Investigation on Fermentation Characteristics and Microbial Communities of Wheat Straw Silage with Different Proportion Artemisia argyi


Abstract
:1. Introduction
2. Results and Discussion
2.1. Characteristics of Fresh Materials before Storage Fermentation
2.2. Characteristics of Different Silage Treatments during Storage Fermentation and Aerobic Exposure
2.2.1. Fermentation Quality
2.2.2. Chemical Composition
2.2.3. Microbial Population
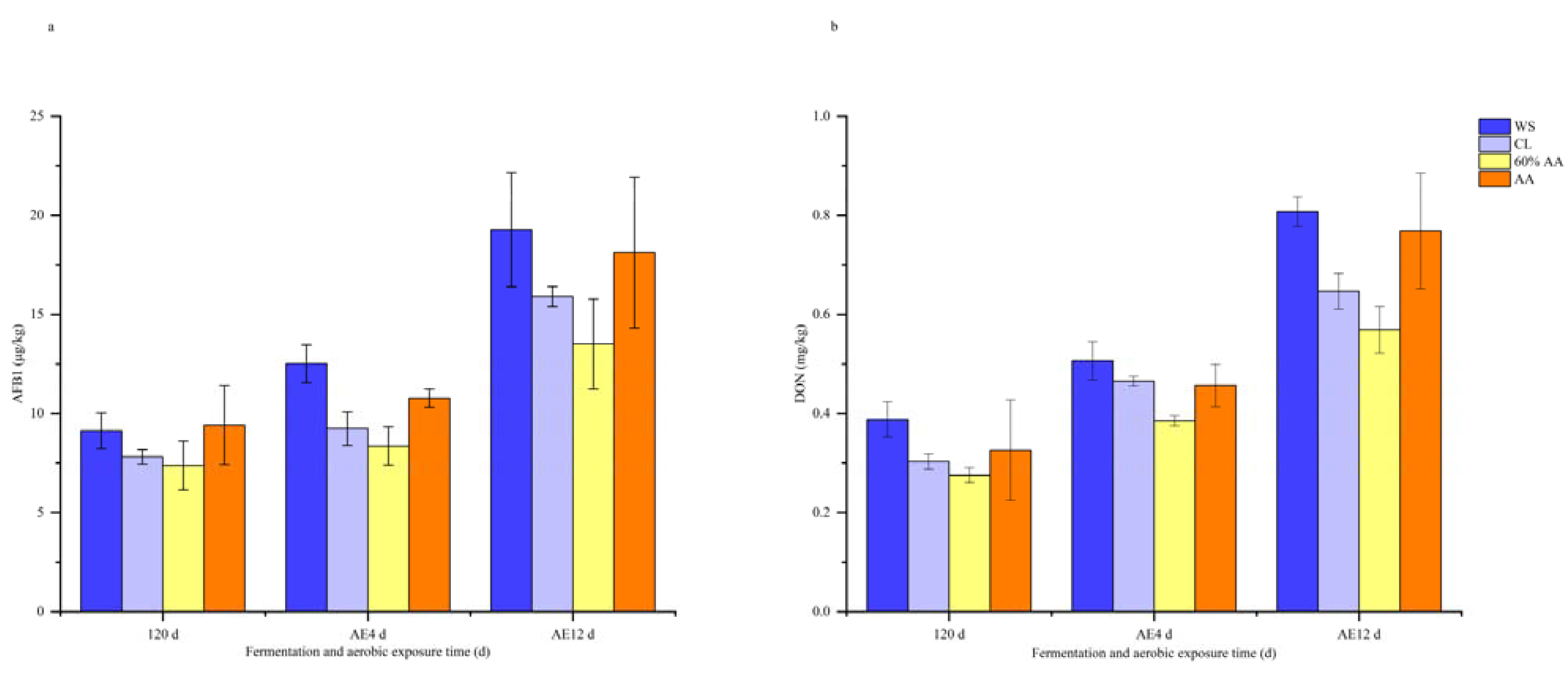
2.3. Mycotoxin Contents of Different Treatments during Aerobic Exposure
2.4. Microbial Community and Correlation Analysis
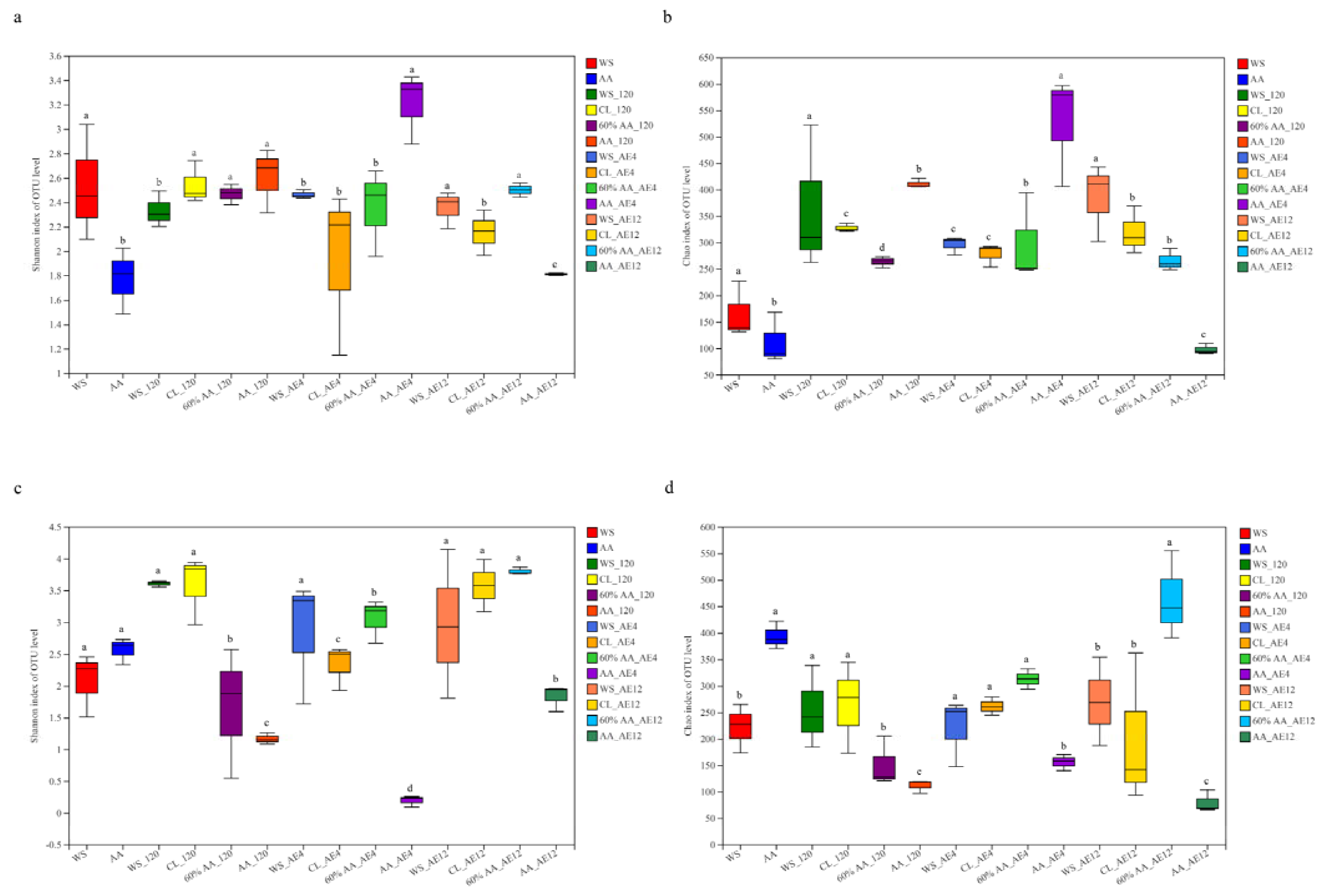
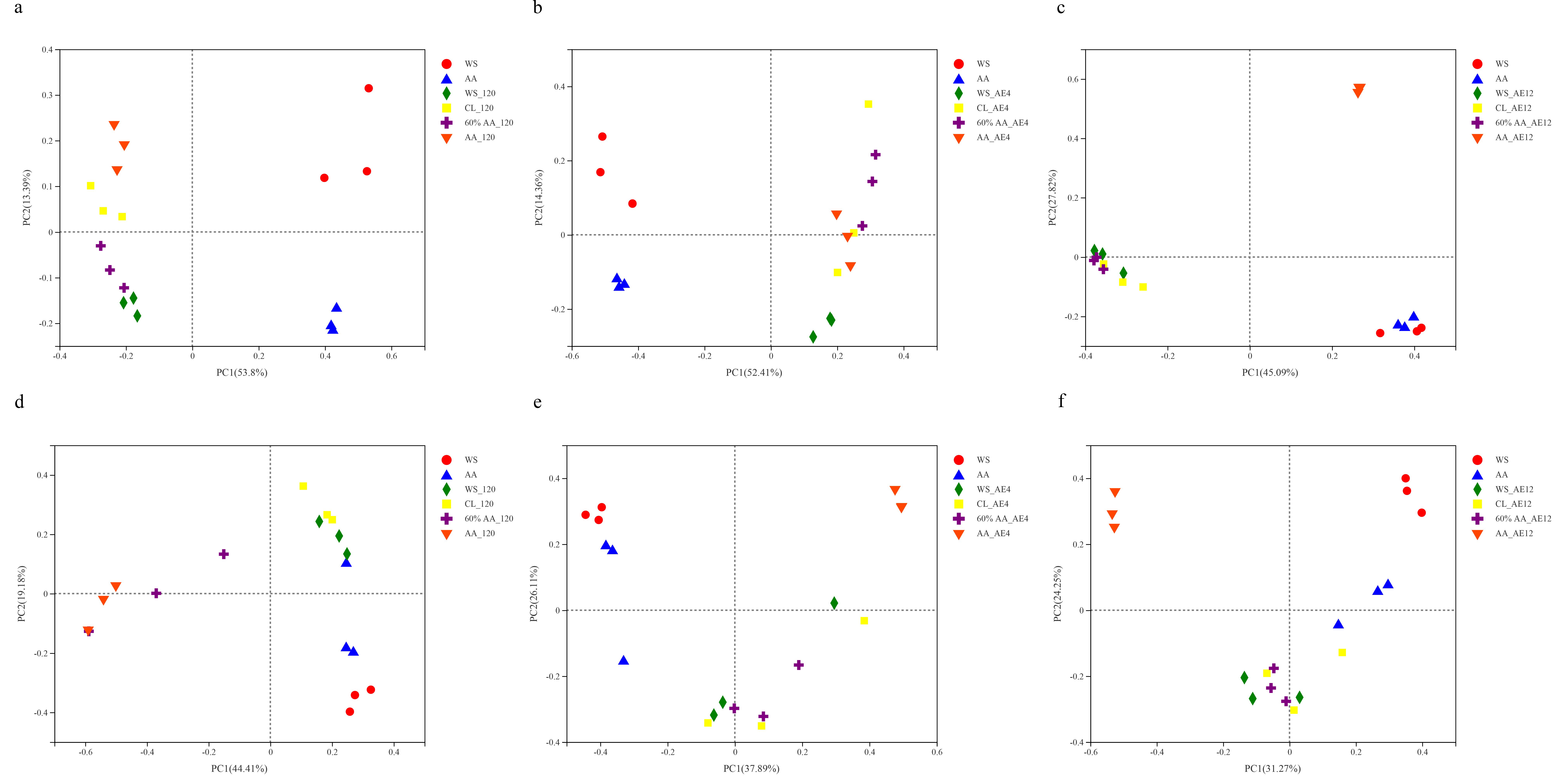
2.4.1. Microbial Species Diversity
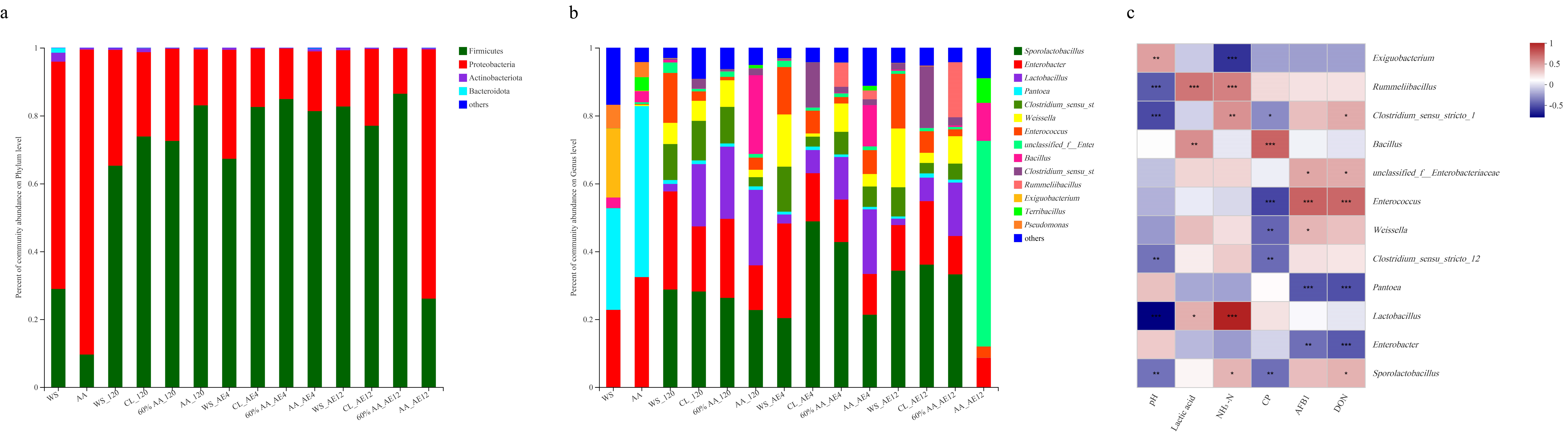
2.4.2. Bacterial Community and Correlation Analysis
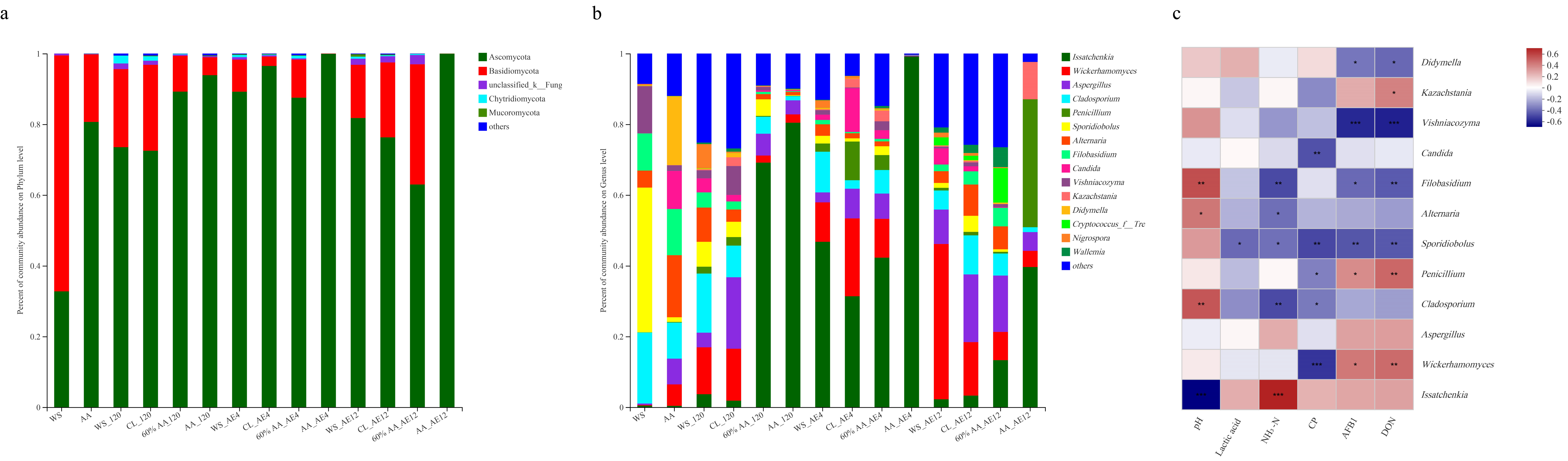
2.4.3. Fungal Community and Correlation Analysis
3. Conclusions
4. Materials and Methods
4.1. Raw Material Collection and Production
4.2. Fermentation Characteristic, Microbial Population, and Mycotoxin Content Analysis
4.3. Bacterial and Fungal Community Analysis
4.4. Calculation and Data Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Johns, L.; Bebber, D.; Gurr, S.; Brown, N. Emerging health threat and cost of Fusarium mycotoxins in European wheat. Nat. Food 2022, 3, 1014–1019. [Google Scholar] [CrossRef]
- Poole, N.; Donovan, J.; Erenstein, O. Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy 2020, 100, 101976. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F. Silage and the safety and quality of dairy foods: A review. Agric. Food Sci. 2013, 22, 16–34. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, Y.; Nennich, T.D.; Li, Y.; Liu, X. Transfer of dietary Aflatoxin B1 to milk Aflatoxin M1 and effect of inclusion of adsorbent in the diet of dairy cows. J. Dairy Sci. 2015, 98, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Wiedmeier, R.; Provenzat, F.; Burritt, E. Exposure to ammoniated wheat straw as suckling calves improves performance of mature beef cows wintered on ammoniated heat straw. J. Anim. Sci. 2002, 80, 2340–2348. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Sun, W.; Yan, Z.; Zhang, Y.; Wang, Z.; Song, B.; Li, J. Bioaugmentation of sweet sorghum ensiling with rumen fluid: Fermentation characteristics, chemical composition, microbial community, and enzymatic digestibility of silages. J. Clean. Prod. 2021, 294, 126308. [Google Scholar] [CrossRef]
- Adácsi, C.; Kovács, S.; Pócsi, I.; Győri, Z.; Dombrádi, Z.; Pusztahelyi, T. Microbiological and toxicological evaluation of fermented forages. Agriculture 2022, 12, 421. [Google Scholar] [CrossRef]
- Gallo, A.; Fancello, F.; Ghilardelli, F.; Zara, S.; Spanghero, M. Effects of several commercial or pure lactic acid bacteria inoculants on fermentation and mycotoxin levels in high-moisture corn silage. Anim. Feed. Sci. Technol. 2022, 286, 115256. [Google Scholar] [CrossRef]
- Ogunade, I.; Martinez-Tuppia, C.; Queiroz, O.C.M.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A.T. Silage review: Mycotoxins in silage: Occurrence, effects, prevention, and mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef]
- Prieto-Prieto, R.; Valencia, E.; Tirado-Corbala, R. Nitrogen levels and age at harvest and their effects on the nutritive value and fermentative characteristics of corn (Zea mays L.). J. Agr. Univ. Puerto Rico 2018, 101, 79–90. [Google Scholar] [CrossRef]
- Randby, A.; Bakken, A.K. Bunkers or round bales: Losses and silage quality with or without acid treatment of low dry matter grass crops. Anim. Feed. Sci. Technol. 2021, 275, 114868. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef] [PubMed]
