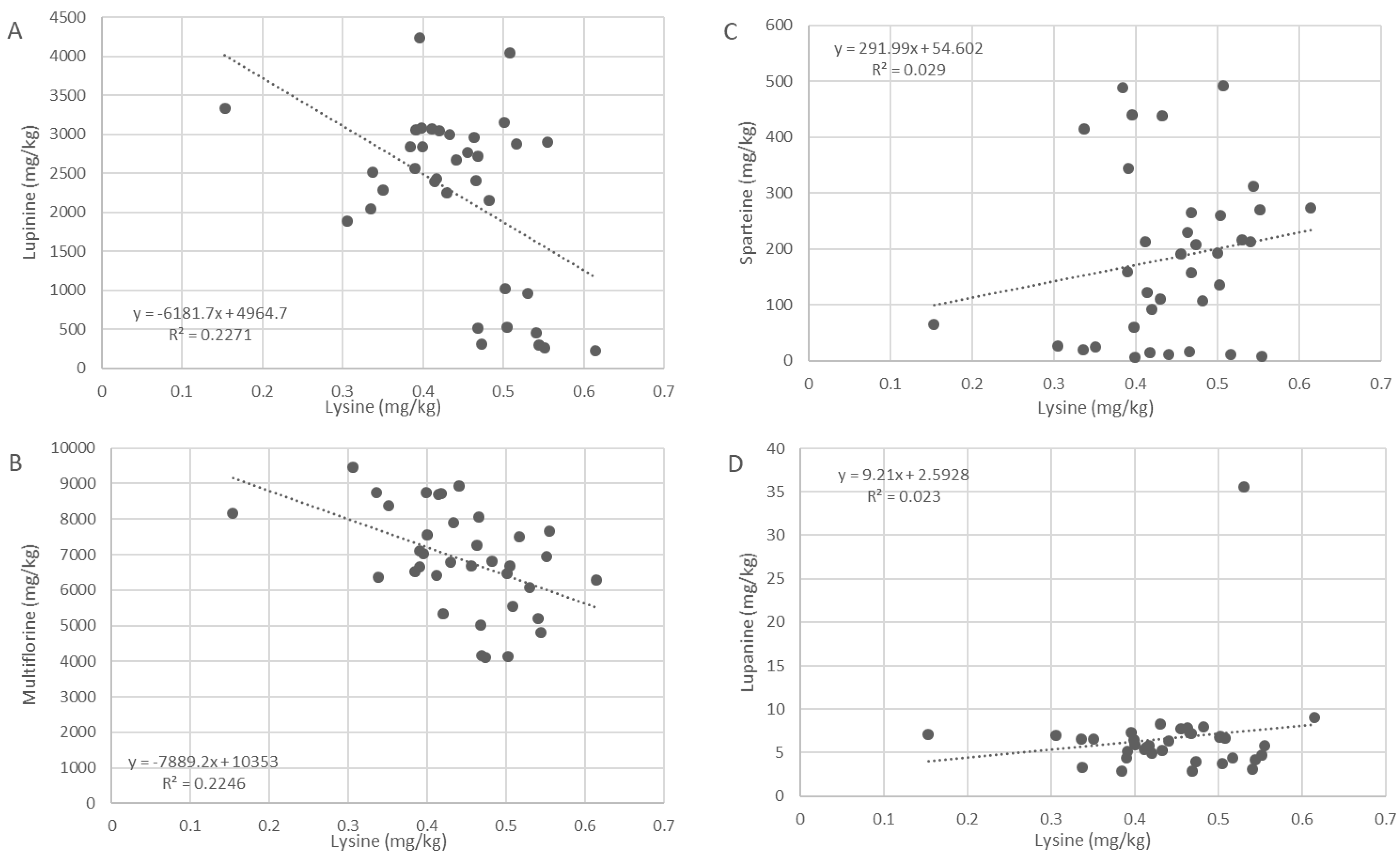
2.3. Correlation with L-Lysine
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Methods
5.2.1. Seed Collection Method
5.2.2. Analytical Standards
5.2.3. Sample Pretreatment
5.2.4. Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS)
5.2.5. Validation of the LC–MS/MS Method
5.2.6. Gas Chromatography Coupled with Mass Spectroscopy (GC–MS)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Dyer, L.A.; Philbin, C.S.; Ochsenrider, K.M.; Richards, L.A.; Massad, T.J.; Smilanich, A.M.; Forister, M.L.; Parchman, T.L.; Galland, L.M.; Hurtado, P.J.; et al. Modern approaches to study plant-insect interactions in chemical ecology. Nat. Rev. Chem. 2018, 2, 50–64. [Google Scholar] [CrossRef]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Ann. Rev. Plant Biol. 2012, 63, 431. [Google Scholar] [CrossRef]
- Adamski, Z.; Bufo, S.A.; Milella, L.; Scrano, L. Identification and Functional Characterization of Plant Toxins. Toxins 2021, 13, 228. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.(R.); Leblanc, J.; Nebbia, C.S.; et al. Scientific opinion on the risks for animal and human health related to the presence of quinolizidine alkaloids in feed and food, in particular in lupins and lupin-derived products. EFSA J. 2019, 17, e05860. [Google Scholar] [PubMed]
- Michael, J.P. The Alkaloids; Knölker, H.-J., Ed.; Academic Press: London, UK, 2016; Volume 75, pp. 1–498. [Google Scholar]
- Wink, M.; Witte, L. Cell-free synthesis of the alkaloids ammodendrine and smipine. Z. Für Naturforschung C 1987, 42, 197–204. [Google Scholar] [CrossRef]
- Boschin, G.; Resta, D. Alkaloids Derived from Lysine: Quinolizidine (a Focus on Lupin Alkaloids). In Natural Products; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Frick, K.M.; Kamphuis, L.G.; Siddique, K.H.M.; Singh, K.B.; Foley, R.C. Quinolizidine alkaloid biosynthesis in lupins