Development of a Rapid and Sensitive CANARY Biosensor Assay for the Detection of Shiga Toxin 2 from Escherichia coli


Abstract
:1. Introduction
2. Results
2.1. Development and Characterization of a CANARY® B-Cell Based Assay to Detect Shiga Toxin 2
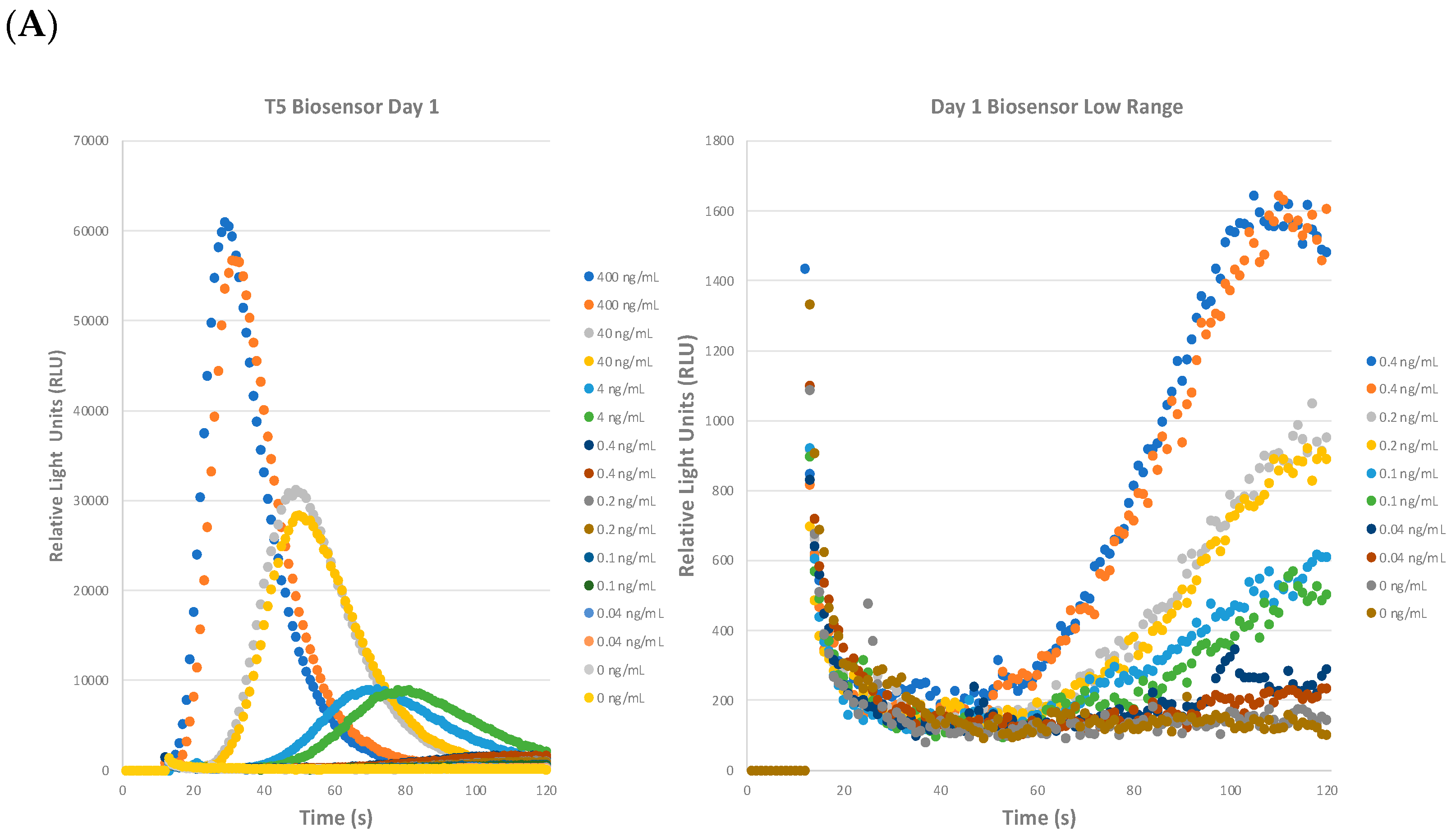
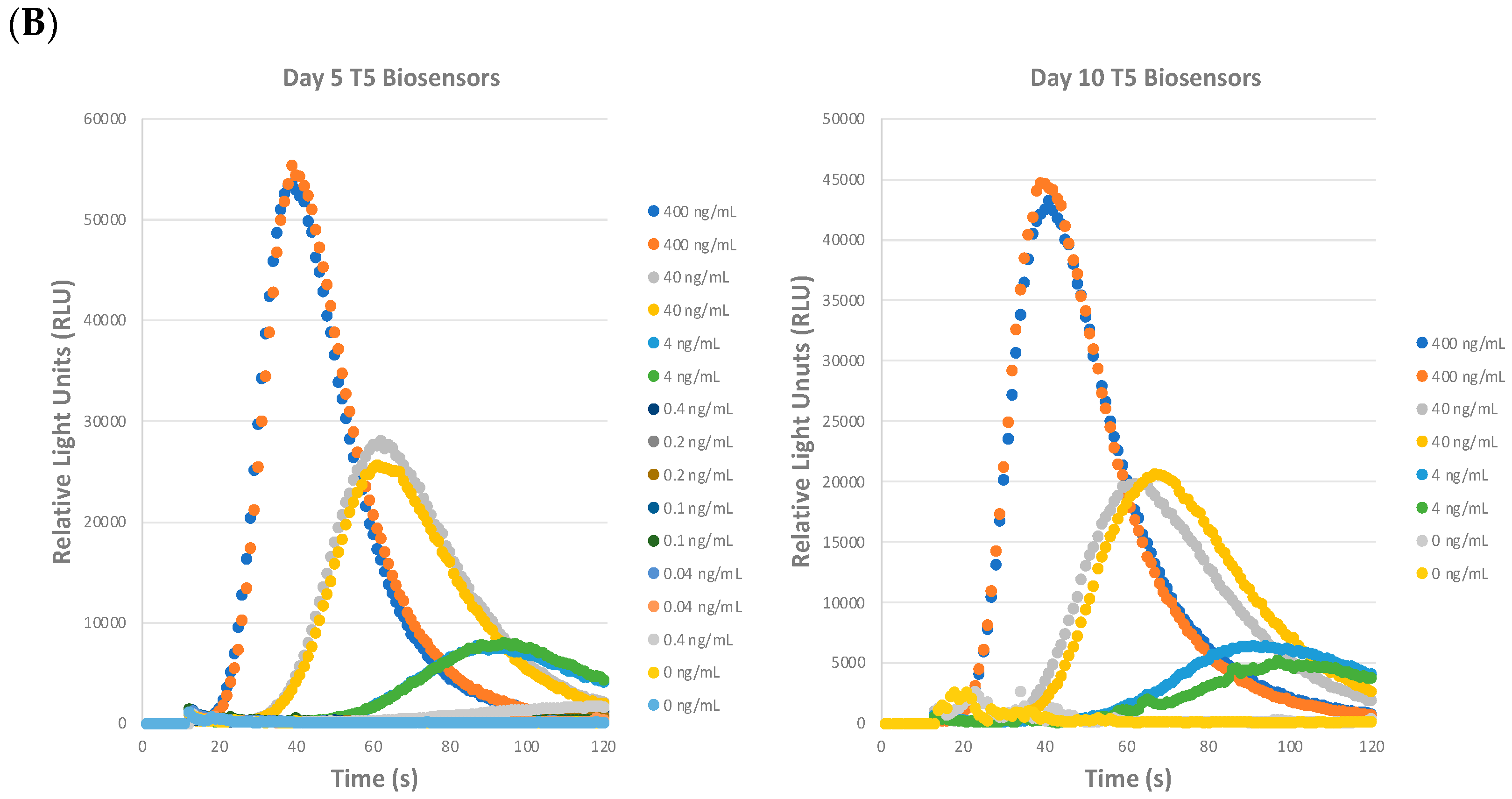
2.2. Precision of the STEC Biosensor Assay: Repeatability, Reproducibility, and Shelf-Life
2.3. STEC Biosensor Specifically Recognizes Shiga Toxins
2.4. Detection of Native Stx2 Produced by STEC in Media
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents
5.2. Biosensor Engineering
5.3. Biosensor Reconstitution
5.4. STEC Biosensor Assay for Stx2a Toxoid Detection
5.5. Shelf-Life of STEC Biosensors
5.6. STEC Biosensor Specificity Assay
5.7. Initial E. coli 0121:H19 BPW + Novobiocin +/− Mitomycin C Study
5.8. Detection of Stx from STEC Supernatants with T5 Biosensor
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Centers for Disease Control and Prevention, National Outbreak Reporting System Dashboard. Available online: https://wwwn.cdc.gov/norsdashboard/ (accessed on 13 September 2023).
