Analysis of the Effect of Plutella xylostella Polycalin and ABCC2 Transporter on Cry1Ac Susceptibility by CRISPR/Cas9-Mediated Knockout

 and
and
Abstract
:1. Introduction
2. Results
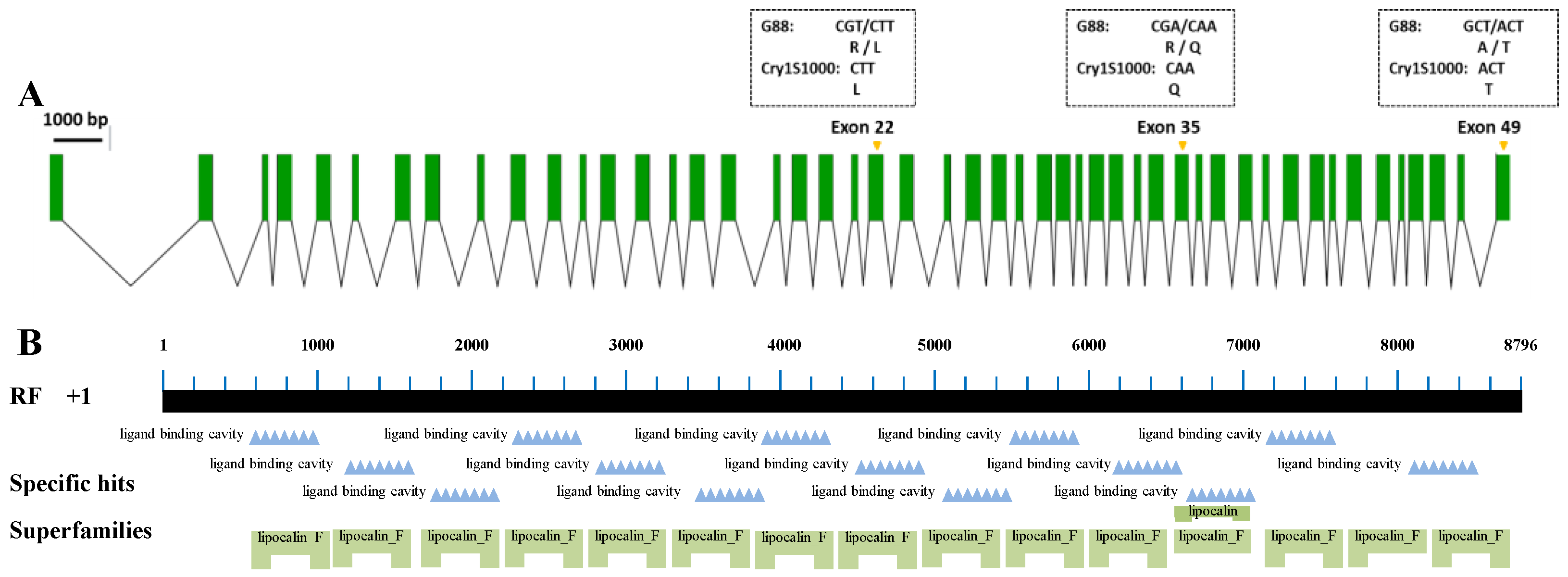
2.1. Pxpolycalin Sequence Comparison between P. xylostella Strains
2.2. Phylogenetic Analysis of Pxpolycalin in P. xylostella
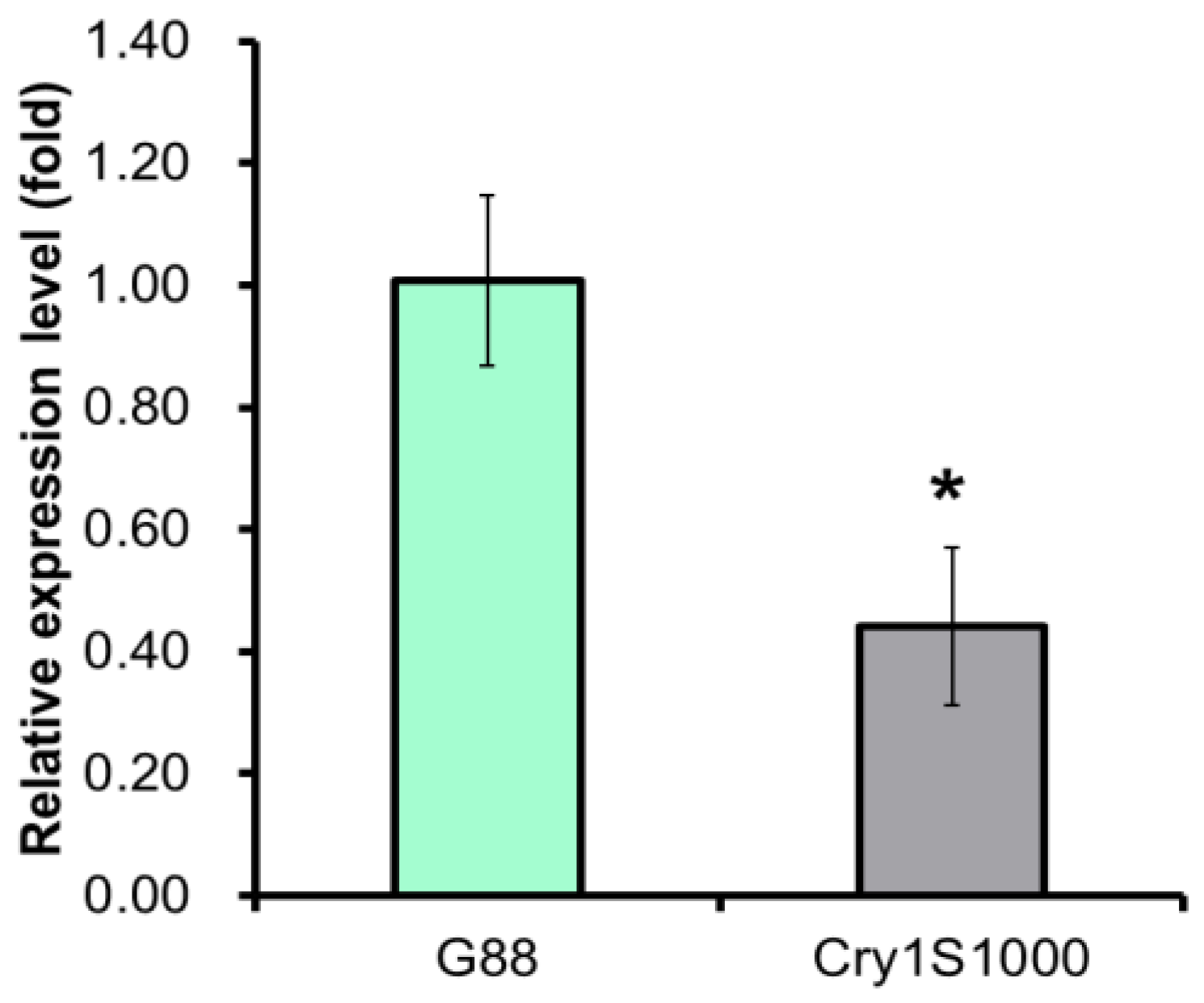
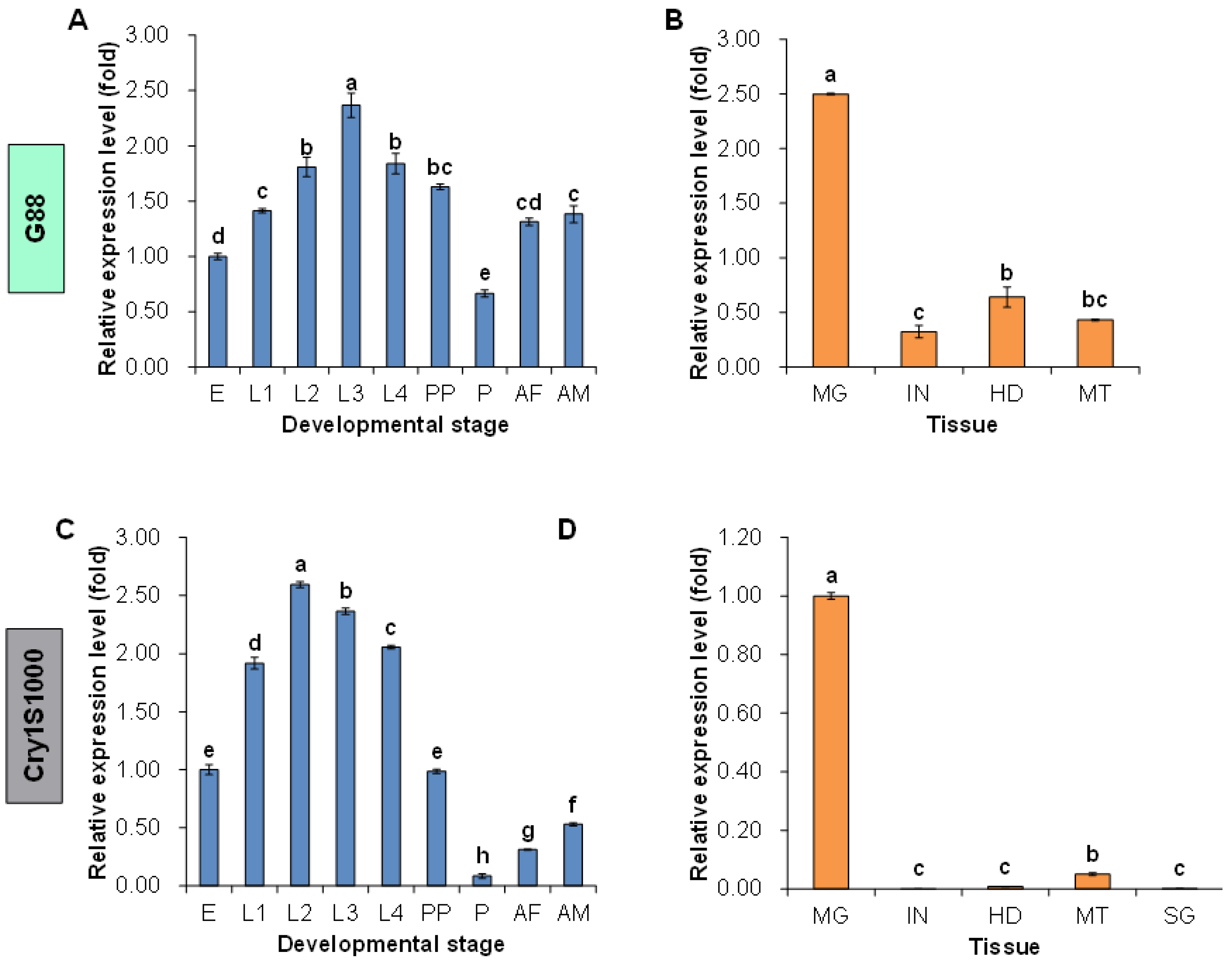
2.3. Spatio-Temporal Expression Patterns of the Pxpolycalin Gene in G88 and Cry1S1000 Strains
2.4. Induction of PxABCC2 and Pxpolycalin by Cry1Ac Toxin in G88 Strain
2.5. Genetic Linkage Analysis of the Pxpolycalin and PxABCC2 Genes
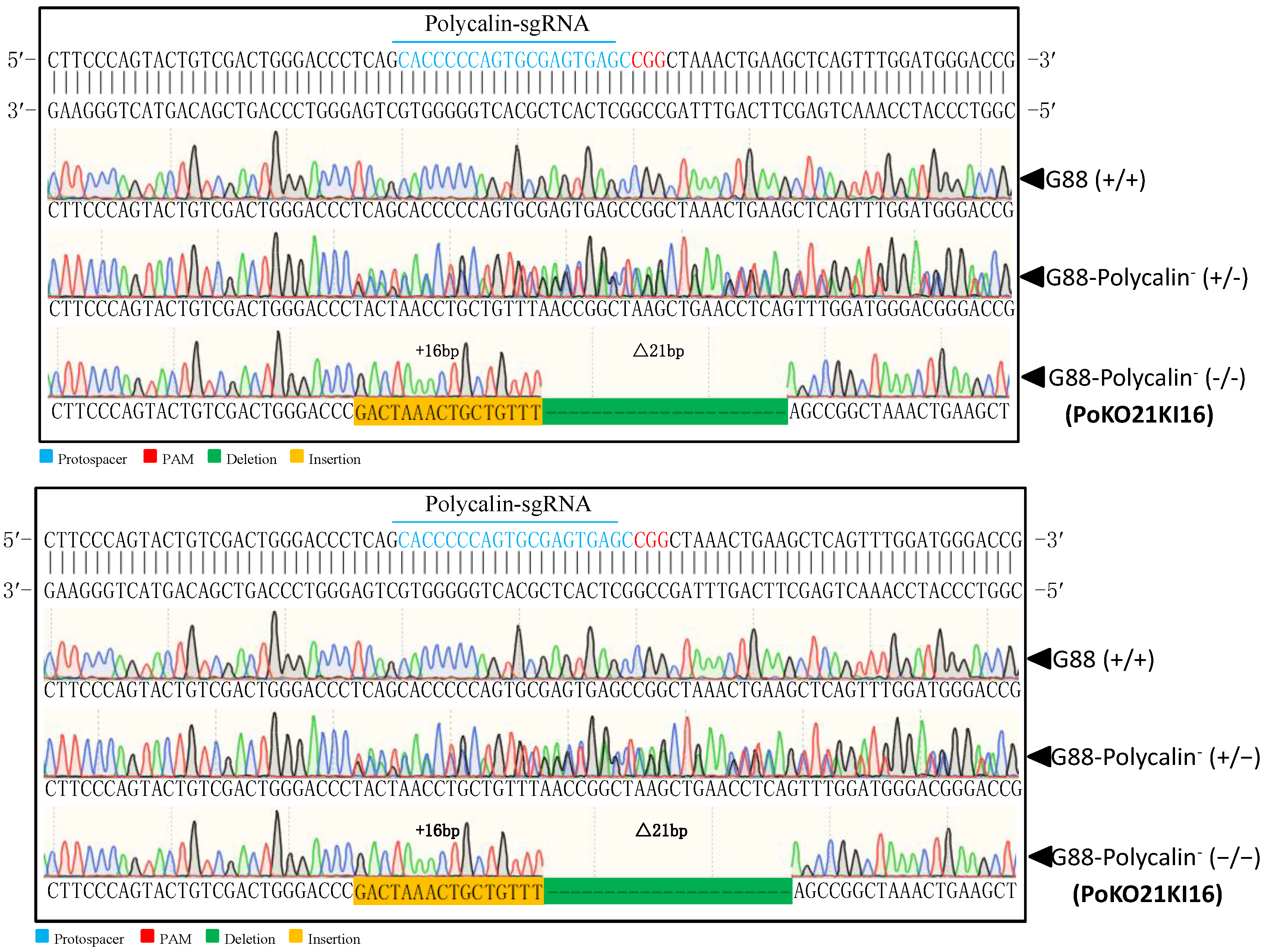
2.6. Mutagenesis of Pxpolycalin and PxABCC2 Mediated by CRISPR/Cas9
2.7. Effect of Pxpolycalin and PxABCC2 Mutations on the Susceptibility of Cry1Ac Toxin in P. xylostella
3. Discussion
4. Materials and Methods
4.1. Insect Strain and Rearing
4.2. Bt toxins and Bioassays
4.3. Identification of Sequence Differences between G88 and Cry1S1000 Strains
4.4. Construction of Phylogenetic Tree
4.5. Spatio-Temporal Expression Patterns of polycalin in P. xylostella
4.6. Cry1Ac Treatment
4.7. Genetic Linkage Analysis
4.8. Preparation of sgRNA and Cas9 Protein
4.9. Microinjection of P. xylostella Embryos and Screening of Homozygous Strains
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Usta, C. Microorganisms in Biological Pest Control—A Review (Bacterial Toxin Application and Effect of Environmental Factors); Silva-Opps, M., Ed.; InTech: Rijeka, Croatia, 1976; pp. 515–518. [Google Scholar]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Carriere, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberon, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabashnik, B.E.; Cushing, N.L.; Finson, N.; Johnson, M.W. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 1990, 83, 1671–1676. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.L.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of action, resistance, and new applications: A review. Crit. Rev. Biotechnol. 2016, 36, 317–326. [Google Scholar] [CrossRef]
- Zhang, X.; Candas, M.; Griko, N.B.; Rose-Young, L.; Bulla, L.A. Cytotoxicity of Bacillus thuringiensis Cry1Ab toxin depends on specific binding of the toxin to the cadherin receptor BT-R1 expressed in insect cells. Cell Death Differ. 2005, 12, 1407–1416. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Kang, S.; Chen, D.F.; Wu, Q.J.; Wang, S.; Xie, W.; Zhu, X.G.; Baxter, S.W.; Zhou, X.; Jurat-Fuentes, J.L.; et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth. PLoS Genet. 2015, 11, e1005124. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Kang, S.; Sun, D.; Gong, L.J.; Zhou, J.L.; Qin, J.Y.; Guo, L.; Zhu, L.H.; Bai, Y.; Ye, F.; et al. MAPK-dependent hormonal signaling plasticity contributes to overcoming Bacillus thuringiensis toxin action in an insect host. Nat. Commun. 2020, 11, 3003. [Google Scholar] [CrossRef]
