Genomic Insights into Virulence Factors and Multi-Drug Resistance in Clostridium perfringens IRMC2505A

 , ,
, ,  and
and 
Abstract
:1. Introduction
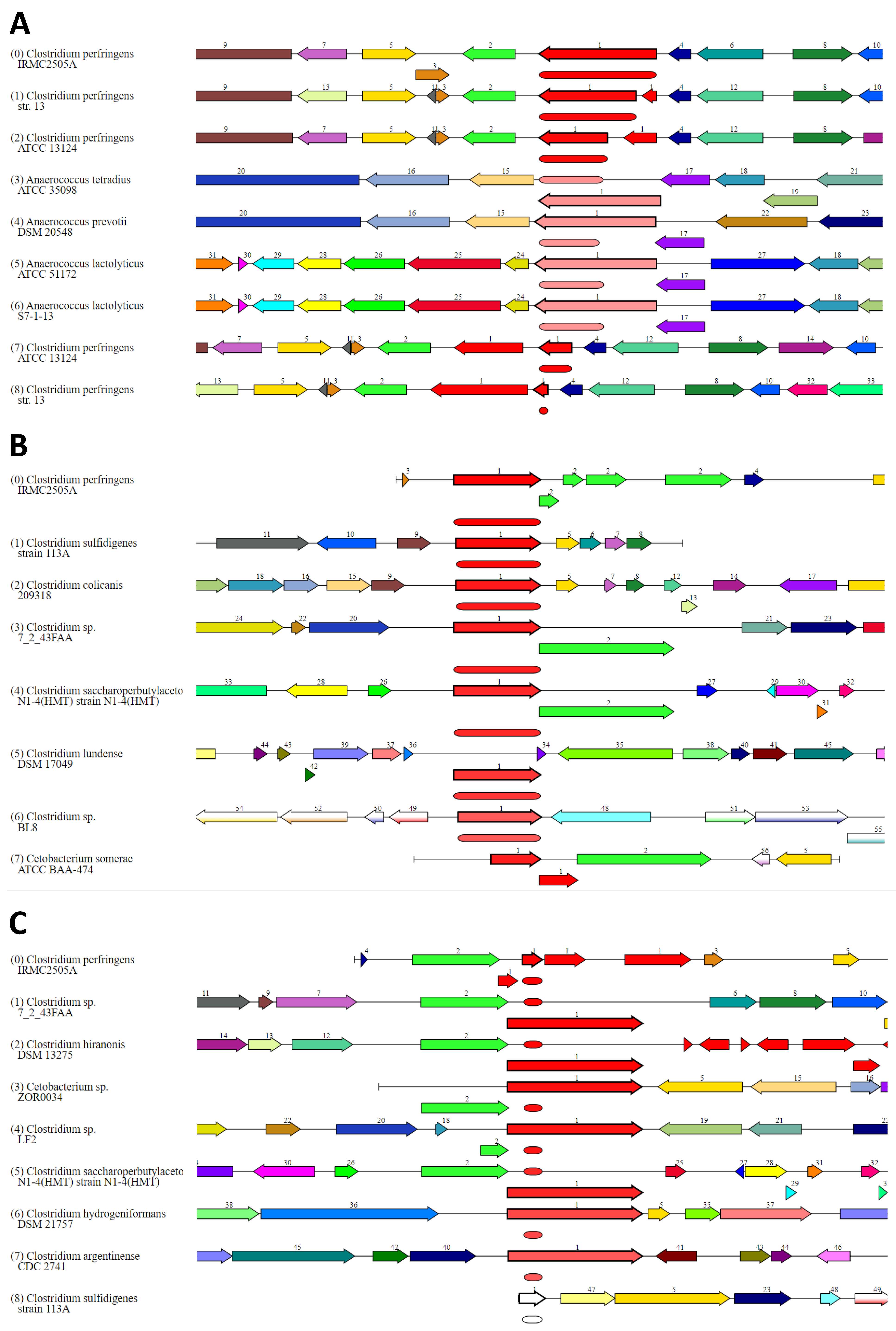
2. Results
Antimicrobial Resistance Genes
3. Discussion
3.1. Phylogenetic Analyses
3.2. Antimicrobial Resistance
3.3. Toxigenicity
4. Materials and Methods
4.1. Isolation and DNA Extraction of C. perfringens IRMC2505A
4.2. Virulence Genes Identification Using Whole Genome Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ueda, K.; Kawahara, K.; Kimoto, N.; Yamaguchi, Y.; Yamada, K.; Oki, H.; Yoshida, T.; Matsuda, S.; Matsumoto, Y.; Motooka, D.; et al. Analysis of the complete genome sequences of Clostridium perfringens strains harbouring the binary enterotoxin BEC gene and comparative genomics of pCP13-like family plasmids. BMC Genom. 2022, 23, 226. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ma, M.; Uzal, F.A.; McClane, B.A. Host cell-induced signaling causes Clostridium perfringens to upregulate production of toxins important for intestinal infections. Gut. Microbes 2013, 5, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Kiu, R.; Caim, S.; Alexander, S.; Pachori, P.; Hall, L.J. Probing genomic aspects of the multi-host pathogen Clostridium perfringens reveals significant pangenome diversity, and a diverse array of virulence factors. Front. Microbiol. 2017, 8, 2485. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Elbourne, L.D.H.; Tetu, S.G.; Melville, S.B.; Rood, J.I.; Paulsen, I.T. Genomic analyses of Clostridium perfringens isolates from five toxinotypes. Res. Microbiol. 2015, 166, 255–263. [Google Scholar] [CrossRef]
