Toxic Effects of Bt-(Cry1Ab+Vip3Aa) Maize on Storage Pest Paralipsa gularis (Zeller)


Abstract
:1. Introduction
2. Results
2.1. Susceptibility of P. gularis to Bt Insecticidal Proteins
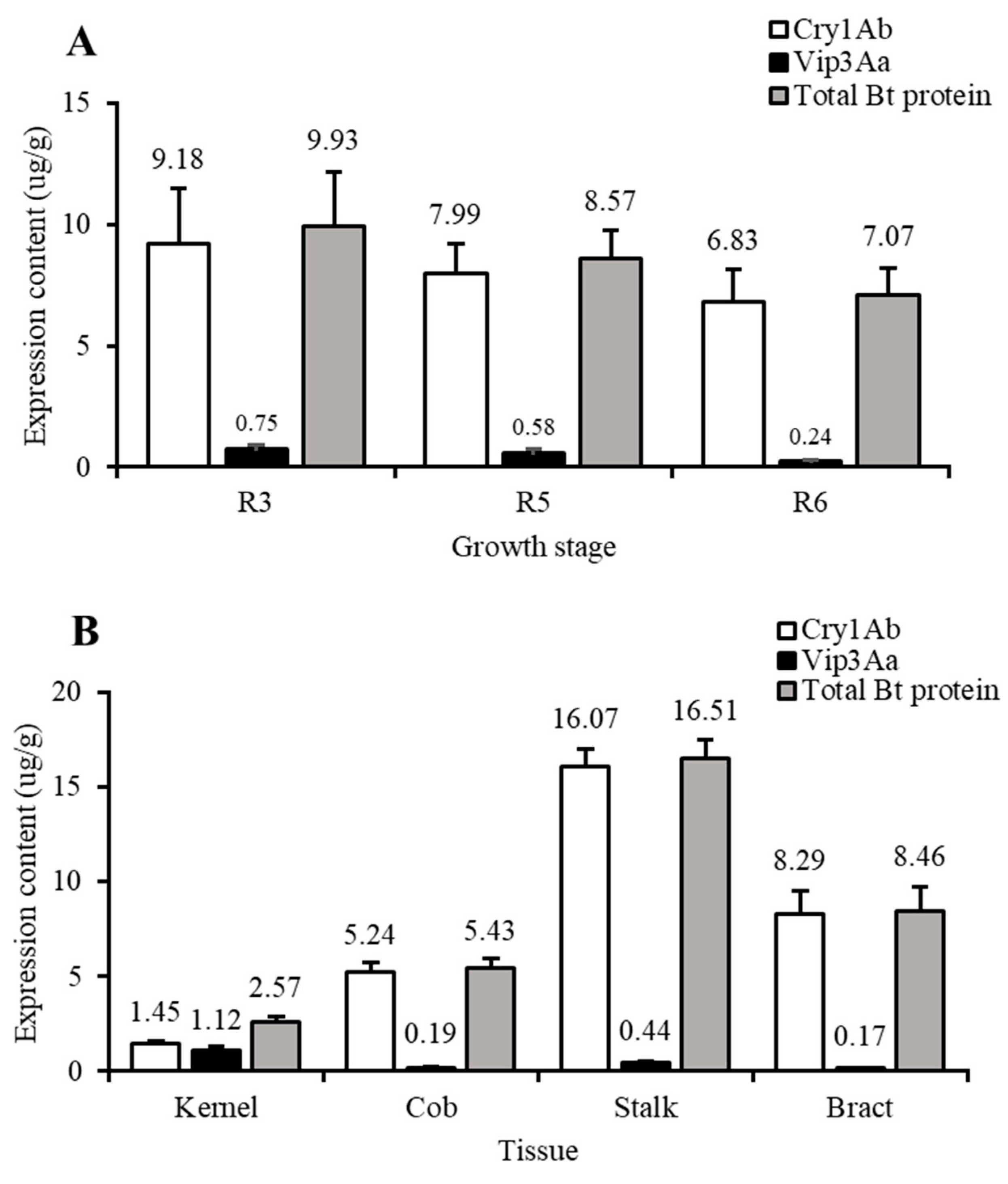
2.2. Insecticidal Protein Expression Content and Insecticidal Activity of Different Tissues in Bt-(Cry1Ab+Vip3Aa) Maize
2.3. Control Efficiency of Bt-(Cry1Ab+Vip3Aa) Maize against P. gularis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Collection and Rearing of P. gularis
5.2. Determination of the Susceptibility of P. gularis to Bt Insecticidal Proteins
5.3. Determination of Insecticidal Protein Expression Content and Insecticidal Activity in Different Tissues of Bt-(Cry1Ab+Vip3Aa) Maize
5.4. Field Surveys of the Control Efficiency of Bt-(Cry1Ab+Vip3Aa) Maize against P. gularis
5.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hong, K.J.; Lee, J.H.; Lee, G.S.; Lee, S. The status quo of invasive alien insect species and plant quarantine in Korea. J. Asia-Pac. Entomol. 2012, 15, 521–532. [Google Scholar] [CrossRef]
- Kageyama, D.; Narita, S.; Imamura, T.; Miyanoshita, A. Detection and identification of Wolbachia endosymbionts from laboratory stocks of stored-product insect pests and their parasitoids. J. Stored Prod. Res. 2010, 46, 13–19. [Google Scholar] [CrossRef]
- Wang, P.Y. Economic Insect Fauna of China; Science Press: Beijing, China, 1980; pp. 33–34. [Google Scholar]
- Trematerra, P. Distribution and activity of Paralipsa gularis (Zeller) (Lep., Galleriidae) in Italy. J. Appl. Entomol. 1987, 104, 227–233. [Google Scholar] [CrossRef]
- Tai, H.K.; Bai, S.X.; Han, Y.L.; Liu, Z.; Zhang, F.; Wang, Z.Y. Biological characteristics of the stored nut moth Paralipsa gularis (Zeller) and its damage on corn in Dehong prefecture of Yunnan province. J. Plant Prot. 2018, 45, 251–256. [Google Scholar]
- Chen, S.; Zhao, S.Y.; Liu, D.Z.; Yang, X.M.; Li, N.; Wu, K.M. The damage characteristics and spatial distribution pattern of the new maize pest Paralipsa gularis (Zeller) in southwestern China. Chin. J. Appl. Entomol. 2022, 59, 1385–1393. [Google Scholar]
- Xie, X.B.; Meng, J.Z.; Li, H.H.; Zhao, G.A.; Xu, J.B.; Shen, Y.F. Preliminary report on the damage caused by Paralipsa gularis (Zeller) in summer corn fields in Baoshan, Yunnan. China Plant Prot. 2023, 43, 38–40. [Google Scholar]
- Zhao, X.M.; Liu, Y.; Zheng, X.; Li, Q.C.; Wang, L.X.; Liu, Y.; Lan, Y.; Wang, Z.Y. Current status, strategies and suggestions for controlling the corn pests in China. Mod. Agrochem. 2022, 21, 6–14. [Google Scholar]