- Guo, L.; Cheng, Z.Q.; Qin, J.Y.; Sun, D.; Wang, S.L.; Wu, Q.J.; Crickmore, N.; Zhou, X.G.; Bravo, A.; Soberon, M.; et al. MAPK-mediated transcription factor GATAd contributes to Cry1Ac resistance in diamondback moth by reducing PxmALP expression. PLoS Genet. 2022, 18, e1010037. [Google Scholar] [CrossRef]
- Sun, D.; Zhu, L.; Guo, L.; Wang, S.; Wu, Q.; Crickmore, N.; Zhou, X.; Bravo, A.; Soberon, M.; Guo, Z.; et al. A versatile contribution of both aminopeptidases N and ABC transporters to Bt Cry1Ac toxicity in the diamondback moth. BMC Biol. 2022, 20, 33. [Google Scholar] [CrossRef]
- Jha, S.; Rollins, M.G.; Fuchs, G.; Procter, D.J.; Hall, E.A.; Cozzolino, K.; Sarnow, P.; Savas, J.N.; Walsh, D. Trans-kingdom mimicry underlies ribosome customization by a poxvirus kinase. Nature 2017, 546, 651–655. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, K.; Yaoi, K.; Nagino, Y.; Hara, H.; Kitami, M.; Atsumi, S.; Miura, N.; Sato, R. Aminopeptidase N isoforms from the midgut of Bombyx mori and Plutella xylostella—Their classification and the factors that determine their binding specificity to Bacillus thuringiensis Cry1A toxin. FEBS Lett. 2002, 519, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Zhang, M.; Liang, G.; Wu, K.; Guo, Y.; Ni, X.; Li, X. APN1 is a functional receptor of Cry1Ac but not Cry2Ab in Helicoverpa zea. Sci. Rep. 2016, 6, 19179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Fan, J.; Zhang, B.; Liu, L.; Wang, X.; Lei, C.; Lin, Y.; Ma, W. RNA interference knockdown of aminopeptidase N genes decrease the susceptibility of Chilo suppressalis larvae to Cry1Ab/Cry1Ac and Cry1Ca-expressing transgenic rice. J. Invertebr. Pathol. 2017, 145, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, X.; He, H.; Chen, J.; Liu, Y.; Huang, W.; Ou, L.; Yang, Z.; Guan, X.; Zhang, L.; et al. Knockout of two Cry-binding aminopeptidase N isoforms does not change susceptibility of Aedes aegypti larvae to Bacillus thuringiensis subsp. israelensis Cry4Ba and Cry11Aa toxins. Insects 2021, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, P.; Jin, H.; Liu, H.; Zhou, H.; Qiu, L.; Lin, Y.; Ma, W. Knockdown of the aminopeptidase N genes decreases susceptibility of Chilo suppressalis larvae to Cry1Ab/Cry1Ac and Cry1Ca. Pestic. Biochem. Physiol. 