- Sakurai, J.; Nagahama, M.; Oda, M. Clostridium perfringens Alpha-Toxin: Characterization and Mode of Action. J. Biochem. 2004, 136, 569–574. [Google Scholar] [CrossRef]
- Kubota, H.; Sakai, T.; Gawad, A.; Makino, H.; Akiyama, T.; Ishikawa, E.; Oishi, K. Development of TaqMan-Based Quantitative PCR for Sensitive and Selective Detection of Toxigenic Clostridium difficile in Human Stools. PLoS ONE 2014, 9, e111684. [Google Scholar] [CrossRef]
- Miyamoto, K.; Li, J.; McClane, B.A. Enterotoxigenic Clostridium perfringens: Detection and identification. Microbes Environ. 2012, 27, 343–349. [Google Scholar] [CrossRef]
- Bennett, S.D.; Walsh, K.A.; Gould, L.H. Foodborne Disease Outbreaks Caused by Bacillus cereus, Clostridium perfringens, and Staphylococcus aureus—United States, 1998–2008. Clin. Infect. Dis. 2013, 57, 425–433. [Google Scholar] [CrossRef]
- Martin, J.S.H.; Monaghan, T.M.; Wilcox, M.H. Clostridium difficile infection: Epidemiology, diagnosis and understanding transmission. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 206–216. [Google Scholar] [CrossRef]
- Eyre, D.W.; Walker, A.S. Clostridium difficile surveillance: Harnessing new technologies to control transmission. Expert Rev. Anti-Infect. Ther. 2014, 11, 1193–1205. [Google Scholar] [CrossRef]
- C. perfringens|CDC. Available online: https://www.cdc.gov/foodsafety/diseases/clostridium-perfringens.html (accessed on 5 February 2023).