- Muck, R. A Lactic Acid Bacteria Strain To Improve Aerobic Stability Of Silages. In Research Summaries; US Dairy Forage Research Center: Madison, WI, USA, 1996; pp. 42–43. [Google Scholar]
- Muck, R.; Nadeau, E.; McAllister, T.; Contreras-Govea, F.; Santos, M.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, W.; Gao, C.; Zhang, X.; Weng, B.; Cai, Y. Silage preparation and fermentation quality of kudzu, sugarcane top and their mixture treated with lactic acid bacteria, molasses and cellulase. J. Anim. Sci. 2017, 88, 1715–1721. [Google Scholar] [CrossRef]
- Molnar, J.; Szebeni, G.J.; Csupor-Loffler, B.; Hajdú, Z.; Szekeres, T.; Saiko, P.; Ocsovszki, I.; Puskás, L.; Hohmann, J.; Zupkó, I. Investigation of the Antiproliferative properties of natural sesquiterpenes from Artemisia asiatica and Onopordum acanthium on HL-60 cells in vitro. Int. J. Mol. Sci. 2016, 17, 83. [Google Scholar] [CrossRef]
- Lee, D.; Kim, C.E.; Park, S.Y.; Kim, K.; Hiep, N.T.; Lee, D.; Jang, H.; Lee, J.; Kang, K. Protective effect of Artemisia argyi and its flavonoid constituents against contrast-induced cytotoxicity by iodixanol in LLC-PK1 cells. Int. J. Mol. Sci. 2018, 19, 1387. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Variation of microbial community and fermentation quality in corn silage treated with lactic acid bacteria and Artemisia argyi during aerobic exposure. Toxins 2022, 14, 349. [Google Scholar] [CrossRef]
- Kim, S.; Jin, S.; Kang, S. Effects of feeding mugwort powder on meat composition and sensory characteristics in gilt. Korean J. Food Sci. Animal 2009, 29, 68–74. [Google Scholar] [CrossRef]
- Ko, Y.D.; Kim, J.H.; Adesogan, A.T.; Ha, M.H.; Kim, S.C. The effect of replacing rice straw with dry wormwood (Artemisia sp.) on intake, digestibility, nitrogen balance and ruminal fermentation characteristics in sheep. Anim. Feed. Sci. Technol. 2006, 125, 99–110. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Yang, J.; Qing, Y.; Xiao, J.; Mei, Y.; Lin, S. Effects of Artemisia argyi flavonoids on growth performance and immune function in broilers challenged with lipopolysaccharide. Animal Biosci. 2021, 34, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zuo, W.; Li, F. Dietary addition of Artemisia argyi reduces diarrhea and modulates the gut immune function without affecting growth performances of rabbits after weaning. J. Animal Sci. 2019, 97, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Jessie, G.; Vern, S.B.; Karen, A.B. Corn forage yield and quality for silage in short growing season areas of the Canadian prairies. Agronomy 2018, 8, 164. [Google Scholar]
- Smith, L.H. Theoretical carbohydrate requirement for alfalfa silage production. Agron. J. 1962, 54, 291–293. [Google Scholar] [CrossRef]
- Haigh, P.M.; Parker, J.W.G. Effect of silage additives and wilting on silage fermentation, digestibility and intake, and on liveweight change of young cattle. Grass Forage Sci. 1985, 40, 429–436. [Google Scholar] [CrossRef]
- Pitt, J.I.; Taniwaki, M.H.; Cole, M.B. Mycotoxin production in major crops as influenced by growing, harvesting, storage and processing, with emphasis on the achievement of food safety objectives. Food Control 2013, 32, 205–215. [Google Scholar] [CrossRef]
- Santos Pereira, C.; Cunha, S.C.; Fernandes, J.O. Prevalent mycotoxins in animal feed: Occurrence and analytical methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef]