2020, 162, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Atsumi, S.; Yaoi, K.; Nakanishi, K.; Higurashi, S.; Miura, N.; Tabunoki, H.; Sato, R. A cadherin-like protein functions as a receptor for Bacillus thuringiensis Cry1Aa and Cry1Ac toxins on midgut epithelial cells of Bombyx mori larvae. FEBS Lett. 2003, 538, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Morin, S.; Biggs, R.W.; Sisterson, M.S.; Shriver, L.; Ellers-Kirk, C.; Higginson, D.; Holley, D.; Gahan, L.J.; Heckel, D.G.; Carrière, Y.; et al. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc. Natl. Acad. Sci. USA 2003, 100, 5004–5009. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Xu, X.; Ye, W.; Yu, Z.; Sun, M. Helicoverpa armigera cadherin fragment enhances Cry1Ac insecticidal activity by facilitating toxin-oligomer formation. Appl. Microbiol. Biotechnol. 2010, 85, 1033–1040. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.; Guo, X.; Wan, P.; Liu, K.; Cong, S.; Wang, J.; Xu, D.; Xiao, Y.; Li, X.; et al. Pink bollworm resistance to Bt toxin Cry1Ac associated with an insertion in aadherin exon 20. Toxins 2019, 11, 186. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Zhang, X.; Zhong, J.; Liu, Y.; Zhang, C.; Xie, Y.; Lin, M.; Xu, C.; Lu, L.; Zhu, Q.; et al. Expression of Cry1Ac toxin-binding region in Plutella xyllostella cadherin-like receptor and studying their interaction mode by molecular docking and site-directed mutagenesis. Int. J. Biol. Macromol. 2018, 111, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Hu, X.; Zhang, X.; Zhong, J.; Lu, L.; Liu, Y.; Dong, S.; Wang, Y.; Liu, X. Identification of a Cry1Fa binding site of cadherin in Plutella xylostella through fragment exchanging and molecular docking methods. Int. J. Biol. Macromol. 2019, 146, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Gao, M.; Hu, X.; Lu, L.; Zhang, X.; Liu, Y.; Zhong, J.; Liu, X. Synergistic selection of a Helicoverpa armigera cadherin fragment with Cry1Ac in different cells and insects. Int. J. Biol. Macromol. 2020, 164, 3667–3675. [Google Scholar] [CrossRef] [PubMed]
- Jurat-Fuentes, J.L.; Adang, M.J. Characterization of a Cry1Ac-receptor alkaline phosphatase in susceptible and resistant Heliothis virescens larvae. Eur. J. Biochem. 2004, 271, 3127–3135. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, M.; Liang, G.; Li, X. Alkaline phosphatase 2 is a functional receptor of Cry1Ac but not Cry2Ab in Helicoverpa zea. Insect Biochem. Mol. Biol. 2019, 28, 372–379. [Google Scholar] [CrossRef]