- Omer, S.A.; Al-Olayan, E.M.; Babiker, S.E.H.; Aljulaifi, M.Z.; Alagaili, A.N.; Mohammed, O.B. Genotyping of Clostridium perfringens Isolates from Domestic Livestock in Saudi Arabia. BioMed Res. Int. 2020, 2020, 9035341. [Google Scholar] [CrossRef] [PubMed]
- Bendary, M.M.; Abd El-Hamid, M.I.; El-Tarabili, R.M.; Hefny, A.A.; Algendy, R.M.; Elzohairy, N.A.; Ghoneim, M.M.; Al-Sanea, M.M.; Nahari, M.H.; Moustafa, W.H. Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes. Biology 2022, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Collins, D.A.; Imwattana, K.; Knight, D.R.; Perumalsamy, S.; Hain-Saunders, N.M.R.; Putsathit, P.; Speers, D.; Riley, T.V. Whole-genome sequencing links Clostridium (Clostridioides) difficile in a single hospital to diverse environmental sources in the community. J. Appl. Microbiol. 2022, 133, 1156–1168. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.; Nguyen, S.V.; MacOri, G.; Bolton, D.; McMullan, G.; Drudy, D.; Fanning, S. Clostridioides difficile as a Potential Pathogen of Importance to One Health: A Review. Foodborne Pathog. Dis. 2022, 19, 806–816. [Google Scholar] [CrossRef]
- Shimizu, T.; Ohtani, K.; Hirakawa, H.; Ohshima, K.; Yamashita, A.; Shiba, T.; Ogasawara, N.; Hattori, M.; Kuhara, S.; Hayashi, H. Complete genome sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Natl. Acad. Sci. USA 2002, 99, 996–1001. [Google Scholar] [CrossRef]
- Feng, Y.; Fan, X.; Zhu, L.; Yang, X.; Liu, Y.; Gao, S.; Jin, X.; Liu, D.; Ding, J.; Guo, Y.; et al. Phylogenetic and genomic analysis reveals high genomic openness and genetic diversity of Clostridium perfringens. Microb. Genom. 2020, 6, e000441. [Google Scholar] [CrossRef]
- Arefiev, V.; Kovalenko, G.; Frant, M.; Chumachenko, T.; Polyvianna, Y.; Pivnenko, S.; Bolotin, V.; Mayboroda, O.; Solodiankin, O.; Tarasov, O.; et al. Complete Genome Sequence of Salmonella enterica subsp. enterica Serovar Kottbus Strain Kharkiv, Isolated from a Commercial Pork Production Facility in Ukraine. Microbiol. Resour. Announc. 2020, 9, e01171-20. [Google Scholar] [CrossRef]
- Sloan, J.; McMurry, L.M.; Lyras, D.; Levy, S.B.; Rood, J.I. The Clostridium perfringens Tet P determinant comprises two overlapping genes: TetA(P), which mediates active tetracycline efflux, and tetB(P), which is related to the ribosomal protection family of tetracycline-resistance determinants. Mol. Microbiol. 1994, 11, 403–415. [Google Scholar] [CrossRef]
- Ernst, C.M.; Staubitz, P.; Mishra, N.N.; Yang, S.-J.; Hornig, G. The Bacterial Defensin Resistance Protein MprF Consists of Separable Domains for Lipid Lysinylation and Antimicrobial Peptide Repulsion. PLoS Pathog. 2009, 5, 1000660. [Google Scholar] [CrossRef]
- Prezioso, S.M.; Brown, N.E.; Goldberg, J.B. Elfamycins: Inhibitors of elongation factor-Tu. Mol. Microbiol. 2017, 106, 22–34. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Sani, N.A. Antibiotic resistance and burden of foodborne diseases in developing countries. Futur. Sci. OA 2016, 2, FSO139. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Rooney, A.P.; Hecht, D.W.; Li, J.; McClane, B.A.; Nayak, R.; Paine, D.D.; Rafii, F. Phenotypic and genotypic characterization of tetracycline and minocycline resistance in Clostridium perfringens. Arch. Microbiol. 2010, 192, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, H.; Fathollahi, M.; Abiri, R.; Kadivarian, S.; Rostamian, M.; Alvandi, A. A worldwide systematic review and metaanalysis of bacteria related to antibioticassociated diarrhea in hospitalized patients. PLoS ONE 2021, 16, e0260667. [Google Scholar] [CrossRef]