- Kung, L.; Taylor, C.; Lynch, M.P.; Neylon, J.M. The effect of treating alfalfa with Lactobacillus buchneri 40788 on silage fermentation, aerobic stability, and nutritive value for lactating dairy cows. J. Dairy Sci. 2003, 86, 336–343. [Google Scholar] [CrossRef]
- Kung, L.M.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Robinson, P.; Swanepoel, N.; Heguy, J.; Price, T.; Meyer, D. “Shrink” losses in commercially sized corn silage piles: Quantifying total losses and where they occur. Sci. Total Environ. 2016, 542, 530–539. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Dynamics changes of microorganisms community and fermentation quality in soybean meal prepared with lactic acid bacteria and Artemisia argyi through fermentation and aerobic exposure processes. Foods 2022, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Pahlow, G.; Muck, R.; Driehuis, F.; Oude, S.; Spoelstra, S. Microbiology of ensiling. In Silage Science and Technology; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; American Society of Agronomy: Madison, WI, USA, 2003; pp. 31–93. [Google Scholar]
- Wang, X.; Song, J.; Liu, Z.; Zhang, G.; Zhang, Y. Fermentation quality and microbial community of corn stover or rice straw silage mixed with soybean curd residue. Animals 2022, 12, 919. [Google Scholar] [CrossRef] [PubMed]
- Mugabe, W.Z.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Effects of hexanoic acid, Lactobacillus plantarum and their combination on the fermentation characteristics of Napier grass. Animal Feed. Sci. Tech. 2019, 253, 135–140. [Google Scholar] [CrossRef]
- Wang, Y.; He, L.; Xing, Y.; Zheng, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Dynamics of bacterial community and fermentation quality during ensiling of wilted and unwilted Moringa oleifera leaf silage with or without lactic acid bacterial inoculants. mSphere 2019, 4, e00341-19. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Chen, N.; Lv, H.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Gallic acid influencing fermentation quality, nitrogen distribution and bacterial community of high-moisture mulberry leaves and stylo silage. Bioresour. Technol. 2020, 295, 122255. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M.; Keim, J.; Balocchi, O.; Lee, M. In vitro fermentation patterns and methane output of perennial ryegrass differing in water-soluble carbohydrate and nitrogen concentrations. Animals 2020, 10, 1076. [Google Scholar] [CrossRef] [PubMed]
- Wen, A.; Yuan, X.; Wang, J.; Desta, S.; Shao, T. Effects of four short-chain fatty acids or salts on dynamics of fermentation and microbial characteristics of alfalfa silage. Animal Feed. Sci. Technol. 2017, 223, 141–148. [Google Scholar] [CrossRef]
- Li, M.; Zi, X.; Zhou, H.; Lv, R.; Tang, J.; Cai, Y. Silage fermentation and ruminal degradation of cassava foliage prepared with microbial additive. AMB Express 2019, 9, 180. [Google Scholar] [CrossRef]
- Silva, E.B.; Smith, M.L.; Savage, R.M.; Polukis, S.A.; Drouin, P.; Kung, L. Effects of Lactobacillus hilgardii 4785 and Lactobacillus buchneri 40788 on the bacterial community, fermentation and aerobic stability of high-moisture corn silage. J. Appl. Microbiol. 2021, 130, 1481–1493. [Google Scholar] [CrossRef]
- Bao, W.; Mi, Z.; Xu, H.; Zheng, Y.; Kwok, L.; Zhang, H.; Zhang, W. Assessing quality of Medicago sativa silage by monitoring bacterial composition with single molecule, real-time sequencing technology and various physiological parameters. Sci. Rep. 2016, 6, 28358. [Google Scholar] [CrossRef]