and prospects for grain quality improvement. Front. Plant Sci. 2017, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Resta, D.; Boschin, G.; D’Agostina, A.; Arnoldi, A. Evaluation of total quinolizidine alkaloids content in lupin flours, lupin-based ingredients, and foods. Mol. Nutr. Food Res. 2008, 52, 490–495. [Google Scholar] [CrossRef]
- Boschin, G.; Tesio, E.; Arnoldi, A. A field case of pig poisoning by accidental feed contamination by alkaloid-rich lupin seeds. J. Appl. Anim. Res. 2022, 50, 725–731. [Google Scholar] [CrossRef]
- Yovo, K.; Huguet, F.; Pothier, J.; Durand, M.; Breteau, M.; Narcisse, G. Comparative Pharmacological Study of Sparteine and its Ketonic Derivative Lupanine from Seeds of Lupinus albus. Planta Medica 1984, 50, 420–424. [Google Scholar] [CrossRef]
- Otterbach, S.L.; Yang, T.; Kato, L.; Janfelt, C.; Geu-Flores, F. Quinolizidine alkaloids are transported to seeds of bitter narrow-leafed lupin. J. Exp. Bot. 2019, 70, 5799–5808. [Google Scholar] [CrossRef]
- Engel, A.M.; Engel, A.M.; Klevenhusen, F.; Klevenhusen, F.; Moenning, J.-L.; Moenning, J.-L.; Numata, J.; Numata, J.; Fischer-Tenhagen, C.; Fischer-Tenhagen, C.; et al. Investigations on the transfer of quinolizidine alkaloids from Lupinus angustifolius into the milk of dairy cows. J. Agric. Food Chem. 2022, 70, 11749–11758. [Google Scholar] [CrossRef]
- Mancinotti, D.; Frick, K.M.; Geu-Flores, F. Biosynthesis of quinolizidine alkaloids in lupins: Mechanistic considerations and prospects for pathway elucidation. Nat. Prod. Rep. 2022, 39, 1423. [Google Scholar] [CrossRef]
- Bundesinstitut für Risikobewertung. Risk Assessment of the Occurrence of Alkaloids in Lupin Seeds; BfR Opinion No 003/2017; 2017; Volume 1, pp. 1–35. Available online: https://mobil.bfr.bund.de/cm/349/risk-assessment-of-the-occurrence-of-alkaloids-in-lupin-seeds.pdf (accessed on 12 February 2024).