- Banawas, S.S. Systematic Review and Meta-Analysis on the Frequency of Antibiotic-Resistant Clostridium Species in Saudi Arabia. Antibiotics 2022, 11, 1165. [Google Scholar] [CrossRef]
- Fayez, M.; Elsohaby, I.; Al-Marri, T.; Zidan, K.; Aldoweriej, A.; El-Sergany, E.; Elmoslemany, A. Genotyping and antimicrobial susceptibility of Clostridium perfringens isolated from dromedary camels, pastures and herders. Comp. Immunol. Microbiol. Infect. Dis. 2020, 70, 101460. [Google Scholar] [CrossRef] [PubMed]
- Fayez, M.; El-Ghareeb, W.R.; Elmoslemany, A.; Alsunaini, S.J.; Alkafafy, M.; Alzahrani, O.M.; Mahmoud, S.F.; Elsohaby, I. Genotyping and Antimicrobial Susceptibility of Clostridium perfringens and Clostridioides difficile in Camel Minced Meat. Pathogens 2021, 10, 1640. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Abdelrahim, A.M.; Radomski, N.; Delannoy, S.; Djellal, S.; Le Négrate, M.; Hadjab, K.; Fach, P.; Hennekinne, J.A.; Mistou, M.Y.; Firmesse, O. Large-Scale genomic analyses and toxinotyping of Clostridium perfringens implicated in foodborne outbreaks in France. Front. Microbiol. 2019, 10, 777. [Google Scholar] [CrossRef]
- Francis Borgio, J.; Rasdan, A.S.; Sonbol, B.; Alhamid, G.; Almandil, N.B.; Azeez, S.A. Emerging status of multidrug-resistant bacteria and fungi in the arabian peninsula. Biology 2021, 10, 1144. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, I.; Chang, A.; Ebeling, C.; Gremse, M.; Heldt, C.; Huhn, G.; Schomburg, D. BRENDA, the enzyme database: Updates and major new developments. Nucleic Acids Res. 2004, 32, D431–D433. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Consortium, T.; Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Michael Cherry, J.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; et al. Gene Ontology: Tool for the unification of biology NIH Public Access Author Manuscript. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.J.; Gerdes, S.; Olsen, G.J.; Olson, R.; Pusch, G.D.; Shukla, M.; Vonstein, V.; Wattam, A.R.; Yoo, H. PATtyFams: Protein families for the microbial genomes in the PATRIC database. Front. Microbiol. 2016, 7, 118. [Google Scholar] [CrossRef]
- Overbeek, R.; Begley, T.; Butler, R.M.; Choudhuri, J.V.; Chuang, H.Y.; Cohoon, M.; de Crécy-Lagard, V.; Diaz, N.; Disz, T.; Edwards, R.; et al. The Subsystems Approach to Genome Annotation and its Use in the Project to Annotate 1000 Genomes. Nucleic Acids Res. 2005, 33, 5691–5702. [Google Scholar] [CrossRef]
- Saier, M.H.; Reddy, V.S.; Tsu, B.V.; Ahmed, M.S.; Li, C.; Moreno-Hagelsieb, G. The Transporter Classification Database (TCDB): Recent advances. Database Issue Publ. 2016, 44, D372–D379. [Google Scholar] [CrossRef]
- Mao, C.; Abraham, D.; Wattam, A.R.; Wilson, M.J.C.; Shukla, M.; Yoo, H.S.; Sobral, B.W. Curation, integration and visualization of bacterial virulence factors in PATRIC. Bioinformatics 2015, 31, 252–258. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis-10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Zhu, F.; Han, B.; Kumar, P.; Liu, X.; Ma, X.; Wei, X.; Huang, L.; Guo, Y.; Han, L.; Zheng, C.; et al. Update of TTD: Therapeutic Target Database. Nucleic Acids Res. 2010, 38, D787–D791. [Google Scholar] [CrossRef]
- Law, V.; Knox, C.; Djoumbou, Y.; Jewison, T.; Guo, A.C.; Liu, Y.; MacIejewski, A.; Arndt, D.; Wilson, M.; Neveu, V.; et al. DrugBank 4.0: Shedding new light on drug metabolism. Nucleic Acids Res. 2014, 42, D1091–D1097. [Google Scholar] [CrossRef] [PubMed]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2016, 45, D566–D573. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| S. No. | Antimicrobial Resistant Mechanism | Genes |