- Siegler, R.L. Postdiarrheal Shiga toxin-mediated hemolytic uremic syndrome. JAMA 2003, 290, 1379–1381. [Google Scholar] [CrossRef]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef]
- Russmann, H.; Schmidt, H.; Heesemann, J.; Caprioli, A.; Karch, H. Variants of Shiga-like toxin II constitute a major toxin component in Escherichia coli O157 strains from patients with haemolytic uraemic syndrome. J. Med. Microbiol. 1994, 40, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Head, S.C.; Karmali, M.A.; Lingwood, C.A. Preparation of VT1 and VT2 hybrid toxins from their purified dissociated subunits. Evidence for B subunit modulation of a subunit function. J. Biol. Chem. 1991, 266, 3617–3621. [Google Scholar] [CrossRef] [PubMed]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef]
- Lacher, D.W.; Gangiredla, J.; Patel, I.; Elkins, C.A.; Feng, P.C. Use of the Escherichia coli Identification Microarray for Characterizing the Health Risks of Shiga Toxin-Producing Escherichia coli Isolated from Foods. J. Food Prot. 2016, 79, 1656–1662. [Google Scholar] [CrossRef]
- Di, R.; Kyu, E.; Shete, V.; Saidasan, H.; Kahn, P.C.; Tumer, N.E. Identification of amino acids critical for the cytotoxicity of Shiga toxin 1 and 2 in Saccharomyces cerevisiae. Toxicon 2011, 57, 525–539. [Google Scholar] [CrossRef]
- Endo, Y.; Tsurugi, K.; Yutsudo, T.; Takeda, Y.; Ogasawara, T.; Igarashi, K. Site of action of a Vero toxin (VT2) from Escherichia coli O157:H7 and of Shiga toxin on eukaryotic ribosomes. RNA N-glycosidase activity of the toxins. Eur. J. Biochem. 1988, 171, 45–50. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Marques, L.R.; Newland, J.W.; Smith, H.W.; Holmes, R.K.; O’Brien, A.D. Two toxin-converting phages from Escherichia coli O157:H7 strain 933 encode antigenically distinct toxins with similar biologic activities. Infect. Immun. 1986, 53, 135–140. [Google Scholar] [CrossRef]
- Luna-Gierke, R.E.; Griffin, P.M.; Gould, L.H.; Herman, K.; Bopp, C.A.; Strockbine, N.; Mody, R.K. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol. Infect. 2014, 142, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Krause, G.; Zimmermann, S.; Kaulfuss, S.; Gleier, K. Characterization of Shiga toxin-producing Escherichia coli strains isolated from human patients in Germany over a 3-year period. J. Clin. Microbiol. 2004, 42, 1099–1108. [Google Scholar] [CrossRef]
- Fasel, D.; Mellmann, A.; Cernela, N.; Hachler, H.; Fruth, A.; Khanna, N.; Egli, A.; Beckmann, C.; Hirsch, H.H.; Goldenberger, D.; et al. Hemolytic uremic syndrome in a 65-Year-old male linked to a very unusual type of stx2e- and eae-harboring O51:H49 shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2014, 52, 1301–1303. [Google Scholar] [CrossRef]
- Friesema, I.; van der Zwaluw, K.; Schuurman, T.; Kooistra-Smid, M.; Franz, E.; van Duynhoven, Y.; van Pelt, W. Emergence of Escherichia coli encoding Shiga toxin 2f in human Shiga toxin-producing E. coli (STEC) infections in the Netherlands, January 2008 to December 2011. Euro Surveill 2014, 19, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, J.A.; Clark, M.; Cariou, A.; Leveau, A.; Pierre, S.; Fratamico, P.; Strobaugh, T.P., Jr.; Armstrong, C.M. Detection of Shiga toxin-producing Escherichia coli (STEC) in beef products using droplet digital PCR. Int. J. Food Microbiol. 2020, 319, 108499. [Google Scholar] [CrossRef] [PubMed]