- Wink, M.; Meißner, C.; Witte, L. Patterns of quinolizidine alkaloids in 56 species of the genus Lupinus. Phytochemistry 1995, 38, 139–153. [Google Scholar] [CrossRef]
- Bunsupa, S.; Katayama, K.; Ikeura, E.; Oikawa, A.; Toyooka, K.; Saito, K.; Yamazaki, M. Lysine decarboxylase catalyzes the first step of quinolizidine alkaloid biosynthesis and coevolved with alkaloid production in leguminosae. Front. Plant Sci. 2012, 3, 239. [Google Scholar] [CrossRef]
- Cely-Veloza, W.; Quiroga, D.; Coy-Barrera, E. Quinolizidine-based variations and antifungal activity of eight Lupinus species grown under greenhouse conditions. Molecules 2022, 27, 305. [Google Scholar] [CrossRef] [PubMed]
- Shelef, O.; Weisberg, P.J.; Provenza, F.D. The value of native plants and local production in an era of global agriculture. Front. Plant Sci. 2017, 8, 2069. [Google Scholar] [CrossRef] [PubMed]
- Hernández-López, I.; Ortiz-Solà, J.; Alamprese, C.; Barros, L.; Shelef, O.; Basheer, L.; Rivera, A.; Abadias, M.; Aguiló-Aguayo, I. Valorization of local legumes and nuts as key components of the mediterranean diet. Foods 2022, 11, 3858. [Google Scholar] [CrossRef] [PubMed]
- Martiniello, P.; Teixeira da Silva, J.A. Physiological and bio-agronomical aspects involved in growth and yield components of cultivated forage species in Mediterranean environments: A review. Eur. J. Plant Sci. Biotech. 2011, 5, 64–98. [Google Scholar]
- Guilengue, N.A. Genetic and Morphological Diversity of Germplasm Collection from Lupinus mutabilis Sweet Grown under Mediterranean Climate. Ph.D. Thesis, Universidade de Lisboa, Lisbon, Portugal, 2021. [Google Scholar]
- Bermúdez-Torres, K.; Ferval, M.; Hernández-Sánchez, A.M.; Tei, A.; Gers, C.; Wink, M.; Legal, L. Molecular and chemical markers to illustrate the complex diversity of the genus Lupinus (Fabaceae). Diversity 2021, 13, 263. [Google Scholar] [CrossRef]
- Romeo, F.V.; Fabroni, S.; Ballistreri, G.; Muccilli, S.; Spina, A.; Rapisarda, P. Characterization and antimicrobial activity of alkaloid extracts from seeds of different genotypes of Lupinus spp. Sustainability 2018, 10, 788. [Google Scholar] [CrossRef]
- Święcicki, W.; Czepiel, K.; Wilczura, P.; Barzyk, P.; Kaczmarek, Z.; Kroc, M. Chromatographic Fingerprinting of the Old World Lupins Seed Alkaloids: A Supplemental Tool in Species Discrimination. Plants 2019, 8, 548. [Google Scholar] [CrossRef]
- Heistinger, A.; Pistrick, K. ‘Altreier Kaffee’: Lupinus pilosus L. cultivated as coffee substitute in northern Italy (Alto Adige/Südtirol). Genet. Resour. Crop Evol. 2007, 54, 1623–1630. [Google Scholar] [CrossRef]
- Carvajal-Larenas, F.E.; Linnemann, A.R.; Nout, M.J.R.; Koziol, M.; van Boekel, M.A.J.S. Lupinus mutabilis: Composition, uses, toxicology, and debittering. Crit. Rev. Food Sci. Nutr. 2016, 56, 1454–1487. [Google Scholar] [CrossRef]
- Kamel, K.A.; Święcicki, W.; Kaczmarek, Z.; Barzyk, P. Quantitative and qualitative content of alkaloids in seeds of a narrow-leafed lupin (Lupinus angustifolius L.) collection. Genet. Resour. Crop Evol. 2016, 63, 711–719. [Google Scholar] [CrossRef]