- Wu, K.M. Development direction of crop pest control science and technology in China. J. Agric. 2018, 8, 35–38. [Google Scholar]
- Song, Y.F.; Wu, K.M. Investigation on controlling status of fall armyworm in sweet/waxy corn fields in western Yunnan province. Plant Prot. 2020, 46, 217–222. [Google Scholar]
- Li, Q.; Men, X.Y.; Jing, C.; Yu, Y.; Zhou, X.H.; Dai, X.Y.; Lü, S.H.; Li, L.L. Research progress in emergency prevention and control of Spodoptera frugiperda in China. Plant Prot. 2021, 47, 21–27. [Google Scholar]
- Diez-rodriguez, G.I.; Omoto, C. Inheritance of lambda-cyhalothrin resistance in Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Neotrop. Entomol. 2001, 30, 311–316. [Google Scholar] [CrossRef]
- Lai, T.C.; Li, J.; Su, J.Y. Monitoring of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) resistance to chlorantraniliprole in China. Pestic. Biochem. Physiol. 2011, 101, 198–205. [Google Scholar] [CrossRef]
- You, M.S. Conservation and utilization of the insect diversity in China. Biodivers. Sci. 1997, 5, 135–141. [Google Scholar]
- Karabörklü, S.; Azizoglu, U.; Azizoglu, Z.B. Recombinant entomopathogenic agents: A review of biotechnological approaches to pest insect control. World J. Microbiol. Biotechnol. 2018, 34, 14. [Google Scholar] [CrossRef] [PubMed]
- Afidchao, M.M.; Musters, C.J.M.; De Snoo, G.R. Asian corn borer (ACB) and non-ACB pests in GM corn (Zea mays L.) in the Philippines. Pest Manag. Sci. 2013, 69, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Botha, A.S.; Erasmus, A.; Du Plessis, H.; Van den Berg, J. Efficacy of Bt maize for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in South Africa. J. Econ. Entomol. 2019, 112, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Li, G.P.; Wu, K.M. Commercial strategy of transgenic insect-resistant maize in China. J. Plant Prot. 2022, 49, 17–32. [Google Scholar]
- International Service for the Acquisition of Agri-biotech Applications (ISAAA). In Proceedings of the Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; ISAAA Brief 2019, No. 55; ISAAA: Ithaca, NY, USA, 2019. [Google Scholar]
- Li, Y.; Wang, T.Y. Germplasm enhancement in maize: Advances and prospects. J. Maize Sci. 2018, 26, 1–15. [Google Scholar]
- Wu, J.Z.; Zhang, J.; Ge, Z.M.; Xing, L.W.; Han, S.Q.; Shen, C.; Kong, F.T. Impact of climate change on maize yield in China from 1979 to 2016. J. Integr. Agric. 2021, 20, 289–299. [Google Scholar] [CrossRef]
