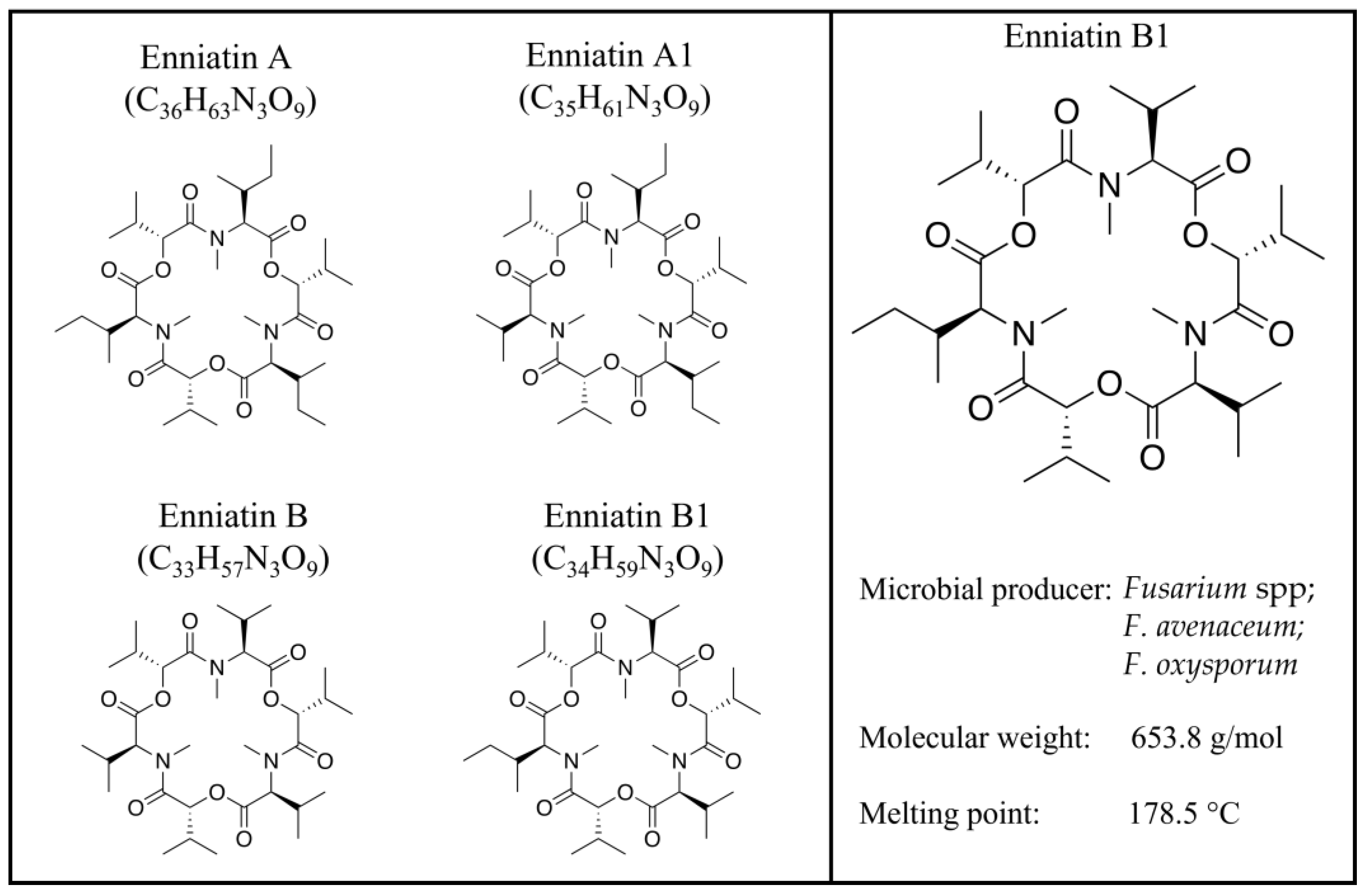
Enniatin B1: Emerging Mycotoxin and Emerging Issues


Abstract
:1. Introduction
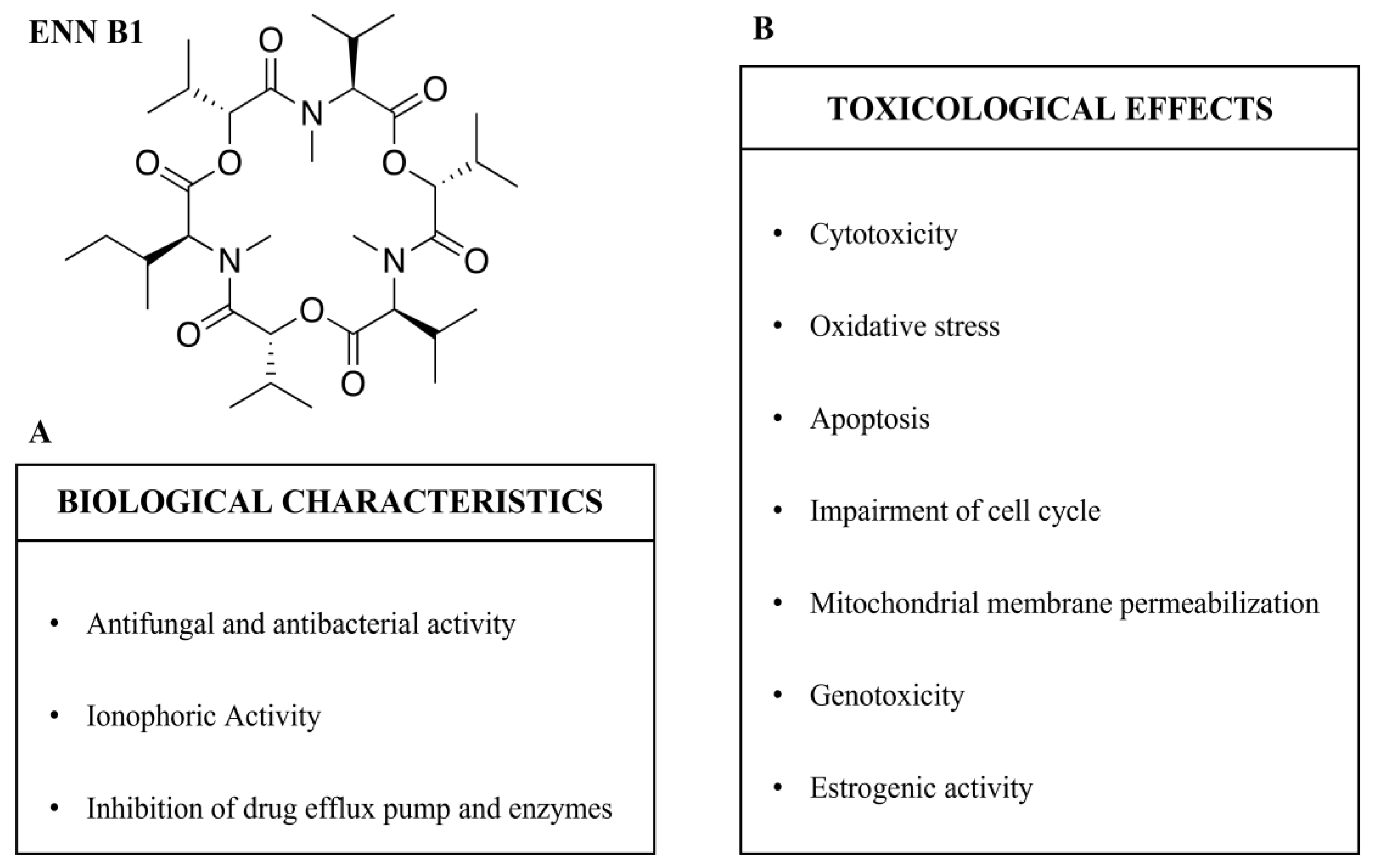
2. Enniatin B1
3. Biological Characteristics
3.1. Antifungal and Antibacterial Activity
3.2. Ionophoric Activity
3.3. Inhibition of Drug Efflux Pump and Enzymes

{kind=link}
{kind=link}
| Characteristics | Enniatins | Toward/Activity | Ref. |
|---|---|---|---|
| Antifungal activity | ENN B1 |
| [21] |
| ENN B1 |
| [27] | |
| ENN B1 |
| [29] | |
| Antibacterial activity | ENN B1 |
| [30] |
| ENN B1, B, B4 |
| [31] | |
| ENN B1 |
| [35] | |
| Ionophoric activity | ENN B1 |
| [39] |
| ENN B1 |
| [40] | |
| ENN B1 |
| [41] | |
| Inhibition of drug efflux pumps and enzymes | ENN B1 |
| [42,43] |
| ENN B1 |
| [35] | |
| ENN B1 |
| [35] |
4. Toxicological Effects
4.1. Cytotoxicity
4.2. Oxidative Stress
4.3. Apoptosis
4.4. Impairment of Cell Cycle
4.5. Mitochondrial Membrane Permeabilization
4.6. Genotoxicity
4.7. Estrogenic Activity
4.8. In Vivo Toxicity
| Toxicological Effects | Models/Cells | Concentrations and Exposure Times | Effects | Ref. |
|---|---|---|---|---|
| CaCo-2 cells | IC50 between 10.8 µM to 0.8 µM |
| [51] | |
| CaCo-2 cells | In total, 0.9 to 15.0 µM of binary, tertiary, and quaternary mixtures (ENN A, ENN A1, ENN B, ENN B1) |
| [45] | |
| HT-29 cells | IC50 between 16.6 µM and 3.7 µM |
| [28] | |
| IPEC-J2 cells | 5 µM |
| [55] | |
| Cytotoxicity | HepG2 cells | IC50 between 24.3 µM and 8.5 µM |
| [56] |
| MRC-5 cells | IC50 between 4.7 µM and 4.5 µM |
| [56] | |
| CHO-K1 cells | IC50 between 4.53 µM and 2.47 μM |
| [48] | |
| CHO-K1 cells | IC50 of 0.44 ± 0.15, ENN A1 + B mixture; IC50 of 0.97 ± 0.48, ENN A1 + B + B1 mixture. |
| [46] | |
| Insect SF-9 cells | IC50 of 6.6 µM |
| [34] | |
| PK-15 cells | IC50 of 41 µM |
| [35] | |
| CaCo-2 cells | 1.5 and 3 µM |
| [51] | |
| Oxidative stress | Mouse blastocysts | 1–10 µM |
| [60] |
| Wistar rats | Mixture of ENNs (ENN A, ENN A1, ENN B and ENN B1) |
| [49] | |
| Apoptosis | Pig embryos | 10, 25, and 50 μM |
| [47] |
| SH-SY5Y cells | 0.1 µM and 10 µM |
| [47] | |
| HepG2 cells | 1.5 µM and 3 µM |
| [48] | |
| Impairment of cell cycle | CaCo-2 cells | 0.9 µM to 15 µM |
| [51] |
| KB-3-1 cells | mixture of ENNs (3% ENN A, 20% ENN A1, 19% ENN B, and 54% ENN B1) |
| [48] | |
| Mouse blastocysts | 1–10 µM |
| [50] | |
| Porcine embryo | 10 µM to 50 µM |
| [47] | |
| Boar spermatozoa | 0.7 µM |
| [52] | |
| Mitochondrial membrane permeabilization | Rat liver mitochondria | Mixture of ENNs (3% ENN A, 20% ENN A1, 19% ENN B, and 54% of ENN B1) |
| [36] |
| Caco-2 cells | 1.5–3 µM |
| [51] | |
| Genotoxicity | Caco-2 cells | 1.5 µM to 3 µM |