- Kroc, M.; Rybiński, W.; Wilczura, P.; Kamel, K.; Kaczmarek, Z.; Barzyk, P.; Święcicki, W. Quantitative and qualitative analysis of alkaloids composition in the seeds of a white lupin (Lupinus albus L.) collection. Genet. Resour. Crop Evol. 2017, 64, 1853–1860. [Google Scholar] [CrossRef]
- Eugelio, F.; Palmieri, S.; Fanti, F.; Messuri, L.; Pepe, A.; Compagnone, D.; Sergi, M. Development of an HPLC-MS/MS Method for the Determination of Alkaloids in Lupins. Molecules 2023, 28, 1531. [Google Scholar] [CrossRef]
- El-Shazly, A.; Ateya, A.M.M.; Wink, M. Quinolizidine Alkaloid Profiles of Lupinus varius orientalis, L. albus albus, L. hartwegii, and L. densiflorus. Z. Für Naturforschung C 2001, 56, 21–30. [Google Scholar] [CrossRef]
- Magalhães, S.C.; Fernandes, F.; Cabrita, A.R.; Fonseca, A.J.; Valentão, P.; Andrade, P.B. Alkaloids in the valorization of European Lupinus spp. seeds crop. Ind. Crop. Prod. 2017, 95, 286–295. [Google Scholar] [CrossRef]
- Ainouche, A.; Greinwald, R.; Witte, L.; Huon, A. Seed alkaloid composition of Lupinus tassilicus Maire (Fabaceae: Genisteae) and comparison with its related rough seeded lupin species. Biochem. Syst. Ecol. 1996, 24, 405–414. [Google Scholar] [CrossRef]
- Thomas, A.F.; Vipond, H.J.; Marion, L. The papilionaceous alkaloids: Xxi. The alkaloids of Lupinus pilosus walt. And the structure of tetralupine. Can. J. Chem. 1955, 33, 1290–1294. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.Q.; Fang, J. The biological activities of quinolizidine alkaloids. In The Alkaloids: Chemistry and Biology; Academic Press: Cambridge, MA, USA, 2023; Volume 89, pp. 1–37. [Google Scholar]
- Vishnyakova, M.A.; Kushnareva, A.V.; Shelenga, T.V.; Egorova, G.P. Alkaloids of narrow-leaved lupine as a factor determining alternative ways of the crop’s utilization and breeding. Vavilov J. Genet. Breed. 2020, 24, 625–635. [Google Scholar] [CrossRef]
- Gavilan, J.; Mennickent, D.; Ramirez-Molina, O.; Triviño, S.; Perez, C.; Silva-Grecchi, T.; Godoy, P.A.; Becerra, J.; Aguayo, L.G.; Moraga-Cid, G.; et al. 17 Oxo Sparteine and Lupanine, Obtained from Cytisus scoparius, Exert a Neuroprotection against Soluble Oligomers of Amyloid-β Toxicity by Nicotinic Acetylcholine Receptors. J. Alzh. Dis. 2019, 67, 343. [Google Scholar] [CrossRef]
- Lu, A.; Wang, T.; Hui, H.; Wei, X.; Chui, W.; Zhou, C.; Li, H.; Wang, Z.; Guo, J.; Ma, D.; et al. Natural products for drug discovery: Discovery of gramines as novel agents against a plant virus. J. Agric. Food Chem. 2019, 67, 2148. [Google Scholar] [CrossRef] [PubMed]
- Mironets, R.V.; Garazd, Y.L.; Garazd, M.M. Synthesis of Conjugates of the Alkaloids Cytisine and Lupinine. Chem. Nat. Compd. 2019, 55, 1115. [Google Scholar] [CrossRef]
- Tsutsumi, T.; Karanjit, S.; Nakayama, A.; Namba, K. A concise asymmetric total synthesis of (+)-Epilupinine. Org. Lett. 2019, 21, 2620. [Google Scholar] [CrossRef]
- Xue, S.; Xu, H.; Sun, Z.; Shen, H.; Chen, S.; Ouyang, J.; Zhou, Q.; Hu, X.; Cui, H. Depletion of TRDMT1 affects 5-methylcytosine modification of mRNA and inhibits HEK293 cell proliferation and migration. Biochemic. Biophysic. Res. Commun. 2019, 520, 60. [Google Scholar] [CrossRef]