- Wu, L.H.; Zheng, L.X. Research progress in maize pests. Guizhou Agric. Sci. 2018, 46, 53–58. [Google Scholar]
- Zhao, S.Y.; Yang, X.M.; Liu, D.Z.; Sun, X.X.; Li, G.P.; Wu, K.M. Performance of the domestic Bt corn event expressing pyramided Cry1Ab and Vip3Aa19 against the invasive Spodoptera frugiperda (J. E. Smith) in China. Pest Manag. Sci. 2023, 79, 1018–1029. [Google Scholar] [CrossRef]
- Sun, D.D.; Quan, Y.D.; Wang, Y.Q.; Wang, Z.Y.; He, K.L. Resistance of transgenic Bt maize (Ruifeng125, DBN9936 & DBN9978) to Asian corn borer. Plant Prot. 2021, 47, 206–211. [Google Scholar]
- Sun, H.W.; Li, F.; Gao, R.; Xu, X.H.; Ke, Y.S.; Bo, L.X. Bt protein spatial-temporal expression and evaluation for resistance of transgenic Cry1Ab/Cry2Aj and G10evo-epsps maize. J. Biosaf. 2018, 27, 63–68. [Google Scholar]
- Le, D.K.; Le, Q.K.; Tran, T.T.H.; Nguyen, D.V.; Dao, T.H.; Nguyen, T.T.; Truong, X.L.; Nguyen, Q.C.; Pham, H.P.; Phan, T.T.T.; et al. Baseline susceptibility of Asian corn borer (Ostrinia furnacalis (Guenée)) populations in Vietnam to Cry1Ab insecticidal protein. J. Asia-Pac. Entomol. 2019, 22, 493–498. [Google Scholar] [CrossRef]
- Hu, X.H.; Wang, Z.Y.; He, K.L.; Xie, H.C.; Wang, Y.Q. Insecticidal activity and synergism of combinations of Bacillus thuringiensis Cry and Vip3Aa against Ostrinia furnacalis. Plant Prot. 2023, 49, 310–315. [Google Scholar]
- Tabashnik, B.E. Evaluation of synergism among Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 1992, 58, 3343–3346. [Google Scholar] [CrossRef] [PubMed]
- Hatzios, K.K. Interactions of herbicides with other agrochemicals in higher plants. Rev. Weed Sci. 1985, 1, 1–63. [Google Scholar]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing pyramided transgenic Bt crops for sustainable pest management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Bergamasco, V.B.; Mendes, D.R.P.; Fernandes, O.A.; Desidério, J.A.; Lemos, M.V.F. Bacillus thuringiensis Cry1Ia10 and Vip3Aa protein interactions and their toxicity in Spodoptera spp. (Lepidoptera). J. Invertebr. Pathol. 2013, 112, 152–158. [Google Scholar] [CrossRef]
- Soares Figueiredo, C.; Nunes Lemes, A.R.; Sebastião, I.; Desidério, J.A. Synergism of the Bacillus thuringiensis Cry1, Cry2, and Vip3 proteins in Spodoptera frugiperda control. Appl. Biochem. Biotechnol. 2019, 188, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Zhang, D.D.; Zhao, S.Y.; Wu, K.M. Susceptibilities of the invasive fall armyworm (Spodoptera frugiperda) to the insecticidal proteins of Bt maize in China. Toxins 2022, 14, 507. [Google Scholar] [CrossRef]