| [51] |
| HEK 293T cells | 25 µM |
| [54] | |
| Estrogenic activity | VM7Luc4E2 cells | IC50 = 6.76 × 10−7 M |
| [53] |
| VM7Luc4E2 cells | IC50 = 8.13 × 10−7 M |
| [53] |
5. Conclusions and Future Challenges
Author Contributions
Funding
Conflicts of Interest
References
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. Emerging mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-Occurrence of Regulated, Masked and Emerging Mycotoxins and Secondary Metabolites in Finished Feed and Maize—An Extensive Survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekwomadu, T.I.; Akinola, S.A.; Mwanza, M. Fusarium Mycotoxins, Their Metabolites (Free, Emerging, and Masked), Food Safety Concerns, and Health Impacts. Int. J. Environ. Res. Public Health 2021, 18, 11741. [Google Scholar] [CrossRef] [PubMed]
- Jestoi, M. Emerging Fusarium-Mycotoxins Fusaproliferin, Beauvericin, Enniatins, And Moniliformin—A Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Gäumann, E.; Roth, S.; Ettlinger, L.; Plattner, P.l.A.; Nager, U. Enniatin, ein neues, gegen Mykobakterien wirksames Antibiotikum. Experientia 1947, 3, 202–203. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 2082. [Google Scholar]
- Maranghi, F.; Tassinari, R.; Narciso, L.; Tait, S.; Rocca, C.L.; Felice, G.D.; Butteroni, C.; Corinti, S.; Barletta, B.; Cordelli, E. In vivo toxicity and genotoxicity of beauvericin and enniatins. Combined approach to study in vivo toxicity and genotoxicity of mycotoxins beauvericin (BEA) and enniatin B (ENNB). EFSA Support. Publ. 2018, 15, 1406. [Google Scholar] [CrossRef] [Green Version]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the Mycotoxin Enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Escrivá, L.; Font, G.; Manyes, L.; Berrada, H. Studies on the Presence of Mycotoxins in Biological Samples: An Overview. Toxins 2017, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Křížová, L.; Dadáková, K.; Dvořáčková, M.; Kašparovský, T. Feedborne Mycotoxins Beauvericin and Enniatins and Livestock Animals. Toxins 2021, 13, 32. [Google Scholar] [CrossRef]
- Zingales, V.; Taroncher, M.; Martino, P.A.; Ruiz, M.J.; Caloni, F. Climate Change and Effects on Molds and Mycotoxins. Toxins 2022, 14, 445. [Google Scholar] [CrossRef]
- Jajić, I.; Dudaš, T.; Krstović, S.; Krska, R.; Sulyok, M.; Bagi, F.; Savić, Z.; Guljaš, D.; Stankov, A. Emerging Fusarium Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin in Serbian Maize. Toxins 2019, 11, 357. [Google Scholar] [CrossRef] [Green Version]
- De Colli, L.; De Ruyck, K.; Abdallah, M.F.; Finnan, J.; Mullins, E.; Kildea, S.; Spink, J.; Elliott, C.; Danaher, M. Natural Co-Occurrence of Multiple Mycotoxins in Unprocessed Oats Grown in Ireland with Various Production Systems. Toxins 2021, 13, 188. [Google Scholar] [CrossRef]