- Cai, Y.; Kumai, S.; Ogawa, M.; Benno, Y.; Nakase, T. Characterization and identification of Pediococcus species isolated from forage crops and their application for silage preparation. Appl. Environ. Microbiol. 1999, 65, 2901–2906. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jiang, D.; Zheng, M.; Tian, P.; Zeng, M.; Xu, C. Microbial community dynamics during alfalfa silage with or without clostridial fermentation. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Guan, X.; Ge, D.; Li, S.; Huang, K.; Liu, J.; Li, F. Chemical composition and antimicrobial activities of Artemisia argyi Levl. et Vant essential oils extracted by simultaneous distillation-extraction, subcritical extraction and hydrodistillation. Molecules 2019, 24, 483. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.; Henderson, A.; Heron, S. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991; p. 340. [Google Scholar]
- Cogan, T.; Hawkey, R.; Higgie, E.; Lee, M.R.F.; Mee, E.; Parfitt, D.; Raj, J.; Roderick, S.; Walker, N.; Ward, P.; et al. Silage and total mixed ration hygienic quality on commercial farms: Implications for animal production. Grass Forage Sci. 2017, 72, 601–613. [Google Scholar] [CrossRef]
- González, J.J.; Ferreiroa, V.M.; Rodríguez, C.I.; Alfonso, A.; Sainz, M.J.; Aguín, O.; Vieytes, M.; Gomes, A.; Ramos, I.; Botana, L.M. Occurrence of mycotoxins and mycotoxigenic fungi in silage from the north of Portugal at feed-out. Int. J. Food Microbiol. 2022, 365, 109556. [Google Scholar] [CrossRef] [PubMed]
- Federica, C.; Campagnoli, A.; Orto, V.D. Fungal populations and mycotoxins in silages: From occurrence to analysis. Animal Feed. Sci. Technol. 2013, 183, 1–16. [Google Scholar]
- Antonio, G.; Francesco, F.; Francesca, G.; Severino, Z.; Federico, F.; Mauro, S. Effects of several lactic acid bacteria inoculants on fermentation and mycotoxins in corn silage. Animal Feed. Sci. Technol. 2021, 277, 114962. [Google Scholar]
- Adebo, O.A.; Kayitesi, E.; Njobeh, P.B. Reduction of mycotoxins during fermentation of whole grain sorghum to whole grain ting (a Southern African food). Toxins 2019, 11, 180. [Google Scholar] [CrossRef]
- Abiri, R.; Silva, A.L.M.; Mesquita, L.; Mesquita, J.; Atabaki, N.; Almeida, E., Jr.; Shaharuddin, N.; Malik, S. Towards a better understanding of Artemisia vulgaris: Botany, phytochemistry, pharmacological and biotechnological potential. Food Res. Int. 2018, 109, 403–415. [Google Scholar] [CrossRef]
- Lee, D.; Kim, K.; Lee, D.; Kang, K. Anti-apoptotic and antioxidant effects of 3-epi-iso-seco-tanapartholide isolated from Artemisia argyi against iodixanol-induced kidney epithelial cell death. Biomolecules 2020, 10, 867. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Yang, S.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; Jiang, Y.; Cervantes, A.A.; Kim, D.; Oliveria, A.; Vyas, D.; Weinberg, Z.; Jeong, K.; Adesogan, A. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157: H7 and silage additives. J. Dairy Sci. 2017, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Zhang, Q. Ensiling characteristics, proteolysis and bacterial community of high-moisture corn stalk and stylo silage prepared with Bauhinia variegate flower. Bioresour. Technol. 2019, 296, 122336. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Yang, X.; Wang, W.; Li, J.; Dong, Z.; Zhao, J.; Shao, T. The effects of natamycin and hexanoic acid on the bacterial community, mycotoxins concentrations, fermentation profiles, and aerobic stability of high moisture whole-crop corn silage. Animal Feed. Sci. Technol. 2022, 286, 115250. [Google Scholar] [CrossRef]