- Lehner, Z.; Stadlbauer, K.; Brunmair, B.; Adorjan, I.; Genov, M.; Kautzky-Willer, A.; Scherer, T.; Scheinin, M.; Bauer, L.; Fürnsinn, C. Evidence that the multiflorine-derived substituted quinazolidine 55P0251 augments insulin secretion and lowers blood glucose via antagonism at α2-adrenoceptors in mice. Diabetes Obes. Metab. 2020, 22, 290. [Google Scholar] [CrossRef] [PubMed]
- Lucio FN, M.; da Silva, J.E.; Marinho, E.M.; Mendes FR, D.S.; Marinho, M.M.; Marinho, E.S. Methylcytisine alcaloid potentially active against dengue virus: A molecular docking study and electronic structural characterization. Int. J. Res. Gran. 2020, 8, 221. [Google Scholar] [CrossRef]
- Mazumder, K.; Nabila, A.; Aktar, A.; Farahnaky, A. Bioactive variability and in vitro and in vivo antioxidant activity of unprocessed and processed flour of nine cultivars of Australian lupin species: A comprehensive substantiation. Antioxidants 2020, 9, 282. [Google Scholar] [CrossRef]
- Nurmaganbetov, Z.S.; Savelyev, V.A.; Gatilov, Y.V.; Nurkenov, O.A.; Seidakhmetova, R.B.; Shulgau, Z.T.; Mukusheva, G.K.; Fazylov, S.D.; Shults, E.E. Synthesis and analgesic activity of 1-[(1, 2, 3-triazol-1-yl) methyl] quinolizines based on the alkaloid lupinine. Chem. Heterocycl. Compd. 2021, 57, 977. [Google Scholar] [CrossRef]
- Lu, X.; Lu, F.; Yu, J.; Xue, X.; Jiang, H.; Jiang, L.; Yang, Y. Gramine promotes functional recovery after spinal cord injury via ameliorating microglia activation. J. Cell. Mol. Med. 2021, 25, 7980. [Google Scholar] [CrossRef]
- Petruczynik, A.; Wróblewski, K.; Misiurek, J.; Plech, T.; Szalast, K.; Wojtanowski, K.; Mroczek, T.; Szymczak, G.; Waksmundzka-Hajnos, M.; Tutka, P. Determination of Cytisine and N-Methylcytisine from Selected Plant Extracts by High-Performance Liquid Chromatography and Comparison of Their Cytotoxic Activity. Toxins 2020, 12, 557. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.H.; Elissawy, A.M.; Eldahshan, O.A.; Elshanawany, M.A.; Singab, A.N.B. Variability of the Chemical Composition of the Essential Oils of Flowers and the Alkaloid Contents of Leaves of Sophora secundiflora and Sophora tomentosa. J. Essent. Oil Bear. Plants 2020, 23, 442–452. [Google Scholar] [CrossRef]
- Kushnareva, A.V.; Shelenga, V.; Perchuk, I.N.; Egorova, G.P.; Malyshev, L.L.; Kerv, Y.A.; Shavarda, A.L.; Vishnyakova, M.A. Selection of an optimal method for screening the collection of narrow-leaved lupine held by the Vavilov Institute for the qualitative and quantitative composition of seed alkaloids. Vavilov J. Genet. Breed. 2020, 24, 829. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, M.; Hummert, C.; Luckas, B. Hydrolysis of microcystins and nodularin by microwave radiation. Chromatographia 1999, 49, 671–677. [Google Scholar] [CrossRef]
- Li, N.; Liu, Y.; Zhao, Y.; Zheng, X.; Lu, J.; Liang, Y. Simultaneous HPLC determination of amino acids in tea infusion coupled to pre-column derivatization with 2, 4-dinitrofluorobenzene. Food Anal. Met. 2016, 9, 1307–1314. [Google Scholar] [CrossRef]