|---|---|---|
| 1 | Antibiotic inactivation enzyme | NimB |
| 2 | Antibiotic target in susceptible species | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p |
| 3 | Antibiotic target protection protein | TetB(P) |
| 4 | Antibiotic target replacement protein | FabV |
| 5 | Efflux pump conferring antibiotic resistance | TetA(P) |
| 6 | Gene conferring resistance via absence | GidB |
| 7 | Protein altering cell wall charge conferring antibiotic resistance | GdpD, MprF, PgsA |
| 8 | Protein involved in antibiotic sequestration | FabK-like |
| Source | Gene | Product/Function | Classification | Subject Coverage | Query Coverage | Identity | E-Value | |
|---|---|---|---|---|---|---|---|---|
| 1 | NDARO/CARD | tetB(P) | Tetracycline resistance, ribosomal protection type ≥ TetB(P) | antibiotic target protection protein, tetracycline-resistant gene | 13 | 91 | 95 | 5 × 10−40 |
| 2 | CARD/NDARO | tetB(P) | Tetracycline resistance, ribosomal protection type ≥ TetB(P) | antibiotic target protection protein, tetracycline-resistant gene | 48 | 99 | 98 | 1 × 10−180 |
| 3 | CARD/PATRIC | EF-Tu | Translation elongation factor Tu | antibiotic resistant gene variant or mutant, elfamycin-resistant gene/antibiotic target in susceptible species | 6 | 96 | 83 | 7 × 10−3 |
| 4 | NDARO/CARD/PATRIC | tetA(P) | Tetracycline resistance, MFS efflux pump ≥ TetA(P) | efflux pump conferring antibiotic resistance | 100 | 100 | 98 | 1 × 10−234 |
| 5 | CARD | mprF | putative membrane protein | antibiotic target modifying enzyme, peptide antibiotic resistance gene | 100 | 100 | 95 | 0.0 |
| Source | Source ID | Source Organism | Gene | Product | Classification | Subject Coverage | Query Coverage | Identity | E-Value |
|---|---|---|---|---|---|---|---|---|---|
| Victors | 15676067 | Neisseria meningitidis MC58 | tufA | Translation elongation factor Tu | 5 | 92 | 86 | 3 × 10−3 | |
| VFDB | VFG002285 | Clostridium perfringens ATCC 13124 | nanH | Sialidase (EC 3.2.1.18) | Exoenzyme, Carbohydrate-active enzyme, Sialidase | 100 | 100 | 98 | 1 × 10−226 |
| Victors | 110801372 | Clostridium perfringens ATCC 13124 | virS | Hypothetical protein | 99 | 100 | 95 | 1 × 10−243 | |
| Victors | 18309145 | Clostridium perfringens str. 13 | pfoA | Thiol-activated cytolysin | 92 | 88 | 87 | 1 × 10−243 | |
| Victors | 15676067 | Neisseria meningitidis MC58 | tufA | Translation elongation factor Tu | 5 | 92 | 86 | 3 × 10−3 | |
| VFDB | VFG002278 | Clostridium perfringens str. 13 | nagI | Protein O-GlcNAcase (EC 3.2.1.169) | Exoenzyme, Carbohydrate-active enzyme, Hyaluronidase | 100 | 100 | 96 | 0.0 |
| Victors | 29376182 | Enterococcus faecalis V583 | EF1623 | Ethanolamine utilization protein similar to PduA/PduJ | 91 | 91 | 92 | 4 × 10−38 | |
| VFDB | VFG002274 | Clostridium perfringens str. 13 | plc | Broad-substrate range phospholipase C (EC 3.1.4.3) | Toxin, Zinc-metallophospholipase C | 100 | 100 | 99 | 1 × 10−242 |
| VFDB | VFG002282 | Clostridium perfringens str. 13 | cloSI | Hypothetical protein | Exoenzyme, Cysteine endopeptidase | 99 | 100 | 97 | 1 × 10−306 |
| VFDB | VFG002279 | Clostridium perfringens str. 13 | nagJ | Protein O-GlcNAcase (EC 3.2.1.169) | Exoenzyme, Carbohydrate-active enzyme, Hyaluronidase | 100 | 100 | 98 | 0.0 |
| Victors | 18309018 | Clostridium perfringens str. 13 | plc | Broad-substrate range phospholipase C (EC 3.1.4.3) | 100 | 100 | 99 | 1 × 10−242 | |