- Ma, H.; Wang, W.; Wang, Z.; Tan, Z.; Qin, G.; Wang, Y.; Pang, H. Microbial population succession and community diversity and its correlation with fermentation quality in soybean meal treated with Enterococcus faecalis during fermentation and aerobic exposure. Microorganisms 2022, 10, 530. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, M.Y.; Wu, S.; Zou, X.; Chen, X.; Ge, L.F.; Zhang, Q. Effects of gallic acid on fermentation parameters, protein fraction, and bacterial community of whole plant soybean silage. Front. Microbiol. 2021, 12, 962. [Google Scholar] [CrossRef]
- Walterson, A.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef]
- Walker, G.; Yustyniuk, V.; Shamoun, J.; Jacob, M.; Correa, M.; Vaden, S.; Borst Luke, B. Detection of Escherichia coli and Enterococcus spp. in dogs with polymicrobial urinary tract infections: A 5-year retrospective study. J. Vet. Intern. Med/ 2022, 36, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Kim, J.; Joo, M.; Shin, Y.J.; Lee, K.; Lee, C.; Kim, H.; Kim, D. Enterococcus faecium and Pediococcus acidilactici deteriorate Enterobacteriaceae-induced depression and colitis in mice. Sci. Rep/ 2022, 12, 9389. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Bai, C.; Xu, H.; Na, N.; Jiang, Y.; Yin, G.; Liu, S.; Xue, Y. Succession of bacterial community during the initial aerobic, intense fermentation and stable phases of whole-plant corn silages treated with lactic acid bacteria suspensions prepared from other silages. Front. Microbiol. 2021, 12, 655095. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculants. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yuan, X.; Dong, Z.; Li, J.; Shao, T. Isolating and evaluating lactic acid bacteria strains for effectiveness on silage quality at low temperatures on the Tibetan Plateau. Anim. Sci. J. 2017, 88, 1722–1729. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yan, Y.; Dong, D.; Bao, Y.; Luo, T.; Chen, Q.; Wang, J. Effect of Issatchenkia terricola WJL-G4 on deacidification characteristics and antioxidant activities of red raspberry wine processing. J. Fungi 2021, 8, 17. [Google Scholar] [CrossRef]
- Roncero, M.I.G.; Concepción, H.; Manuel, R.; Fe, I.; Marta, M.P.; Zaira, C.; Fernando, C.; Jesús, D.; Raquel, R.; Ana, L.; et al. Fusarium as a model for studying virulence in soilborne plant pathogens. Physiol. Mol. Plant 2003, 62, 87–98. [Google Scholar]
- Storm, I.M.L.D.; Kristensen, N.B.; Raun, B.M.L.; Smedsgaard, J.; Thrane, U. Dynamics in the microbiology of maize silage during whole-season storage. J. Appl. Microbiol. 2010, 109, 1017–1026. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.; Yue, Z.; Tan, P.; Sun, M.; Ji, L.; Bai, Y.; Ma, X. Wickerhamomyces anomalus relieves weaning diarrhea via improving gut microbiota and redox homeostasis using a piglet model. Food Funct. 2022, 13, 11223–11235. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Y.; Fu, G.; Chen, Y.; Wan, Y.; Deng, M.; Cai, W.; Li, M. Improvement of the flavor of major ethyl ester compounds during Chinese Te-flavor Baijiu brewing by Wickerhamomyces anomalus. Food Biosci. 2022, 50, 102022. [Google Scholar] [CrossRef]
- Cristina, R.; Monia, L.; Aurelio, S.; Giovanni, M.; Gabriella, C. Combined application of antagonistic Wickerhamomyces anomalus BS91 strain and Cynara cardunculus L. leaf extracts for the control of postharvest decay of citrus fruit. Food Microbiol. 2020, 92, 103583. [Google Scholar]