- García-Nicolás, M.; Arroyo-Manzanares, N.; Campillo, N.; Viñas, P. Cellulose-ferrite nanocomposite for monitoring enniatins and beauvericins in paprika by liquid chromatography and high-resolution mass spectrometry. Talanta 2021, 226, 122144. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, Q.; Zhang, Y.; Li, J.; Zhang, M.; Hu, X.; Li, F. Further data on the levels of emerging Fusarium mycotoxins in cereals collected from Tianjin, China. Food Addit. Contam. Part B 2021, 14, 74–80. [Google Scholar] [CrossRef]
- Giannioti, Z.; Albero, B.; Hernando, M.D.; Bontempo, L.; Pérez, R.A. Determination of Regulated and Emerging Mycotoxins in Organic and Conventional Gluten-Free Flours by LC-MS/MS. Toxins 2023, 15, 155. [Google Scholar] [CrossRef]
- Serrano, A.; Font, G.; Ruiz, M.; Ferrer, E. Co-occurrence and risk assessment of mycotoxins in food and diet from Mediterranean area. Food Chem. 2012, 135, 423–429. [Google Scholar] [CrossRef]
- EFSA. Mycotoxin mixtures in food and feed: Holistic, innovative, flexible risk assessment modelling approach. EFSA J. 2020, 17, 1757. [Google Scholar]
- Bertero, A.; Fossati, P.; Tedesco, D.E.A.; Caloni, F. Beauvericin and enniatins: In vitro intestinal effects. Toxins 2020, 12, 686. [Google Scholar] [CrossRef]
- Chiminelli, I.; Spicer, L.J.; Maylem, E.R.S.; Caloni, F. Emerging mycotoxins and reproductive effects in animals: A short review. J. Appl. Toxicol. 2022, 42, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- Caloni, F.; Fossati, P.; Anadón, A.; Bertero, A. Beauvericin: The beauty and the beast. Environ. Toxicol. Pharmacol. 2020, 75, 103349. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food—A review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Jayasinghe, L.; Abbas, H.K.; Jacob, M.R.; Herath, W.H.; Nanayakkara, N.P. N-Methyl-4-hydroxy-2-pyridinone analogues from Fusarium oxysporum. J. Nat. Prod. 2006, 6, 439–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, C.; Pinson-Gadais, L.; Richard-Forget, F. Fusarium mycotoxins enniatins: An updated review of their occurrence, the producing Fusarium species, and the abiotic determinants of their accumulation in crop harvests. J. Agric. Food Chem. 2020, 68, 4788–4798. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914. [Google Scholar] [CrossRef] [Green Version]
- Tsantrizos, Y.S.; Xu, X.J.; Sauriol, F.; Hynes, R.C. Novel quinazolinones and enniatins from Fusarium lateritium Nees. Can. J. Chem. 1993, 71, 1362–1367. [Google Scholar] [CrossRef]
- Meca, G.; Font, G.; Ruiz, M.J. Comparative cytotoxicity study of enniatins A, A1, A2, B, B1, B4 and J3 on Caco-2 cells, Hep-G2 and HT-29. Food Chem. Toxicol. 2011, 49, 2464–2469. [Google Scholar] [CrossRef]