- Tesh, V.L.; Burris, J.A.; Owens, J.W.; Gordon, V.M.; Wadolkowski, E.A.; O’Brien, A.D.; Samuel, J.E. Comparison of the relative toxicities of Shiga-like toxins type I and type II for mice. Infect. Immun. 1993, 61, 3392–3402. [Google Scholar] [CrossRef] [PubMed]
- Mulchandani, R.; Brehmer, C.; Butt, S.; Vishram, B.; Harrison, M.; Marchant, E.; Ferris, S.; Jorgensen, F.; Smith, R.; Godbole, G.; et al. Outbreak of Shiga toxin-producing Escherichia coli O157 linked with consumption of a fast-food product containing imported cucumbers, United Kingdom, August 2020. Int. J. Infect. Dis. 2021, 110 (Suppl. S1), S62–S68. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.E.; Mbandi, E.; Buchanan, S.; Abdelmajid, N.; Gonzalez-Rivera, C.; Hale, K.R.; Jacobsen, L.; Webb, J.; Green, J.; Dolan, P. Salmonella and Shiga Toxin-Producing Escherichia coli in Products Sampled in the Food Safety and Inspection Service Raw Pork Baseline Study. J. Food Prot. 2020, 83, 552–559. [Google Scholar] [CrossRef]
- Garofalo, M.; Payros, D.; Taieb, F.; Oswald, E.; Nougayrede, J.P.; Oswald, I.P. From ribosome to ribotoxins: Understanding the toxicity of deoxynivalenol and Shiga toxin, two food borne toxins. Crit. Rev. Food Sci. Nutr. 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Franz, E.; Delaquis, P.; Morabito, S.; Beutin, L.; Gobius, K.; Rasko, D.A.; Bono, J.; French, N.; Osek, J.; Lindstedt, B.A.; et al. Exploiting the explosion of information associated with whole genome sequencing to tackle Shiga toxin-producing Escherichia coli (STEC) in global food production systems. Int. J. Food Microbiol. 2014, 187, 57–72. [Google Scholar] [CrossRef]
- Egervarn, M.; Flink, C. Shiga toxin-producing Escherichia coli (STEC) in meat and leafy greens available in the Swedish retail market—Occurrence and diversity of stx subtypes and serotypes. Int. J. Food Microbiol. 2024, 408, 110446. [Google Scholar] [CrossRef]
- He, X.; Kong, Q.; Patfield, S.; Skinner, C.; Rasooly, R. A New Immunoassay for Detecting All Subtypes of Shiga Toxins Produced by Shiga Toxin-Producing E. coli in Ground Beef. PLoS ONE 2016, 11, e0148092. [Google Scholar] [CrossRef]
- Hughes, A.C.; Patfield, S.; Rasooly, R.; He, X. Validation of a Cell-Based Assay for Detection of Active Shiga Toxins Produced by Escherichia coli in Water. Int. J. Environ. Res. Public Health 2020, 17, 7901. [Google Scholar] [CrossRef]
- Russo, L.M.; Melton-Celsa, A.R.; Smith, M.A.; Smith, M.J.; O’Brien, A.D. Oral intoxication of mice with Shiga toxin type 2a (Stx2a) and protection by anti-Stx2a monoclonal antibody 11E10. Infect. Immun. 2014, 82, 1213–1221. [Google Scholar] [CrossRef]
- Armstrong, C.M.; Ruth, L.E.; Capobianco, J.A.; Strobaugh, T.P., Jr.; Rubio, F.M.; Gehring, A.G. Detection of Shiga Toxin 2 Produced by Escherichia coli in Foods Using a Novel AlphaLISA. Toxins 2018, 10, 422. [Google Scholar] [CrossRef]
- Rocchetti, L.; Munari, B.; Varrone, E.; Porcellini, E.; Orth-Holler, D.; Wurzner, R.; Carnicelli, D.; Brigotti, M. Method for the Detection of the Cleaved Form of Shiga Toxin 2a Added to Normal Human Serum. Toxins 2021, 13, 94. [Google Scholar] [CrossRef] [PubMed]
- Manage, D.P.; Lauzon, J.; Jones, C.M.; Ward, P.J.; Pilarski, L.M.; Pilarski, P.M.; McMullen, L.M. Detection of pathogenic Escherichia coli on potentially contaminated beef carcasses using cassette PCR and conventional PCR. BMC Microbiol. 2019, 19, 175. [Google Scholar] [CrossRef]
- Li, B.; Liu, H.; Wang, W. Multiplex real-time PCR assay for detection of Escherichia coli O157:H7 and screening for non-O157 Shiga toxin-producing E. coli. BMC Microbiol. 2017, 17, 215. [Google Scholar] [CrossRef] [PubMed]