- Ogle, D.H.; Doll, J.C.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package. 2021. Available online: https://CRAN.R-project.org/package=FSA (accessed on 12 February 2024).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0. 12. 2019. 2019. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 12 February 2024).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Regional Distribution | Population Code | Collection Site/Notes |
|---|---|---|---|
| Golan Heights | PIL-1 | Mapalim Junction | |
| Hula Valley | PIL-2 | Nahal Hamdal | |
| Judean Hills | PIL-3 | Tel Socho South | |
| Golan Heights | PIL-4 | Ofir Viewpoint | |
| Golan Heights | PIL-5 | South-west Hispin | |
| Golan Heights | PIL-10 | Hazeka Road | |
| Judean Hills | PIL-11 | Matta | |
| Golan Heights | PIL-13 | Fares Road (vineyard) | |
| Golan Heights | PIL-14 | Tel Fazra | |
| Lupinus pilosus | Judean Hills | PIL-18 | Khirbet Kanim |
| Judean Hills | PIL-19 | Zechariya (Tel e-Sharia) | |
| Golan Heights | PIL-20 | Avital | |
| Carmel Mountain | PIL-21 | Makura | |
| Samarea Mountains | PIL-23 | Awartha | |
| Samarea Mountains | PL-25 | Shechem Mountains | |
| Samarea Mountains | PIL-26 | Kedumim | |
| Carmel Mountain | PIL-30 | Kerem Maharal South | |
| Judean Hills | PIL-33 | Sarisa | |
| Lower Galilee | PIL-34 | Ahuzat Barak | |
| Lower Galilee | PIL-36 | Nau’ra | |
| Lupinus palaestinus | Sharon—Coastal Plain | PA-2 | Pardes Hana |
| Sharon—Coastal Plain | PA-3 | Ilanot Forest West | |
| Coastal Plain (East) | PA-4 | Yashresh | |
| Sharon—Coastal Plain | PA-5 | Bnei-Tzion North | |
| Sharon—Coastal Plain | PA-7 | Ilanot Forest East | |
| Sharon—Coastal Plain | PA-10 | Netanya | |
| Sharon—Coastal Plain | PA-11 | Hirbet Samara | |
| Sharon—Coastal Plain | PA-12 | Hod Ha-Sharon | |
| Sharon—Coastal Plain | PA-15 | Tel Mond—Kurkar | |
| Coastal Plain (South) | PA-16 | Ashqelon | |
| Coastal Plain (East) | PA-17 | Sitriya | |
| Lupinus albus (bitter) | Egypt, Central Nile | ALB-05 * | |
| Egypt, Central Nile | ALB-07 * | ||
| Lupinus albus (sweet) | Commercial Product 1 | ALB-12 | |
| Commercial Product 2 | ALB-sh | ||
| Lupinus angustifolius | Sharon—Coastal Plain | ANG-sh * | Haogen |
| Lupinus luteus | LUT-03 | ||
| Lupinus mutabilis | The Netherlands | MUT-01 ** | Lupinus mutabilis, var. cruickshankii. Vreeken’s Zaden Seed Company, Dordrecht, The Netherlands, www.vreeken.nl (accessed on 12 March 2024) |
| Glycine max (soybean) | MAX-01 | Organic soybeans certified by Pro-Cert 2020 Crop year, Thompson Limited Canada, Net—25 kg, HOSSQ21-24, Packed 03/2021, Sell by 03/2022 | |
| Cicer arietinum (chickpea) | ARI-01 | Organic Chickpeas by Tvuot, www.tvuot.co.il (accessed on 12 March 2024), 500 g package Best before 01/06/2022 |
| No. | Compound | Precursor Ion (m/z) | Product Ion 1 (m/z) | Col. Energy 1 (eV) | Product Ion 2 (m/z) | Col. Energy 2 (eV) | Product Ion 3 (m/z) | Col. Energy 3 (eV) | Indicative RT (min) | MRM | Compound Quantified |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Standard QAs | |||||||||||
| 1 | Gramine | 130.0 | 77.0 | 20 | 103.0 | 25 | 7.35 | 1 | 1 | ||
| 2 | Epilupinine | 170.2 | 96.0 | 30 | 152.0 | 20 | 124.0 | 25 | 5.50 | 1 | 2 |