- Firakova, S.; Proksa, B.; Šturdíková, M. Biosynthesis and biological activity of enniatins. Pharm.-Int. J. Pharm. Sci. 2006, 62, 563–568. [Google Scholar]
- Roig, M.; Meca, G.; Marin, R.; Ferrer, E.; Manes, J. Antibacterial activity of the emerging Fusarium mycotoxins enniatins A, A1, A2, B, B1, and B4 on probiotic microorganisms. Toxicon 2014, 85, 1–4. [Google Scholar] [CrossRef]
- Clark, T.N.; Carroll, M.; Ellsworth, K.; Guerrette, R.; Robichaud, G.A.; Johnson, J.A.; Gray, C.A. Antibiotic mycotoxins from an endophytic Fusarium acuminatum isolated from the medicinal plant Geum macrophyllum. Nat. Prod. Comm. 2018, 13, 10. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, M.; Zocher, R.; Haese, A. Enniatin production by Fusarium strains and its effect on potato tuber tissue. Appl. Environ. Microbiol. 1996, 62, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Strongman, D.B.; Strunz, G.M.; Giguere, P.; Yu, C.M.; Calhoun, L. Enniatins from Fusarium avenaceum isolated from balsam fir foliage and their toxicity to spruce budworm larvae, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). J. Chem. Ecol. 1988, 14, 753–764. [Google Scholar] [CrossRef]
- Fornelli, F.; Minervini, F.; Logrieco, A. Cytotoxicity of fungal metabolites to lepidopteran (Spodoptera frugiperda) cell line (SF-9). J. Invertebr. Pathol. 2004, 85, 74–79. [Google Scholar] [CrossRef]
- Sy-Cordero, A.A.; Pearce, C.J.; Oberlies, N.H. Revisiting the enniatins: A review of their isolation, biosynthesis, structure determination and biological activities. J. Antibiot. 2012, 65, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Tonshin, A.A.; Teplova, V.V.; Andersson, M.A.; Salkinoja-Salonen, M.S. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by affecting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology 2010, 276, 49–57. [Google Scholar] [CrossRef]
- Kamyar, M.R.; Rawnduzi, P.; Studenik, C.R.; Kouri, K.; Lemmens-Gruber, R. Investigation of the electrophysiological properties of enniatins. Arch. Biochem. Biophys. 2004, 429, 215–223. [Google Scholar] [CrossRef]
- Kamyar, M.R.; Kouri, K.; Rawnduzi, P.; Studenik, C.R.; Lemmens-Gruber, R. Effects of moniliformin in presence of cyclohexadepsipeptides on isolated mammalian tissue and cells. Toxicol. Vitr. 2006, 20, 1284–1291. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Ivanova, L.; Solhaug, A.; Fæste, C.K. Enniatin B 1-induced lysosomal membrane permeabilization in mouse embryonic fibroblasts. Mycotoxin Res. 2020, 36, 23–30. [Google Scholar] [CrossRef]