- Gahan, L.J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin. PLoS Genet. 2010, 6, e1001248. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.T.; Zhang, T.; Liu, C.; Heckel, D.G.; Li, X.; Tabashnik, B.E.; Wu, K.M. Mis-splicing of the ABCC2 gene linked with Bt toxin resistance in Helicoverpa armigera. Sci. Rep. 2014, 4, 6184. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.J.; Kang, S.; Zhu, X.; Xia, J.X.; Wu, Q.; Wang, S.L.; Xie, W.; Zhang, Y.J. The novel ABC transporter ABCH1 is a potential target for RNAi-based insect pest control and resistance management. Sci. Rep. 2015, 5, 13728–13742. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.X.; Fu, S.; Ma, X.L.; Baxter, S.W.; Vasseur, L.; Xiong, L.; Huang, Y.P.; Yang, G.; You, S.J.; You, M.S. Resistance to Bacillus thuringiensis Cry1Ac toxin requires mutations in two Plutella xylostella ATP-binding cassette transporter paralogs. PLoS Pathog. 2020, 16, e1008697. [Google Scholar] [CrossRef]
- Xu, H.Q.; Ma, M.; Ma, Y.P.; Zhang, S.Y.; Li, W.J.; Wei, D.; Wang, J.J. Identification and expression characterization of ATP-binding cassette (ABC) transporter genes in melon fly. Insects 2021, 12, 270. [Google Scholar] [CrossRef]
- Yao, X.; Liu, C.; Duan, Y.; An, S.; Wei, J.; Liang, G. ABCC2 is a functional receptor of Bacillus thuringiensis Cry1Ca in Spodoptera litura. Int. J. Biol. Macromol. 2021, 194, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mao, K.; Jin, R.; Cai, T.; Qin, Y.; Zhang, Y.; He, S.; Ma, K.; Wan, H.; Ren, X.; et al. miRNA novel 268 targeting NlABCG3 is involved in nitenpyram and clothianidin resistance in Nilaparvata lugens. Int. J. Biol. Macromol. 2022, 217, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Shwe, S.M.; Prabu, S.; Jing, D.; He, K.; Wang, Z. Synergistic interaction of Cry1Ah and Vip3Aa19 proteins combination with midgut ATP-binding cassette subfamily C receptors of Conogethes punctiferalis (Guenee) (Lepidoptera: Crambidae). Int. J. Biol. Macromol. 2022, 213, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, S.Y.; Xu, X.J.; Lin, G.F.; Lin, S.J.; Bai, J.L.; Song, Q.S.; You, M.S.; Xie, M. Novel-miR-310 mediated response mechanism to Cry1Ac protoxin in Plutella xylostella (L.). Int. J. Biol. Macromol. 2022, 219, 587–596. [Google Scholar] [CrossRef]
- Zhu, Q.; Hu, X.; Liu, Y.; Xie, Y.; Xu, C.; Lin, M.; Pooe, O.J.; Zhong, J.; Gao, M.; Lu, L.; et al. Identification of single domain antibodies with insect cytotoxicity using phage-display antibody library screening and Plutella xylostella ATP-binding cassette transporter subfamily C member 2 (ABCC2) -based insect cell expression system. Int. J. Biol. Macromol. 2022, 209, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Heckel, D. Insecticide resistance after silent spring. Science 2012, 337, 1612–1614. [Google Scholar] [CrossRef] [Green Version]
- Dechklar, M.; Tiewsiri, K.; Angusuthanasombat, C.; Pootanakit, K. Functional expression in insect cells of glycosylphosphatidylinositol-linked alkaline phosphatase from Aedes aegypti larval midgut: A Bacillus thuringiensis Cry4Ba toxin receptor. Insect Biochem. Mol. Biol. 2011, 41, 159–166. [Google Scholar] [CrossRef]