- Liu, X.B.; Liu, S.; Long, Y.; Wang, Y.Q.; Zhao, W.L.; Shwe, S.M.; Wang, Z.Y.; He, K.L.; Bai, S.X. Baseline susceptibility and resistance allele frequency in Ostrinia furnacalis in relation to Cry1Ab toxins in China. Toxins 2022, 14, 255. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.G.; Zhang, D.D.; Li, D.Y.; Zhao, S.Y.; Wang, C.Y.; Xiao, Y.T.; Xu, D.; Yang, Y.Z.; Li, G.P.; Wang, L.L.; et al. Expression profiles of Cry1Ab protein and its insecticidal efficacy against the invasive fall armyworm for Chinese domestic GM maize DBN9936. J. Integr. Agric. 2021, 20, 792–803. [Google Scholar] [CrossRef]
- Wang, D.Y.; Wang, Z.Y.; He, K.L.; Cong, B.; Bai, S.X.; Wen, L.P. Temporal and spatial expression of Cry1Ab toxin in transgenic Bt corn and its effects on Asian corn borer, Ostrinia furnacalis (Guenée). Sci. Agric. Sin. 2004, 37, 1155–1159. [Google Scholar]
- Storer, N.P.; Kubiszak, M.E.; King, J.E.; Thompson, G.D.; Santos, A.C. Status of resistance to Bt maize in Spodoptera frugiperda: Lessons from Puerto Rico. J. Invertebr. Pathol. 2012, 110, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J. First report of field resistance by the stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. S. Afr. J. Plant Soil 2007, 24, 147–151. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Finkenbinder, C. Field-evolved resistance in corn earworm to Cry proteins expressed by transgenic sweet corn. PLoS ONE 2016, 11, e0169115. [Google Scholar] [CrossRef]
- Kaur, G.; Guo, J.G.; Brown, S.; Head, G.P.; Price, P.A.; Paula-Moraes, S.; Ni, X.Z.; Dimase, M.; Huang, F.N. Field-evolved resistance of Helicoverpa zea (Boddie) to transgenic maize expressing pyramided Cry1A.105/Cry2Ab2 proteins in northeast Louisiana, the United States. J. Invertebr. Pathol. 2019, 163, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Farhan, Y.; Schaafsma, A.W. Practical resistance of Ostrinia nubilalis (Lepidoptera: Crambidae) to Cry1F Bacillus thuringiensis maize discovered in Nova Scotia, Canada. Sci. Rep. 2019, 9, 18247. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, J.R. Discussion about cetrtain technological links in creating new transgenic maize germplasm. J. Agric. Sci. Technol. 2011, 13, 46–52. [Google Scholar]
- Liang, G.M.; Tan, W.J.; Guo, Y.Y. Improvement of artificial feeding technology of cotton bollworm. Plant Prot. 1999, 25, 16–18. [Google Scholar]


{kind=link}