- Pérez-Fuentes, N.; Alvarino, R.; Alfonso, A.; González-Jartín, J.; Gegunde, S.; Vieytes, M.R.; Botana, L.M. Enniatins A1 and B1 alter calcium homeostasis of neuronal cells leading to apoptotic death. Food Chem. Toxicol. 2022, 168, 113361. [Google Scholar] [CrossRef]
- Stein, U.; Walther, W. Reversal of ABC transporter-dependent multidrug resistance in cancer: A realistic option? Am. J. Cancer 2006, 5, 285–297. [Google Scholar] [CrossRef]
- Hiraga, K.; Yamamoto, S.; Fukuda, H.; Hamanaka, N.; Oda, K. Enniatin has a new function as an inhibitor of the Pdr5p, one of the ABC transporter in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2005, 328, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Hiraga, K.; Abiko, A.; Hamanaka, N.; Oda, K. A new function of isonitrile as an inhibitor of the Pdr5p multidrug ABC transporter in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2005, 330, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, L.; Uhlig, S.; Devreese, M.; Croubels, S.; Fæste, C.K. Biotransformation of the mycotoxin enniatin B1 in pigs: A comparative in vitro and in vivo approach. Food Chem. Toxicol. 2017, 105, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Font, G.; Ruiz, M.J. Interaction effects of Fusarium enniatins (A, A1, B and B1) combinations on in vitro cytotoxicity of Caco-2 cells. Toxicol. Vitr. 2014, 28, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Fernández-Franzón, M.; Font, G.; Ruiz, M.J. Toxicity evaluation of individual and mixed enniatins using an in vitro method with CHO-K1 cells. Toxicol. Vitr. 2013, 27, 672–680. [Google Scholar] [CrossRef]
- Wang, X.; Sun, M.; Li, J.; Song, X.; He, H.; Huan, Y. Melatonin protects against defects induced by Enniatin B1 during porcine early embryo development. Aging 2021, 13, 5553. [Google Scholar] [CrossRef]
- Juan-García, A.; Juan, C.; Bind, M.A.; Engert, F. Study of locomotion response and development in zebrafish (Danio rerio) embryos and larvae exposed to enniatin A, enniatin B, and beauvericin. Sci. Total Environ. 2021, 777, 146075. [Google Scholar] [CrossRef]
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Frangiamone, M.; Manyes, L. Transcriptional changes after enniatins A, A1, B and B1 ingestion in rat stomach, liver, kidney and lower intestine. Foods 2021, 10, 1630. [Google Scholar] [CrossRef]
- Huang, C.H.; Wang, F.T.; Chan, W.H. Enniatin B1 exerts embryotoxic effects on mouse blastocysts and induces oxidative stress and immunotoxicity during embryo development. Environ. Toxicol. 2019, 34, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M.J. eactive oxygen species involvement in apoptosis and mitochondrial damage in Caco-2 cells induced by enniatins A, A1, B and B1. Toxicol. Lett. 2013, 222, 36–44. [Google Scholar] [CrossRef]