- Xie, C.; Xiong, L.; Ye, M.; Shen, L.; Li, J.; Zhang, Z.; You, M.; You, S. Genome-wide analysis of V-ATPase genes in Plutella xylostella (L.) and the potential role of PxVHA-G1 in resistance to Bacillus thuringiensis Cry1Ac toxin. Int. J. Biol. Macromol. 2021, 194, 74–83. [Google Scholar] [CrossRef]
- Mauchamp, B.; Royer, C.; Garel, A.; Jalabert, A.; Da Rocha, M.; Grenier, A.M.; Labas, V.; Vinh, J.; Mita, K.; Kadono, K.; et al. Polycalin (chlorophyllid A binding protein): A novel, very large fluorescent lipocalin from the midgut of the domestic silkworm Bombyx mori L. Insect Biochem. Mol. Biol. 2006, 36, 623–633. [Google Scholar] [CrossRef]
- Angelucci, C.; Barrett-Wilt, G.A.; Hunt, D.F.; Akhurst, R.J.; East, P.D.; Gordon, K.H.; Campbell, P.M. Diversity of aminopeptidases, derived from four lepidopteran gene duplications, and polycalins expressed in the midgut of Helicoverpa armigera: Identification of proteins binding the delta-endotoxin, Cry1Ac of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2008, 38, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.J.; Wei, J.Z.; Wang, Y.A.; Chen, L.; Liang, G.M. Polycalin is involved in the toxicity and resistance to Cry1Ac toxin in Helicoverpa armigera (Hubner). Arch. Insect Biochem. Physiol. 2020, 104, e21661. [Google Scholar] [CrossRef]
- Wang, B.-J.; Wang, Y.-N.; Wei, J.-Z.; Liu, C.; Chen, L.; Khaing, M.M.; Liang, G.-M. Polycalin is involved in the action mechanism of Cry2Aa toxin in Helicoverpa armigera (Hübner). J. Integr. Agric. 2019, 18, 627–635. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, Z.; Liu, Y.; Liang, G.; Shu, C.; Song, F.; Zhou, X.; Bravo, A.; Soberón, M.; Zhang, J.J.M. Identification of ABCC2 as a binding protein of Cry1Ac on brush border membrane vesicles from Helicoverpa armigera by an improved pull-down assay. Microbiologyopen 2016, 5, 659–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandian, G.; Ishikawa, T.; Togashi, M.; Shitomi, Y.; Haginoya, K.; Yamamoto, S.; Nishiumi, T.; Hori, H. Bombyx mori midgut membrane protein P252, which binds to Bacillus thuringiensis Cry1A, is a chlorophyllide-binding protein, and the resulting complex has antimicrobial activity. Appl. Environ. Microbiol. Rep. 2008, 74, 1324–1331. [Google Scholar] [CrossRef] [Green Version]
- Pandian, G.; Ishikawa, T.; Vaijayanthi, T.; Hossain, D.; Yamamoto, S.; Nishiumi, T.; Angsuthanasombat, C.; Haginoya, K.; Mitsui, T.; Hori, H. Formation of macromolecule complex with Bacillus thuringiensis Cry1A toxins and chlorophyllide binding 252-kDa lipocalin-like protein locating on Bombyx mori midgut membrane. J. Membr. Biol. 2010, 237, 125–136. [Google Scholar] [CrossRef]