- Radhamany, D.; Rao, P.S.; Kotigadde, S. Bioregulatory role of Wickerhamomyces anomalus killer toxin against pathogenic members of the genus Candida. J. Microbiol. Infect. Dis. 2019, 9, 155–160. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Agricultural Chemists: Rockville, MD, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Lewis, B.A.; Robertson, J.B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.A. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1997, 28, 639–642. [Google Scholar] [CrossRef]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol. 2011, 34, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. The sustainable mitigation of ruminal methane and carbon dioxide emissions by co-ensiling corn stalk with Neolamarckia cadamba leaves for cleaner livestock production. J. Clean. Prod. 2021, 311, 127680. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | WS | AA | |
|---|---|---|---|
| Fermentation quality | pH | 6.69 | 6.76 |
| NH3-N, g/kg DM | 4.01 | 10.54 | |
| Lactic acid, g/kg DM | 1.76 | 2.95 | |
| Acetic acid, g/kg DM | 0.94 | 1.32 | |
| Chemical composition | DM, % | 94.07 | 34.19 |
| WSC, g/kg DM | 17.54 | 62.13 | |
| NDF, g/kg DM | 705.00 | 497.51 | |
| ADF, g/kg DM | 470.32 | 370.68 | |
| CP, g/kg DM | 32.24 | 130.64 | |
| Microbial population (log cfu/g FM) | LAB | 4.43 | 7.13 |
| Bacillus | 7.10 | 7.32 | |
| Aerobic bacteria | 7.63 | 9.10 | |
| Yeast | 1.45 | 3.36 | |
| AFB1, μg/kg | 2.96 | 4.14 | |
| DON, mg/kg | 0.08 | 0.11 | |
| Item | Treatment | Days of Fermentation (days) | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 120 | AE 2 | AE 4 | AE 8 | AE 12 | T | E | T × E | |||
| pH | WS | 4.91 aC | 5.04 aB | 5.14 aA | 5.32 cA | 5.00 cB | 0.02 | <0.05 | <0.05 | 0.08 |
| CE | 4.92 aC | 5.06 aA | 5.00 aB | 5.02 cB | 5.11 cA | |||||
| LB | 4.83 aA | 4.81 cA | 4.82 bA | 4.83 dA | 5.05 cA | |||||
| CL | 4.81 aB | 4.77 cB | 4.80 bB | 4.78 dB | 5.12 cA | |||||
| 20% AA | 4.89 aB | 4.87 bC | 4.88 cC | 5.12 cA | 5.12 cA | |||||
| 40% AA | 4.81 aC | 4.83 cB | 4.83 cB | 4.92 cA | 5.08 cA | |||||
| 60% AA | 4.57 aC | 4.88 bA | 4.78 cB | 4.77 dB | 4.85 dA | |||||
| 80% AA | 4.09 bC | 4.32 dB | 4.62 cB | 7.00 aA | 7.15 aA | |||||
| AA | 3.97 bB | 3.94 eB | 3.98 dB | 6.27 bA | 5.58 bA | |||||
| NH3-N (g/kg DM) | WS | 10.78 dA | 11.34 dA | 10.87 cA | 12.17 cA | 8.80 cB | 0.01 | <0.05 | <0.05 | <0.05 |
| CE | 10.96 dA | 13.47 dA | 12.92 cA | 11.52 cA | 10.15 cA | |||||
| LB | 17.03 cB | 18.33 cA | 19.01 aA | 16.36 bB | 15.02 aC | |||||
| CL | 18.27 bB | 18.50 cA | 17.12 bA | 16.86 bC | 16.51 aC | |||||
| 20% AA | 21.19 aA | 19.83 cC | 19.96 aC | 19.03 bC | 20.20 aB | |||||
| 40% AA | 26.01 aA | 23.92 bA | 22.97 aA | 21.65 aB | 21.36 aB | |||||
| 60% AA | 23.24 cA | 23.00 bA | 21.34 aA | 21.09 aA | 18.61 aB | |||||
| 80% AA | 25.51 aA | 24.24 cA | 23.13 bA | 22.18 cB | 20.54 cB | |||||
| AA | 26.42 aA | 25.72 aA | 24.47 aA | 23.65 bB | 21.29 bB | |||||
| Lactic acid (g/kg DM) | WS | 2.70 bB | 2.14 bB | 1.95 bB | 2.59 bA | 1.03 aA | 0.08 | 0.22 | 0.12 | 0.38 |
| CE | 2.20 bB | 3.05 bA | 3.27 bA | 3.29 aA | 1.67 aA | |||||
| LB | 1.73 bA | 1.35 bA | 1.35 cA | 1.58 bA | 1.35 bA | |||||
| CL | 1.35 bA | 1.35 bA | 1.35 cA | 1.35 bA | 1.47 bA | |||||
| 20% AA | 1.35 bA | 1.35 bA | 1.35 cA | 1.35 bA | 1.35 bA | |||||
| 40% AA | 2.35 bA | 1.35 bA | 1.35 cA | 1.35 bA | 1.35 bA | |||||
| 60% AA | 3.79 bA | 1.91 bB | 5.79 aA | 3.92 aA | 2.98 aA | |||||
| 80% AA | 9.15 aA | 6.99 aB | 4.02 aB | 1.35 bC | 1.66 bC | |||||
| AA | 11.78 aA | 11.68 aA | 5.31 aB | 1.35 bC | 2.60 aC | |||||
| Acetic acid (g/kg DM) | WS | 2.17 bA | 2.58 bA | 2.25 bA | 2.56 cA | 2.90 bA | 0.11 | 0.32 | 0.18 | 0.55 |
| CE | 2.74 bA | 2.73 bA | 2.33 bA | 2.70 cA | 2.79 bA | |||||
| LB | 5.95 aB | 4.62 aC | 7.01 aA | 6.04 aB | 5.73 aB | |||||
| CL | 4.67 aA | 4.63 aA | 3.56 bA | 2.64 cB | 5.66 aA | |||||
| 20% AA | 4.11 aA | 3.43 bB | 4.16 bA | 4.13 bA | 4.99 aA | |||||
| 40% AA | 3.89 aA | 3.23 bB | 4.40 aA | 2.67 cB | 3.95 aA | |||||