| VFDB | VFG002281 | Clostridium perfringens str. 13 | nagL | Hypothetical protein | Exoenzyme, Carbohydrate-active enzyme, Hyaluronidase | 100 | 100 | 95 | 0.0 |
| VFDB | VFG002283 | Clostridium perfringens str. 13 | nanI | Sialidase (EC 3.2.1.18) | Exoenzyme, Carbohydrate-active enzyme, Sialidase | 100 | 100 | 99 | 0.0 |
| VFDB | VFG002276 | Clostridium perfringens str. 13 | colA | Microbial collagenase (EC 3.4.24.3) | Exoenzyme, Collagenase | 100 | 100 | 98 | 0.0 |
| VFDB | VFG002284 | Clostridium perfringens str. 13 | nanJ | Sialidase (EC 3.2.1.18) | Exoenzyme, Carbohydrate-active enzyme, Sialidase | 100 | 100 | 95 | 0.0 |
| VFDB | VFG002275 | Clostridium perfringens str. 13 | pfoA | Thiol-activated cytolysin | Toxin, Membrane-damaging, Pore-forming, Channel-forming involving beta-sheet-containing toxin (beta-barrel), Cholesterol-dependent cytolysin | 92 | 88 | 87 | 1 × 10−243 |
| VFDB | VFG002277 | Clostridium perfringens str. 13 | nagH | Hyaluronoglucosaminidase (EC 3.2.1.35) | Exoenzyme, Carbohydrate-active enzyme, Hyaluronidase | 100 | 100 | 97 | 0.0 |
| Template | Gene Function | Genome | Score | Expected | Template Length | Template Identity | Template Coverage | Query Identity | Query Coverage | Depth | q-Value | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CARD|ABG86067.1 | Clostridium perfringens mprF | $ | 1395 | 1 | 1710 | 93.8 | 100 | 93.8 | 100 | 1 | 1390.34 | 1.00 × 10−26 |
| CARD|AAA20116.1 | tetA(P) | # | 1218 | 1 | 1263 | 98.73 | 100 | 98.73 | 100 | 1 | 1214.3 | 1.00 × 10−26 |
| VFDB|VFG002274 | phospholipase C | * | 1164 | 7 | 1197 | 99 | 100 | 99 | 100 | 1 | 1140.8 | 1.00 × 10−26 |
| VFDB|VFG002275 | perfringolysin O | * | 915 | 9 | 1503 | 86.56 | 98.27 | 88.08 | 101.76 | 0.98 | 885.75 | 1.00 × 10−26 |
| VFDB|VFG002276 | collagenase | * | 3177 | 19 | 3315 | 98.58 | 100 | 98.58 | 100 | 1 | 3117.65 | 1.00 × 10−26 |
| VFDB|VFG002277 | hyaluronidase | * | 4467 | 27 | 4887 | 97.11 | 100 | 97.11 | 100 | 1 | 4384.16 | 1.00 × 10−26 |
| VFDB|VFG002278 | hyaluronidase | * | 3537 | 23 | 3894 | 96.92 | 100 | 96.92 | 100 | 1 | 3468.34 | 1.00 × 10−26 |
| VFDB|VFG002279 | hyaluronidase | * | 2769 | 18 | 3006 | 97.34 | 100 | 97.34 | 100 | 1 | 2714.3 | 1.00 × 10−26 |
| VFDB|VFG002281 | hyaluronidase | * | 2976 | 20 | 3384 | 95.95 | 100 | 95.95 | 100 | 1 | 2914.96 | 1.00 × 10−26 |
| VFDB|VFG002282 | alpha-clostripain | * | 1459 | 10 | 1575 | 97.65 | 100 | 97.65 | 100 | 1 | 1428.83 | 1.00 × 10−26 |
| VFDB|VFG002283 | exo-alpha-sialidase | * | 1995 | 13 | 2085 | 98.51 | 100 | 98.51 | 100 | 1 | 1955.87 | 1.00 × 10−26 |
| VFDB|VFG002284 | exo-alpha-sialidase | * | 3206 | 21 | 3522 | 97.02 | 100 | 97.02 | 100 | 1 | 3143.05 | 1.00 × 10−26 |
| VFDB|VFG002285 | sialidase | ‡ | 1026 | 7 | 1149 | 96.34 | 100 | 96.34 | 100 | 1 | 1003.63 | 1.00 × 10−26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AlJindan, R.; AlEraky, D.M.; Farhat, M.; Almandil, N.B.; AbdulAzeez, S.; Borgio, J.F. Genomic Insights into Virulence Factors and Multi-Drug Resistance in Clostridium perfringens IRMC2505A. Toxins 2023, 15, 359. https://doi.org/10.3390/toxins15060359
AlJindan R, AlEraky DM, Farhat M, Almandil NB, AbdulAzeez S, Borgio JF. Genomic Insights into Virulence Factors and Multi-Drug Resistance in Clostridium perfringens IRMC2505A. Toxins. 2023; 15(6):359. https://doi.org/10.3390/toxins15060359
Chicago/Turabian StyleAlJindan, Reem, Doaa M. AlEraky, Maha Farhat, Noor B. Almandil, Sayed AbdulAzeez, and Jesu Francis Borgio. 2023. "Genomic Insights into Virulence Factors and Multi-Drug Resistance in Clostridium perfringens IRMC2505A" Toxins 15, no. 6: 359. https://doi.org/10.3390/toxins15060359