- Lin, M.M.; Liu, Y.; Zhang, X.; Zhong, J.F.; Hu, X.D.; Xu, C.J.; Xie, Y.X.; Zhang, C.Z.; Liang, Y.; Liu, X.J.; et al. Anti-idiotypic single-chain variable fragment antibody partially mimic the functionally spatial structure of Cry2Aa toxin. Anal. Biochem. 2021, 625, 114222. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, B.Y.; Liu, L.; Ma, W.H.; Wang, X.P.; Lei, C.L.; Chen, L.Z. Proteomic analysis of Cry2Aa-binding proteins and their receptor function in Spodoptera exigua. Sci. Rep. 2017, 7, 40222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, E.L.; Du, X.; Zhao, A.P.; Sun, C.; Liu, T.X.; Li, Y.P. Gene cloning and expression of polycalin protein from Plutella xylostella (Lepidoptera: Plutellidae) and its binding characteristics with Cry1Ac toxin. Acta Entomol. Sin. 2017, 60, 1155–1167. [Google Scholar]
- Zhu, Q.; Gao, M.J.; Lu, L.N.; Liu, X.J. Synergism of Bacillus thuringiensis toxin Cry1Ac by a fragment of toxin-binding polycalin from Plutella xylostella. J. Agric. Food Chem. 2021, 69, 11816–11824. [Google Scholar] [CrossRef]
- Pardo-Lopez, L.; Soberon, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Gómez, I.; Sánchez, J.; Muñoz-Garay, C.; Matus, V.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis Cry1A toxins are versatile proteins with multiple modes of action: Two distinct pre-pores are involved in toxicity. Biochemical 2014, 459, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xu, D.; Huang, Y.; Zhou, H.; Liu, W.; Cong, S.; Wang, J.; Li, W.; Wan, P. Mutation in the cadherin gene is a key factor for pink bollworm resistance to Bt cotton in China. Toxins 2022, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Yaoi, K.; Kadotani, T.; Kuwana, H.; Shinkawa, A.; Takahashi, T.; Iwahana, H.; Sato, R. Aminopeptidase N from Bombyx Mori as a candidate for the receptor of Bacillus thuringiensis Cry1Aa toxin. Eur. J. Biochem. 1997, 246, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, S.; Rajagopal, R.; Venkatesh, G.R.; Srivastava, A.; Bhatnagar, R.K. Knockdown of aminopeptidase-N from Helicoverpa armigera larvae and in transfected Sf21 cells by RNA interference reveals its functional interaction with Bacillus thuringiensis insecticidal protein Cry1Ac. J. Biol. Chem. 2007, 282, 7312–7319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, C.; Wu, K.; Liu, C.; Gao, Y.; Jurat-Fuentes, J.L.; Gao, X. Characterization of a Cry1Ac toxin-binding alkaline phosphatase in the midgut from Helicoverpa armigera (Hubner) larvae. J. Insect Physiol. 2010, 56, 666–672. [Google Scholar] [CrossRef]
- Tanaka, S.; Endo, H.; Adegawa, S.; Iizuka, A.; Imamura, K.; Kikuta, S.; Sato, R. Bombyx mori ABC transporter C2 structures responsible for the receptor function of Bacillus thuringiensis Cry1Aa toxin. Insect Biochem. Mol. Biol. 2017, 91, 44–54. [Google Scholar] [CrossRef]