- Aminabad, E.D.; Hasanzadeh, M.; Ahmadalipour, A.; Mahmoudi, T.; Feizi, M.A.H.; Safaralizadeh, R.; Mobed, A. Sensitive electrochemical recognition of alpha-synuclein protein in human plasma samples using bioconjugated gold nanoparticles: An innovative immuno-platform to assist in the early stage identification of Parkinson’s disease by biosensor technology. J. Mol. Recognit. 2023, 36, e2952. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Bai, L.; Zhao, L.; Wu, L.; Lv, H.; Li, Q.; Li, X.; Xie, Q.; Wang, L.; Liu, C.; et al. Standardized Shiga-Toxin Encoding Genes Real-Time PCR Screening Methods Comparison and Development of an Internally Controlled Assay for Pan-stx2 Detection. J. AOAC Int. 2021, 104, 1065–1071. [Google Scholar] [CrossRef]
- Fratamico, P.M.; Wasilenko, J.L.; Garman, B.; Demarco, D.R.; Varkey, S.; Jensen, M.; Rhoden, K.; Tice, G. Evaluation of a multiplex real-time PCR method for detecting shiga toxin-producing Escherichia coli in beef and comparison to the U.S. Department of Agriculture Food Safety and Inspection Service Microbiology laboratory guidebook method. J. Food Prot. 2014, 77, 180–188. [Google Scholar] [CrossRef]
- Baker, C.A.; Rubinelli, P.M.; Park, S.H.; Ricke, S.C. Immuno-based detection of Shiga toxin-producing pathogenic Escherichia coli in food—A review on current approaches and potential strategies for optimization. Crit. Rev. Microbiol. 2016, 42, 656–675. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.A.; Bauerfeind, R.; Berens, C.; Menge, C. Shiga Toxin-Producing E. coli in Animals: Detection, Characterization, and Virulence Assessment. Methods Mol. Biol. 2021, 2291, 19–86. [Google Scholar] [CrossRef] [PubMed]
- Costa-Ribeiro, A.; Azinheiro, S.; Roumani, F.; Prado, M.; Lamas, A.; Garrido-Maestu, A. Multiplex Real-Time PCR for the Detection of Shiga Toxin-Producing Escherichia coli in Foods. Methods Mol. Biol. 2023, 2967, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Medina, M.; Greaves, I.; Hunter, P.R.; Minnigh, H.; Ramirez-Toro, G. Detection of Shiga toxin-encoding genes in small community water supplies. J. Water Health 2020, 18, 937–945. [Google Scholar] [CrossRef]
- Ren, K.; Duan, M.; Su, T.; Ying, D.; Wu, S.; Wang, Z.; Duan, N. A colorimetric and SERS dual-mode aptasensor for the detection of Shiga toxin type II based on Mn/Fe-MIL(53)@AuNSs. Talanta 2024, 270, 125636. [Google Scholar] [CrossRef]
- Cao, Y.; Fang, T.; Shen, J.; Zhang, G.; Guo, D.; Zhao, L.; Jiang, Y.; Zhi, S.; Zheng, L.; Lv, X.; et al. Development of Recombinase Aided Amplification (RAA)-Exo-Probe Assay for the Rapid Detection of Shiga Toxin-Producing Escherichia coli. J. AOAC Int. 2023, 106, 1246–1253. [Google Scholar] [CrossRef]
- Lee, J.E.; Toushik, S.H.; Park, H.J.; Kim, S.A.; Shim, W.B. Rapid detection of Shiga-toxin-producing Escherichia coli O157:H7 based on a colorimetric loop-mediated isothermal amplification (cLAMP) assay using a molecular beacon paired with HRPzyme. Anal. Bioanal. Chem. 2023, 415, 4973–4984. [Google Scholar] [CrossRef] [PubMed]
- Wasiewska, L.A.; Juska, V.B.; Seymour, I.; Burgess, C.M.; Duffy, G.; O’Riordan, A. Electrochemical nucleic acid-based sensors for detection of Escherichia coli and Shiga toxin-producing E. coli-Review of the recent developments. Compr. Rev. Food Sci. Food Saf. 2023, 22, 1839–1863. [Google Scholar] [CrossRef]