| 60% AA | 2.63 bA | 4.41 aA | 3.02 bA | 3.47 bA | 2.38 bB | |||||
| 80% AA | 3.97 aA | 3.22 bA | 1.50 bB | 1.29 dB | 1.29 bB | |||||
| AA | 2.16 bA | 1.90 cA | 1.38 bA | 1.29 dA | 1.29 bA | |||||
| Item | Treatment | Days of Fermentation (d) | SEM | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 120 | AE 2 | AE 4 | AE 8 | AE 12 | T | E | T × E | |||
| DM (%) | WS | 48.74 aB | 51.21 aA | 49.95 aB | 59.45 bA | 72.40 aA | 3.45 | 1.41 | 0.81 | 2.44 |
| CE | 44.14 aC | 48.01 aC | 50.24 aB | 60.36 aA | 69.71 aA | |||||
| LB | 37.18 aB | 36.90 bB | 40.46 bB | 46.77 cA | 57.23 bA | |||||
| CL | 33.19 bB | 34.73 bB | 38.54 bB | 39.15 dB | 53.67 bA | |||||
| 20% AA | 33.38 aB | 37.15 bB | 41.02 aA | 49.19 cA | 48.60 bA | |||||
| 40% AA | 30.45 bB | 30.46 cB | 32.83 bA | 36.93 dA | 39.48 cA | |||||
| 60% AA | 27.19 bB | 28.93 cB | 31.74 bB | 32.51 dB | 44.65 cA | |||||
| 80% AA | 26.56 bB | 26.32 cB | 27.27 cB | 32.20 dB | 40.53 cA | |||||
| AA | 24.62 bB | 25.14 cB | 25.85 cB | 31.07 dA | 36.52 cA | |||||
| WSC (g/kg DM) | WS | 6.29 dA | 5.86 cA | 5.03 cA | 3.15 cB | 2.18 cB | 0.11 | < 0.05 | < 0.05 | 0.08 |
| CE | 6.65 dA | 6.47 cA | 5.19 cA | 4.82 cB | 3.17 cB | |||||
| LB | 4.88 eA | 5.62 cA | 4.70 dA | 4.12 cA | 2.76 cB | |||||
| CL | 7.11 dA | 5.80 cA | 5.44 cA | 4.56 cB | 2.97 cB | |||||
| 20% AA | 6.45 dA | 5.62 cA | 5.06 cA | 4.41 cA | 3.96 cA | |||||
| 40% AA | 12.78 dA | 6.96 cA | 7.70 cA | 6.66 cA | 5.98 cA | |||||
| 60% AA | 22.34 cA | 11.71 bA | 12.90 cA | 10.36 bB | 9.62 bB | |||||
| 80% AA | 30.22 bA | 19.96 bB | 17.75 bB | 14.75 bC | 11.56 bC | |||||
| AA | 43.22 aA | 40.97 aA | 35.67 aB | 33.98 aB | 21.72 aC | |||||
| NDF (g/kg DM) | WS | 664.27 aA | 662.37 aA | 660.67 aA | 657.98 aA | 656.96 aA | 4.36 | 1.77 | 1.02 | 3.08 |
| CE | 638.27 bA | 635.66 bA | 630.38 bA | 626.30 bB | 624.80 bB | |||||
| LB | 661.87 aA | 660.95 aA | 659.22 aB | 657.17 aB | 655.37 aB | |||||
| CL | 648.16 bA | 646.58 bA | 645.28 bA | 636.94 aB | 633.42 bB | |||||
| 20% AA | 628.00 cA | 623.19 bA | 622.42 cA | 621.86 bA | 619.07 bA | |||||
| 40% AA | 585.24 dA | 582.38 cA | 580.20 dA | 577.68 cA | 576.34 cA | |||||
| 60% AA | 532.53 eA | 530.11 cA | 526.39 eB | 524.93 cB | 521.22 dB | |||||
| 80% AA | 516.25 eA | 513.05 cA | 512.50 eA | 511.67 cA | 509.70 dA | |||||
| AA | 453.08 fA | 449.18 dA | 448.40 fA | 446.19 dA | 445.64 eA | |||||
| ADF (g/kg DM) | WS | 438.46 aA | 432.26 aA | 430.46 aA | 426.71 aB | 425.39 aB | 3.87 | 1.57 | 0.91 | 2.73 |
| CE | 411.71 bA | 400.05 bA | 397.52 bB | 395.61 bB | 392.31 bB | |||||
| LB | 434.75 aA | 427.87 bA | 423.74 aA | 420.05 aA | 418.83 aA | |||||
| CL | 418.25 bA | 412.97 bA | 409.39 bB | 405.48 aB | 403.98 bB | |||||
| 20% AA | 434.29 aA | 432.04 aA | 431.18 aA | 429.44 aA | 427.85 aA | |||||
| 40% AA | 410.66 bA | 409.01 bA | 407.95 bA | 406.66 aA | 405.97 bA | |||||
| 60% AA | 376.36 cA | 373.68 cA | 370.86 cA | 368.88 cB | 367.82 cB | |||||
| 80% AA | 363.57 cA | 361.18 cA | 360.29 cA | 359.11 cA | 358.66 cA | |||||
| AA | 350.25 dA | 347.23 cA | 346.26 dA | 345.28 cA | 344.49 dA | |||||
| CP (g/kg DM) | WS | 23.93 eA | 23.07 dA | 21.50 eA | 20.93 dB | 19.01 dB | 0.26 | 0.11 | 0.06 | 0.18 |
| CE | 24.87 eA | 23.22 dA | 22.27 eB | 21.37 dB | 20.10 dB | |||||
| LB | 20.40 eA | 18.87 eA | 21.70 eA | 19.73 dA | 18.87 dA | |||||
| CL | 26.93 dA | 23.73 dA | 23.63 eA | 22.93 dA | 21.87 dA | |||||
| 20% AA | 31.07 dA | 31.11 dA | 30.30 dA | 30.03 cA | 29.73 cA | |||||
| 40% AA | 35.67 dA | 34.63 cA | 33.47 dA | 32.37 cB | 30.67 cB | |||||
| 60% AA | 49.03 cA | 48.00 cA | 47.87 cA | 47.53 bA | 48.07 bA | |||||