| 3 | Lupinine | 170.2 | 124.0 | 25 | 152.0 | 20 | 96.0 | 30 | 6.00 | 1 | 3 |
| 4 | Cytisine * | 191.2 | 133.0 | 30 | 148.0 | 20 | 5.70 | 1 | 4 | ||
| 5 | Methylcytisine * | 205.2 | 58.2 | 20 | 108.0 | 20 | 7.20 | 1 | 5 | ||
| 6 | Albine | 233.2 | 112.0 | 20 | 138.0 | 20 | 150.0 | 30 | 7.15 | 2 | 6 |
| 7 | Angustifoline | 235.2 | 112.0 | 20 | 193.0 | 30 | 114.0 | 30 | 8.15 | 2 | 7 |
| 8 | Sparteine | 235.2 | 98.0 | 30 | 233.0 | 30 | 70.0 | 30 | 7.65 | 2 | 8 |
| 9 | Anagyrine * | 245.2 | 70.0 | 45 | 98.0 | 35 | 9.40 | 2 | 9 | ||
| 10 | Thermopsine * | 245.2 | 70.0 | 45 | 98.0 | 35 | 9.50 | 2 | 10 | ||
| 11 | Multiflorine | 247.2 | 70.0 | 40 | 112.0 | 25 | 134.0 | 20 | 7.65 | 2 | 11 |
| 12 | Lupanine | 249.2 | 114.0 | 30 | 136.0 | 30 | 98.0 | 30 | 8.10 | 3 | 12 |
| 13 | Isolupanine | 249.2 | 84.0 | 30 | 98.0 | 30 | 136.0 | 30 | 10.50 | 3 | 13 |
| 14 | 13α-Hydroxylupanine | 265.2 | 114.0 | 30 | 152.0 | 30 | 84.0 | 40 | 6.95 | 3 | 14 |
| 15 | 13α-trans-Cinnamoyloxylupanine * | 395.2 | 112.0 | 30 | 247.0 | 30 | 98.0 | 40 | 11.55 | 1 | 15 |
| Non-standard QAs | |||||||||||
| 16 | Isoangustifoline | 235.2 | 112.0 | 20 | 193.0 | 30 | 114.0 | 30 | 8.00 | 2 | 7 |
| 17 | 5,6-Didehydromultiflorine | 245.2 | 70.0 | 45 | 98.0 | 35 | 7.60 | 2 | 9 | ||
| 18 | 11,12-seco-12,13-Didehydromultiflorine | 247.2 | 98.0 | 30 | 112.0 | 25 | 70 | 40 | 9.95 | 2 | 11 |
| 19 | Tetrahydrorhombifoline | 249.2 | 114.0 | 30 | 136.0 | 30 | 166.0 | 30 | 11.40 | 3 | 13 |
| 20 | 13α-Hydroxymultiflorine | 263.2 | 112.0 | 30 | 245.0 | 20 | 70.0 | 40 | 5.65 | 3 | 14 |
| 21 | 3β-Hydroxylupanine | 265.2 | 114.0 | 30 | 152.0 | 30 | 112.0 | 35 | 6.80 | 3 | 14 |
| 22 | 3β,13α-Dihydroxylupanine | 281.2 | 130.0 | 25 | 152.0 | 25 | 101.0 | 35 | 5.95 | 3 | 14 |
| 23 | 13α-Tigloyloxymultiflorine | 345.2 | 112.0 | 30 | 245.0 | 20 | 70.0 | 40 | 10.10 | 1 | 15 |
| 24 | 13α-Angeloyloxymultiflorine | 345.2 | 112.0 | 30 | 245.0 | 20 | 70.0 | 40 | 10.20 | 1 | 15 |
| 25 | 13α-Tigloyloxylupanine | 347.2 | 112.0 | 30 | 247.0 | 30 | 98.0 | 40 | 10.70 | 1 | 15 |
| 26 | 13α-Angeloyloxylupanine | 347.2 | 112.0 | 30 | 247.0 | 30 | 98.0 | 40 | 10.85 | 1 | 15 |
| 27 | 13α-cis-Cinnamoyloxylupanine | 395.2 | 112.0 | 30 | 247.0 | 30 | 98.0 | 40 | 11.15 | 1 | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Namdar, D.; Mulder, P.P.J.; Ben-Simchon, E.; Hacham, Y.; Basheer, L.; Cohen, O.; Sternberg, M.; Shelef, O. New Analytical Approach to Quinolizidine Alkaloids and Their Assumed Biosynthesis Pathways in Lupin Seeds. Toxins 2024, 16, 163. https://doi.org/10.3390/toxins16030163
Namdar D, Mulder PPJ, Ben-Simchon E, Hacham Y, Basheer L, Cohen O, Sternberg M, Shelef O. New Analytical Approach to Quinolizidine Alkaloids and Their Assumed Biosynthesis Pathways in Lupin Seeds. Toxins. 2024; 16(3):163. https://doi.org/10.3390/toxins16030163
Chicago/Turabian StyleNamdar, Dvory, Patrick P. J. Mulder, Eyal Ben-Simchon, Yael Hacham, Loai Basheer, Ofer Cohen, Marcelo Sternberg, and Oren Shelef. 2024. "New Analytical Approach to Quinolizidine Alkaloids and Their Assumed Biosynthesis Pathways in Lupin Seeds" Toxins 16, no. 3: 163. https://doi.org/10.3390/toxins16030163