- Manfredi, E.; Rocca, M.F.; Zintgraff, J.; Irazu, L.; Miliwebsky, E.; Carbonari, C.; Deza, N.; Prieto, M.; Chinen, I. Rapid and accurate detection of Shiga toxin-producing Escherichia coli (STEC) serotype O157:H7 by mass spectrometry directly from the isolate, using 10 potential biomarker peaks and machine learning predictive models. J. Med. Microbiol. 2023, 72, 001675. [Google Scholar] [CrossRef]
- Cheng, L.W.; Henderson, T.D.; Patfield, S.; Stanker, L.H.; He, X. Mouse in vivo neutralization of Escherichia coli Shiga toxin 2 with monoclonal antibodies. Toxins 2013, 5, 1845–1858. [Google Scholar] [CrossRef]
- Rider, T.H.; Petrovick, M.S.; Nargi, F.E.; Harper, J.D.; Schwoebel, E.D.; Mathews, R.H.; Blanchard, D.J.; Bortolin, L.T.; Young, A.M.; Chen, J.; et al. A B cell-based sensor for rapid identification of pathogens. Science 2003, 301, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, R.A.; Ozanich, R.M.; Arce, J.S.; Engelmann, H.E.; Heredia-Langner, A.; Hofstad, B.A.; Hutchison, J.R.; Jarman, K.; Melville, A.M.; Victry, K.D.; et al. Evaluation of Immunoassays and General Biological Indicator Tests for Field Screening of Bacillus anthracis and Ricin. Health Secur. 2017, 15, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.C.; Flannery, A.R.; Cheng, L.W. A Rapid, Sensitive, and Portable Biosensor Assay for the Detection of Botulinum Neurotoxin Serotype A in Complex Food Matrices. Toxins 2018, 10, 476. [Google Scholar] [CrossRef] [PubMed]
- He, X.; McMahon, S.; Skinner, C.; Merrill, P.; Scotcher, M.C.; Stanker, L.H. Development and characterization of monoclonal antibodies against Shiga toxin 2 and their application for toxin detection in milk. J. Immunol. Methods 2013, 389, 18–28. [Google Scholar] [CrossRef]
- Skinner, C.; Patfield, S.; Stanker, L.H.; Fratamico, P.; He, X. New high-affinity monoclonal antibodies against Shiga toxin 1 facilitate the detection of hybrid Stx1/Stx2 in vivo. PLoS ONE 2014, 9, e99854. [Google Scholar] [CrossRef]
- Skinner, C.; Patfield, S.; Khalil, R.; Kong, Q.; He, X. New Monoclonal Antibodies against a Novel Subtype of Shiga Toxin 1 Produced by Enterobacter cloacae and Their Use in Analysis of Human Serum. mSphere 2016, 1, 10-1128. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Detection |
|---|---|
| Stx2a | + |
| Stx2b | + |
| Stx2c | + |
| Stx2d | + |
| Stx2e | ND |
| Stx2f | ND |
| Stx2g | + |
| Stx1a | ND |
| Abrin | ND |
| Ricin | ND |
| Strains | Type | Detection |
|---|---|---|
| RM5856 | O157:H7 | + |
| RM1913 | O157:H7 | + |
| RM10466 | O103 | + |
| RM12788 | O111 (stx1 and stx2) | + |
| RM6848 | O121 | + |
| RM9872 | O149 | + |
| ATCC25922 | O6 (stx-) | ND |
| RM7103 | O45 (stx-) | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tam, C.C.; Wang, Y.; Du, W.-X.; Flannery, A.R.; He, X. Development of a Rapid and Sensitive CANARY Biosensor Assay for the Detection of Shiga Toxin 2 from Escherichia coli. Toxins 2024, 16, 148. https://doi.org/10.3390/toxins16030148
Tam CC, Wang Y, Du W-X, Flannery AR, He X. Development of a Rapid and Sensitive CANARY Biosensor Assay for the Detection of Shiga Toxin 2 from Escherichia coli. Toxins. 2024; 16(3):148. https://doi.org/10.3390/toxins16030148
Chicago/Turabian StyleTam, Christina C., Yangyang Wang, Wen-Xian Du, Andrew R. Flannery, and Xiaohua He. 2024. "Development of a Rapid and Sensitive CANARY Biosensor Assay for the Detection of Shiga Toxin 2 from Escherichia coli" Toxins 16, no. 3: 148. https://doi.org/10.3390/toxins16030148