| 80% AA | 67.83 bA | 68.87 bA | 62.97 bA | 61.13 aB | 56.97 bB | |||||
| AA | 81.00 aA | 76.87 aA | 75.70 aA | 74.33 aA | 72.53 aB | |||||
| Item | Treatment | Days of Fermnetation (d) | SEM | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 120 | AE 2 | AE 4 | AE 8 | AE 12 | T | E | T × E | |||
| Lactic acid bacteria | WS | 6.60 aA | 5.47 cB | 7.05 aA | 5.36 dB | 7.12 bA | 0.27 | 0.10 | 0.06 | 0.18 |
| CE | 7.04 aA | 7.11 aA | 7.28 aA | 6.94 cB | 7.00 bA | |||||
| LB | 6.61 aA | 7.03 aA | 5.50 cB | 6.79 cB | 6.91 bA | |||||
| CL | 7.11 aA | 6.80 bA | 6.75 aA | 6.57 cB | 6.70 cA | |||||
| 20% AA | 7.19 aA | 7.41 aA | 6.57 bA | 5.13 dB | 6.87 bA | |||||
| 40% AA | 7.24 aA | 7.04 aA | 6.61 bA | 6.47 cA | 6.67 cA | |||||
| 60% AA | 7.27 aA | 7.56 aA | 7.30 aA | 9.16 aA | 7.64 bA | |||||
| 80% AA | 7.16 aB | 6.92 bB | 7.41 aB | 8.00 bA | 8.76 aA | |||||
| AA | 5.19 bC | 6.60 bC | 7.50 aB | 7.06 cB | 8.97 aA | |||||
| Bacillus | WS | 4.50 bB | 5.38 cA | 5.90 cA | 5.07 bB | 4.94 bB | 0.28 | 0.11 | 0.07 | 0.20 |
| CE | 5.22 aB | 5.39 cA | 6.08 bA | 5.27 bB | 5.32 bB | |||||
| LB | 5.12 aA | 5.30 cA | 5.31 cA | 5.19 bA | 5.11 bA | |||||
| CL | 4.80 aA | 4.70 dA | 5.65 cA | 4.91 cA | 4.55 bA | |||||
| 20% AA | 5.27 aB | 5.32 cA | 5.47 cA | 5.28 bB | 5.03 bB | |||||
| 40% AA | 5.20 aA | 5.34 cA | 5.23 cA | 5.55 bA | 6.83 bA | |||||
| 60% AA | 5.08 aB | 5.91 aA | 5.68 cB | 5.81 bA | 5.26 bA | |||||
| 80% AA | 5.18 aB | 6.85 aB | 8.65 aA | 9.48 aA | 9.49 aA | |||||
| AA | 5.27 aC | 5.86 bC | 6.58 bB | 7.90 aB | 9.32 aA | |||||
| Aerobic bacteria | WS | 5.61 bC | 5.46 bC | 8.75 aA | 6.20 cB | 6.97 bB | 0.17 | 0.07 | <0.05 | 0.12 |
| CE | 5.50 bB | 6.71 aA | 6.81 cA | 6.77 cA | 7.14 bA | |||||
| LB | 5.34 bB | 3.80 dC | 6.48 cA | 6.85 cA | 6.79 bA | |||||
| CL | 5.35 bA | 4.78 cA | 5.38 cA | 5.64 cA | 5.31 cA | |||||
| 20% AA | 6.60 aB | 5.70 bB | 6.67 cA | 6.70 cA | 7.05 bA | |||||
| 40% AA | 5.51 bB | 5.49 bB | 6.60 cA | 7.20 bA | 6.85 bA | |||||
| 60% AA | 5.62 bB | 4.39 cC | 6.91 cA | 6.94 cA | 5.23 cB | |||||
| 80% AA | 5.37 bC | 7.36 aB | 7.59 bB | 9.71 aA | 9.62 aA | |||||
| AA | 6.44 aC | 5.22 bC | 7.32 bB | 9.35 aA | 9.77 aA | |||||
| Yeast | WS | 2.34 aC | 3.70 aC | 4.51 aB | 5.23 aB | 8.23 aA | 0.08 | <0.05 | <0.05 | <0.05 |
| CE | 1.23 aB | 1.23 bB | 1.49 bB | 2.97 cB | 5.23 cA | |||||
| LB | 1.39 aC | 2.09 bB | 1.38 bC | 2.39 cB | 3.96 dA | |||||
| CL | 1.23 aB | 1.23 bB | 2.32 bB | 1.23 dB | 5.13 cA | |||||
| 20% AA | ND | 2.88 aA | 3.71 aA | 3.60 bA | 4.89 cA | |||||
| 40% AA | ND | 1.03 bB | 1.39 bB | 2.03 cB | 4.68 cA | |||||
| 60% AA | ND | ND | 1.23 cB | 1.23 dB | 3.86 dA | |||||
| 80% AA | ND | 4.58 aB | 4.21 aB | 4.21 bB | 6.23 bA | |||||
| AA | ND | 3.42 aB | 3.66 aB | 4.36 bB | 7.51 bA | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Tan, Z.; Wu, G.; Wang, L.; Qin, G.; Wang, Y.; Pang, H. Investigation on Fermentation Characteristics and Microbial Communities of Wheat Straw Silage with Different Proportion Artemisia argyi. Toxins 2023, 15, 330. https://doi.org/10.3390/toxins15050330
Wang Z, Tan Z, Wu G, Wang L, Qin G, Wang Y, Pang H. Investigation on Fermentation Characteristics and Microbial Communities of Wheat Straw Silage with Different Proportion Artemisia argyi. Toxins. 2023; 15(5):330. https://doi.org/10.3390/toxins15050330
Chicago/Turabian StyleWang, Zhenyu, Zhongfang Tan, Guofang Wu, Lei Wang, Guangyong Qin, Yanping Wang, and Huili Pang. 2023. "Investigation on Fermentation Characteristics and Microbial Communities of Wheat Straw Silage with Different Proportion Artemisia argyi" Toxins 15, no. 5: 330. https://doi.org/10.3390/toxins15050330