- Hoornstra, D.; Andersson, M.A.; Mikkola, R.; Salkinoja-Salonen, M.S. A new method for in vitro detection of microbially produced mitochondrial toxins. Toxicol. Vitr. 2003, 17, 745–751. [Google Scholar] [CrossRef]
- Park, Y.; Lee, H.S. Cyclic depsipeptide mycotoxin exposure may cause human endocrine disruption: Evidence from OECD in vitro stably transfected transcriptional activation assays. Reprod. Toxicol. 2021, 100, 52–59. [Google Scholar] [CrossRef]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Ruml, T. In silico and in vitro studies of mycotoxins and their cocktails; Their toxicity and its mitigation by silibinin pre-treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef] [Green Version]
- Springler, A.; Vrubel, G.J.; Mayer, E.; Schatzmayr, G.; Novak, B. Effect of Fusarium-derived metabolites on the barrier integrity of differentiated intestinal porcine epithelial cells (IPEC-J2). Toxins 2016, 8, 345. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, L.; Denisov, I.G.; Grinkova, Y.V.; Sligar, S.G.; Fæste, C. Biotransformation of the Mycotoxin Enniatin B1 by CYP P450 3A4 and Potential for Drug-Drug Interactions. Metabolites 2019, 9, 158. [Google Scholar] [CrossRef] [Green Version]
- Fraeyman, S.; Devreese, M.; Antonissen, G.; De Baere, S.; Rychlik, M.; Croubels, S. Comparative Oral Bioavailability, Toxicokinetics, and Biotransformation of Enniatin B1 and Enniatin B in Broiler Chickens. J. Agric. Food Chem. 2016, 64, 7259–7726. [Google Scholar] [CrossRef]
- Novak, B.; Rainer, V.; Sulyok, M.; Haltrich, D.; Schatzmayr, G.; Mayer, E. Twenty-eight fungal secondary metabolites detected in pig feed samples: Their occurrence, relevance and cytotoxic effects in vitro. Toxins 2019, 11, 537. [Google Scholar] [CrossRef] [Green Version]
- Kolf-Clauw, M.; Sassahara, M.; Lucioli, J.; Rubira-Gerez, J.; Alassane-Kpembi, I.; Lyazhri, F.; Oswald, I.P. The emerging mycotoxin, enniatin B1, down-modulates the gastrointestinal toxicity of T-2 toxin in vitro on intestinal epithelial cells and ex vivo on intestinal explants. Arch. Toxicol. 2013, 87, 2233–2241. [Google Scholar] [CrossRef]
- Wätjen, W.; Debbab, A.; Hohlfeld, A.; Chovolou, Y.; Kampkötter, A.; Edrada, R.A.; Proksch, P. Enniatins A1, B and B1 from an endophytic strain of Fusarium tricinctum induce apoptotic cell death in H4IIE hepatoma cells accompanied by inhibition of ERK phosphorylation. Mol. Nutr. Food Res. 2009, 53, 431–440. [Google Scholar] [CrossRef]