| Bt Event | Protein | N | LC50 (95%FL) μg/g | LC95 (95%FL) μg/g | Slope ± SE | χ2 | df |
|---|---|---|---|---|---|---|---|
| DBN9936 | DBNCry1Ab | 432 | 0.038 (0.013–0.062) b | 0.276 (0.207–0.455) c | 1.916 ± 0.406 | 13.043 | 13 |
| DBN9501 | DBNVip3Aa | 432 | 0.114 (0.088–0.150) a | 2.513 (1.225–8.564) a | 1.224 ± 0.169 | 18.141 | 13 |
| DBN3601T | DBN Cry1Ab+Vip3Aa | 432 | 0.110 (0.081–0.136) a | 0.606 (0.463–0.917) b | 2.217 ± 0.288 | 16.306 | 13 |
| Bt11×MIR162 | Syngenta Cry1Ab+Vip3Aa | 432 | 0.147 (0.107–0.184) a | 0.866 (0.657–1.321) ab | 2.138 ± 0.277 | 5.096 | 13 |
| Bt Event | Protein | N | GIC50 (95%FL) μg/g | GIC95 (95%FL) μg/g | Slope ± SE | χ2 | df |
|---|---|---|---|---|---|---|---|
| DBN9936 | DBNCry1Ab | 432 | 0.014 (0.000–0.035) b | 0.072 (0.020–0.096) c | 2.343 ± 0.861 | 4.965 | 13 |
| DBN9501 | DBNVip3Aa | 432 | 0.073 (0.061–0.085) a | 0.249 (0.199–0.349) a | 3.088 ± 0.176 | 34.555 | 10 |
| DBN3601T | DBN Cry1Ab+Vip3Aa | 432 | 0.027 (0.010–0.043) b | 0.129 (0.108–0.154) b | 2.442 ± 0.482 | 3.511 | 13 |
| Bt11×MIR162 | Syngenta Cry1Ab+Vip3Aa | 432 | 0.026 (0.006–0.048) b | 0.160 (0.122–0.196) b | 2.068 ± 0.450 | 1.501 | 13 |
| Growth Stage | Tissue | Cry1Ab (μg/g) | Vip3Aa (μg/g) | Total Bt Protein (μg/g) | |||
|---|---|---|---|---|---|---|---|
| Baozang Town | Longtan Town | Baozang Town | Longtan Town | Baozang Town | Longtan Town | ||
| R3 | Kernel | - | 1.02 ± 0.04 | - | 1.53 ± 0.04 | - | 2.55 ± 0.01 |
| Cob | - | 6.48 ± 1.20 | - | 0.56 ± 0.05 | - | 7.04 ± 1.25 | |
| Stalk | - | 21.60 ± 0.55 | - | 0.64 ± 0.02 | - | 22.24 ± 0.56 | |
| Bract | - | 7.62 ± 0.37 | - | 0.27 ± 0.03 | - | 7.89 ± 0.35 | |
| R5 | Kernel | 1.66 ± 0.30 | 2.42 ± 0.10 | 0.80 ± 0.04 | 2.19 ± 0.05 | 2.46 ± 0.29 | 4.61 ± 0.15 |
| Cob | 7.40 ± 0.68 | 4.89 ± 0.40 | 0.18 ± 0.03 | 0.06 ± 0.02 | 7.58 ± 0.68 | 4.95 ± 0.38 | |
| Stalk | 14.19 ± 2.96 | 15.46 ± 0.92 | 0.42 ± 0.03 | 0.74 ± 0.02 | 14.60 ± 2.97 | 16.20 ± 0.91 | |
| Bract | 15.24 ± 0.31 | 2.65 ± 0.18 | 0.20 ± 0.02 | 0.07 ± 0.00 | 15.44 ± 0.33 | 2.72 ± 0.18 | |
| R6 | Kernel | 1.12 ± 0.08 | 1.04 ± 0.02 | 0.20 ± 0.02 | 0.80 ± 0.03 | 1.40 ± 0.07 | 1.84 ± 0.02 |
| Cob | 3.63 ± 0.32 | 3.79 ± 0.11 | 0.01 ± 0.00 | 0.15 ± 0.02 | 3.64 ± 0.32 | 3.94 ± 0.08 | |
| Stalk | 14.09 ± 1.22 | 14.99 ± 0.49 | 0.12 ± 0.01 | 0.29 ± 0.02 | 14.22 ± 1.23 | 15.29 ± 0.48 | |
| Bract | 7.62 ± 3.50 | 8.32 ± 0.87 | 0.02 ± 0.01 | 0.28 ± 0.01 | 7.64 ± 3.50 | 8.59 ± 0.87 | |
| Tissue | Maize Variety | R3 | R5 | R6 | |||
|---|---|---|---|---|---|---|---|