- Martinez-Solis, M.; Pinos, D.; Endo, H.; Portugal, L.; Sato, R.; Ferre, J.; Herrero, S.; Hernandez-Martinez, P. Role of Bacillus thuringiensis Cry1A toxins domains in the binding to the ABCC2 receptor from Spodoptera exigua. Insect Biochem. Mol. Biol. 2018, 101, 47–56. [Google Scholar] [CrossRef]
- Yang, Y.; Duan, A.; Zhang, C.; Zhang, Y.; Wang, A.; Xue, C.; Wang, H.; Zhao, M.; Zhang, J. Overexpression of ATP-binding cassette transporters ABCG10, ABCH3 and ABCH4 in Aphis craccivora (Koch) facilitates its tolerance to imidacloprid. Pestic. Biochem. Physiol. 2022, 186, 105170. [Google Scholar] [CrossRef]
- Guo, Z.J.; Kang, S.; Zhu, X.; Xia, J.X.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhang, Y.J. Down-regulation of a novel ABC transporter gene (Pxwhite) is associated with Cry1Ac resistance in the diamondback moth, Plutella xylostella (L.). Insect Biochem. Mol. Biol. 2015, 59, 30–40. [Google Scholar] [CrossRef]
- Stevens, T.; Song, S.; Bruning, J.B.; Choo, A.; Baxter, S.W. Expressing a moth abcc2 gene in transgenic Drosophila causes susceptibility to Bt Cry1Ac without requiring a cadherin-like protein receptor. Insect Biochem. Mol. Biol. 2017, 80, 61–70. [Google Scholar] [CrossRef]
- Zhou, J.L.; Guo, Z.J.; Kang, S.; Qin, J.Y.; Gong, L.J.; Sun, D.; Guo, L.; Zhu, L.H.; Bai, Y.; Zhang, Z.Z.; et al. Reduced expression of the P-glycoprotein gene PxABCB1 is linked to resistance to Bacillus thuringiensis Cry1Ac toxin in Plutella xylostella (L.). Pest Manag. Sci. 2020, 76, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Guney, G.; Cedden, D.; Hanniger, S.; Heckel, D.G.; Coutu, C.; Hegedus, D.D.; Mutlu, D.A.; Suludere, Z.; Sezen, K.; Guney, E.; et al. Silencing of an ABC transporter, but not a cadherin, decreases the susceptibility of Colorado potato beetle larvae to Bacillus thuringiensis ssp. tenebrionis Cry3Aa toxin. Arch. Insect Biochem. Physiol. 2021, 108, e21834. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, C.; Chen, G.; Li, Y.; Yang, X.; Han, L.; Peng, Y. Downregulation of the CsABCC2 gene is associated with Cry1C resistance in the striped stem borer Chilo suppressalis. Pestic. Biochem. Physiol. 2022, 184, 105119. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, W.; Song, X.; Ma, X.; Cotto-Rivera, R.O.; Kain, W.; Chu, H.; Chen, Y.R.; Fei, Z.; Wang, P. Mutation of ABC transporter ABCA2 confers resistance to Bt toxin Cry2Ab in Trichoplusia ni. Insect Biochem. Mol. Biol. 2019, 112, 103209. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.A.; LeRoy, D.M.; Mathew, L.G.; Wu, Y.; Unnithan, G.C.; Yelich, A.J.; Carriere, Y.; Li, X.; Tabashnik, B.E. CRISPR-mediated mutations in the ABC transporter gene ABCA2 confer pink bollworm resistance to Bt toxin Cry2Ab. Sci. Rep. 2021, 11, 10377. [Google Scholar] [CrossRef]