- Manyes, L.; Escrivá, L.; Ruiz, M.J.; Juan-García, A. Beauvericin and enniatin B effects on a human lymphoblastoid Jurkat T-cell model. Food Chem. Toxic. 2018, 115, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; Broekaert, N.; De Mil, T.; Fraeyman, S.; De Backer, P.; Croubels, S. Pilot toxicokinetic study and absolute oral bioavailability of the Fusarium mycotoxin enniatin B1 in pigs. Food Chem. Toxic. 2014, 63, 161–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callebaut, F.; Tangni, E.K.; Debongnie, P.; Stals, E.; Huybrechts, B.; Waegeneers, N.; Delezie, E.; Van Pamel, E.; Daeseleire, E. Carry-Over of Mycotoxins to Animal Products: Case Study Poultry; Scientifec Report 211/212 CODA-CERVA; Centrum voor Onderzoek in Diergeneeskunde en Agrochemie-Centre d’Étude et de Recherches Vétérinaires et Agrochemiques: Brussels, Belgium, 2011; pp. 141–144. [Google Scholar]
- Escrivá, L.; Font, G.; Manyes, L. Quantitation of enniatins in biological samples of Wistar rats after oral administration by LC-MS/MS. Toxicol. Mech. Methods 2015, 25, 552–558. [Google Scholar] [PubMed]
- Parikka, P.; Hakala, K.; Tiilikkala, K. Expected shifts in Fusarium species’ composition on cereal grain in Northern Europe due to climatic change. Food Addit. Contam. Part A 2012, 29, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Milicevic, D.; Lakicevic, B.; Petronijevic, R.; Petrovic, Z.; Jovanovic, J.; Stefanovic, S.; Jankovic, S. Climate change: Impact on mycotoxins incidence and food safety. Theory Pract. Meat Process. 2019, 4, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Carrasco, Y.; Narváez, A.; Izzo, L.; Gaspari, A.; Graziani, G.; Ritieni, A. Biomonitoring of Enniatin B1 and Its Phase I Metabolites in Human Urine: First Large-Scale Study. Toxins 2020, 12, 415. [Google Scholar] [CrossRef]
- Jestoi, M.; Rokka, M.; Järvenpää, E.; Peltonen, K. Determination of Fusarium mycotoxins beauvericin and enniatins (A, A1, B, B1) in eggs of laying hens using liquid chromatography-tandem mass spectrometry (LC–MS/MS). Food Chem. 2009, 115, 1120–1127. [Google Scholar] [CrossRef]
- Rossi, F.; Gallo, A.; Bertuzzi, T. Emerging mycotoxins in the food chain. Mediterr. J. Nutr. Metab. 2020, 13, 7–27. [Google Scholar] [CrossRef]
- Tolosa, J.; Font, G.; Mañes, J.; Ferrer, E. Natural occurrence of Fusarium mycotoxins in aquaculture fish food. Rev. Toxicol. 2013, 30, 193–197. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, B.; Spicer, L.J.; Caloni, F. Enniatin B1: Emerging Mycotoxin and Emerging Issues. Toxins 2023, 15, 383. https://doi.org/10.3390/toxins15060383
De Felice B, Spicer LJ, Caloni F. Enniatin B1: Emerging Mycotoxin and Emerging Issues. Toxins. 2023; 15(6):383. https://doi.org/10.3390/toxins15060383
Chicago/Turabian StyleDe Felice, Beatrice, Leon J. Spicer, and Francesca Caloni. 2023. "Enniatin B1: Emerging Mycotoxin and Emerging Issues" Toxins 15, no. 6: 383. https://doi.org/10.3390/toxins15060383