| Baozang Town | Longtan Town | Baozang Town | Longtan Town | Baozang Town | Longtan Town | ||
| Kernel | Bt-(Cry1Ab+Vip3Aa) maize | 100.00 ± 0.00 * | 99.00 ± 1.00 * | 99.50 ± 0.50 * | 99.00 ± 0.58 * | 100.00 ± 0.00 * | 99.00 ± 0.58 * |
| Conventional maize | 17.50 ± 1.26 | 20.50 ± 1.71 | 13.00 ± 1.29 | 18.50 ± 2.36 | 55.00 ± 3.00 | 27.50 ± 1.71 | |
| Cob | Bt-(Cry1Ab+Vip3Aa) maize | 98.50 ± 0.96 * | 98.00 ± 1.15 * | 100.00 ± 0.00 * | 100.00 ± 0.00 * | 99.00 ± 0.58 * | 99.50 ± 0.50 * |
| Conventional maize | 55.50 ± 3.69 | 19.50 ± 4.50 | 31.50 ± 2.63 | 63.50 ± 3.20 | 44.00 ± 3.92 | 29.50 ± 2.99 | |
| Stalk | Bt-(Cry1Ab+Vip3Aa) maize | 99.00 ± 0.58 * | 100.00 ± 0.00 * | 99.50 ± 0.50 * | 100.00 ± 0.00 * | 98.00 ± 0.82 * | 99.50 ± 0.50 * |
| Conventional maize | 47.00 ± 1.92 | 37.00 ± 5.80 | 41.50 ± 4.79 | 51.00 ± 4.12 | 42.50 ± 6.60 | 24.00 ± 4.83 | |
| Bract | Bt-(Cry1Ab+Vip3Aa) maize | 93.50 ± 2.22 * | 98.50 ± 0.96 * | 100.00 ± 0.00 * | 97.00 ± 0.58 * | 100.00 ± 0.00 * | 100.00 ± 0.00 * |
| Conventional maize | 76.00 ± 2.31 | 75.50 ± 1.89 | 85.00 ± 4.44 | 66.50 ± 7.63 | 87.50 ± 2.50 | 74.50 ± 3.30 | |
| Survey Year | Maize Variety | R3 | R5 | R6 | |||
|---|---|---|---|---|---|---|---|
| Numbers of Larvae per 100 Plants | Plant Damage Rates (%) | Numbers of Larvae per 100 Plants | Plant Damage Rates (%) | Numbers of Larvae per 100 Plants | Plant Damage Rates (%) | ||
| 2022 year | Bt-(Cry1Ab+Vip3Aa) maize | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Conventional maize | 208.90 ± 27.53 * | 60.30 ± 5.18 * | 104.80 ± 13.06 * | 75.80 ± 5.60 * | 422.10 ± 44.70 * | 94.40 ± 3.22 * | |
| 2023 year | Bt-(Cry1Ab+Vip3Aa) maize | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Conventional maize | 6.00 ± 2.15 * | 3.40 ± 0.73 * | 0.80 ± 0.61 | 0.4 ± 0.27 | 1.40 ± 0.99 | 0.80 ± 0.44 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Wang, W.; Kang, G.; Yang, X.; Wu, K. Toxic Effects of Bt-(Cry1Ab+Vip3Aa) Maize on Storage Pest Paralipsa gularis (Zeller). Toxins 2024, 16, 92. https://doi.org/10.3390/toxins16020092
Chen S, Wang W, Kang G, Yang X, Wu K. Toxic Effects of Bt-(Cry1Ab+Vip3Aa) Maize on Storage Pest Paralipsa gularis (Zeller). Toxins. 2024; 16(2):92. https://doi.org/10.3390/toxins16020092
Chicago/Turabian StyleChen, Shuang, Wenhui Wang, Guodong Kang, Xianming Yang, and Kongming Wu. 2024. "Toxic Effects of Bt-(Cry1Ab+Vip3Aa) Maize on Storage Pest Paralipsa gularis (Zeller)" Toxins 16, no. 2: 92. https://doi.org/10.3390/toxins16020092