- Jin, W.; Zhai, Y.; Yang, Y.; Wu, Y.; Wang, X. Cadherin protein is involved in the action of Bacillus thuringiensis Cry1Ac toxin in Ostrinia furnacalis. Toxins 2021, 13, 658. [Google Scholar] [CrossRef]
- Jin, M.; Cheng, Y.; Guo, X.; Li, M.; Chakrabarty, S.; Liu, K.; Wu, K.; Xiao, Y. Down-regulation of lysosomal protein ABCB6 increases gossypol susceptibility in Helicoverpa armigera. Insect Biochem. Mol. Biol. 2020, 122, 103387. [Google Scholar] [CrossRef]
- Shelton, A.M.; Cooley, R.J.; Kroening, M.K.; Wilsey, W.T.; Eigenbrode, S.D. Comparative analysis of two rearing procedures for diamondback moth (Lepidoptera: Plutellidae). J. Entomol. Sci. 1991, 26, 17–26. [Google Scholar] [CrossRef]
- Xiong, L.; Liu, Z.X.; Shen, L.L.; Xie, C.; Ye, M.; Li, Z.Y.; Zhang, Z.; Li, J.G.; Dong, Y.; You, M.S.; et al. A novel reference for Bt-resistance mechanism in Plutella xylostella based on analysis of the midgut transcriptomes. Insects 2021, 12, 1091. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Heckel, D.G.; Gahan, L.J.; Liu, Y.B.; Tabashnik, B.E. Genetic mapping of resistance to Bacillus thuringiensis toxins in diamondback moth using biphasic linkage analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 8373–8377. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation | N a | Genotype b | |
|---|---|---|---|
| RPoSPo | RPoRPo | ||
| F1 (G88 ♀ × Cry1S1000 ♂) | 10 | 10 | 0 |
| Backcross family a with toxin treated | 12 | 9 | 3 |
| Backcross family a without toxin treated | 24 | 11 | 13 |
| Backcross family b with toxin treated | 12 | 3 | 9 |
| Backcross family b without toxin treated | 24 | 12 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, L.; Liu, Z.; Li, J.; Yao, S.; Li, Z.; Chen, X.; Shen, L.; Zhang, Z.; Li, Y.; Hou, Q.; et al. Analysis of the Effect of Plutella xylostella Polycalin and ABCC2 Transporter on Cry1Ac Susceptibility by CRISPR/Cas9-Mediated Knockout. Toxins 2023, 15, 273. https://doi.org/10.3390/toxins15040273
Xiong L, Liu Z, Li J, Yao S, Li Z, Chen X, Shen L, Zhang Z, Li Y, Hou Q, et al. Analysis of the Effect of Plutella xylostella Polycalin and ABCC2 Transporter on Cry1Ac Susceptibility by CRISPR/Cas9-Mediated Knockout. Toxins. 2023; 15(4):273. https://doi.org/10.3390/toxins15040273
Chicago/Turabian StyleXiong, Lei, Zhaoxia Liu, Jingge Li, Shuyuan Yao, Zeyun Li, Xuanhao Chen, Lingling Shen, Zhen Zhang, Yongbin Li, Qing Hou, and et al. 2023. "Analysis of the Effect of Plutella xylostella Polycalin and ABCC2 Transporter on Cry1Ac Susceptibility by CRISPR/Cas9-Mediated Knockout" Toxins 15, no. 4: 273. https://doi.org/10.3390/toxins15040273