Probing the Interactions of 31 Mycotoxins with Xanthine Oxidase: Alternariol, Alternariol-3-Sulfate, and α-Zearalenol Are Allosteric Inhibitors of the Enzyme

 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
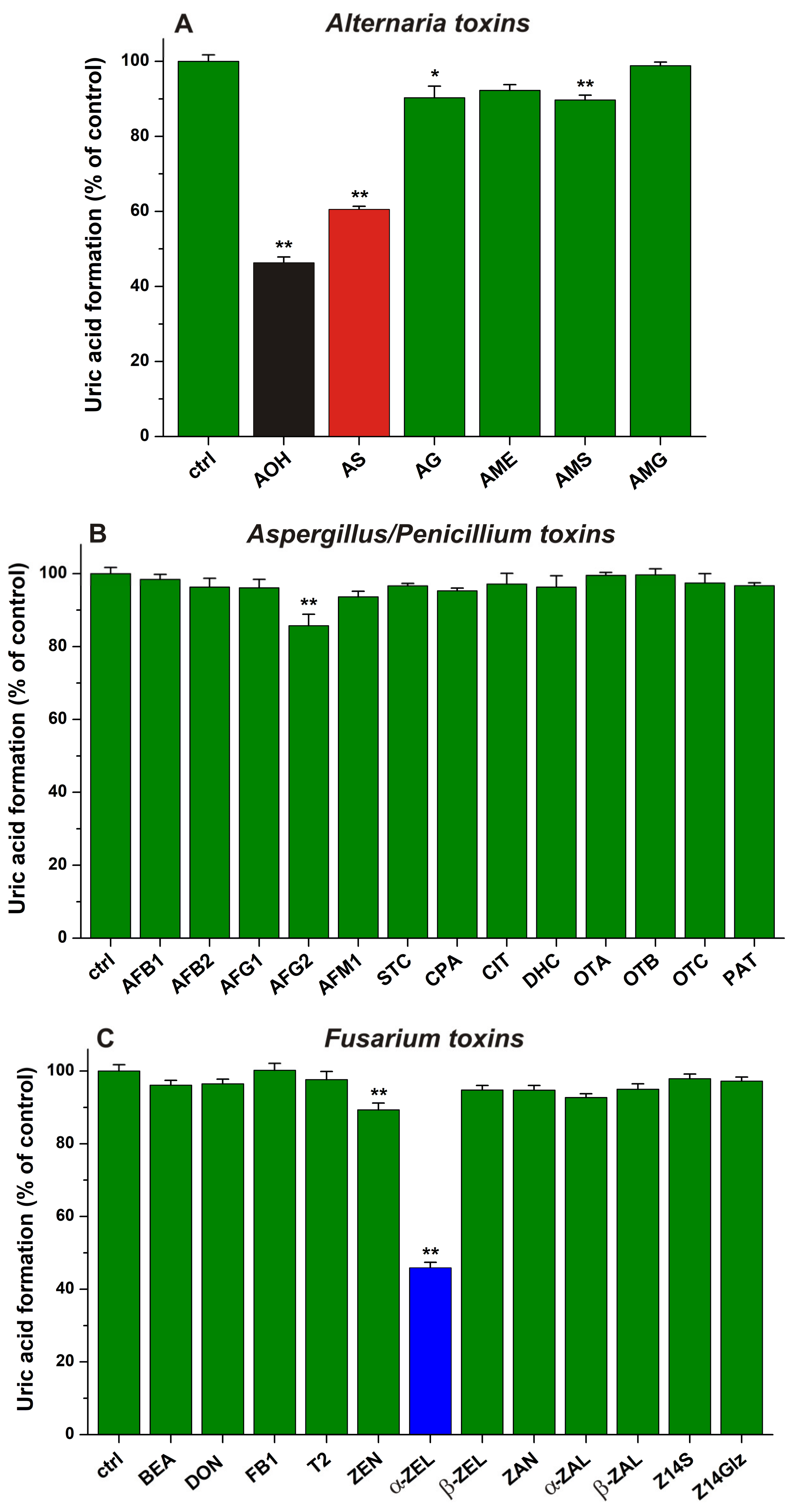
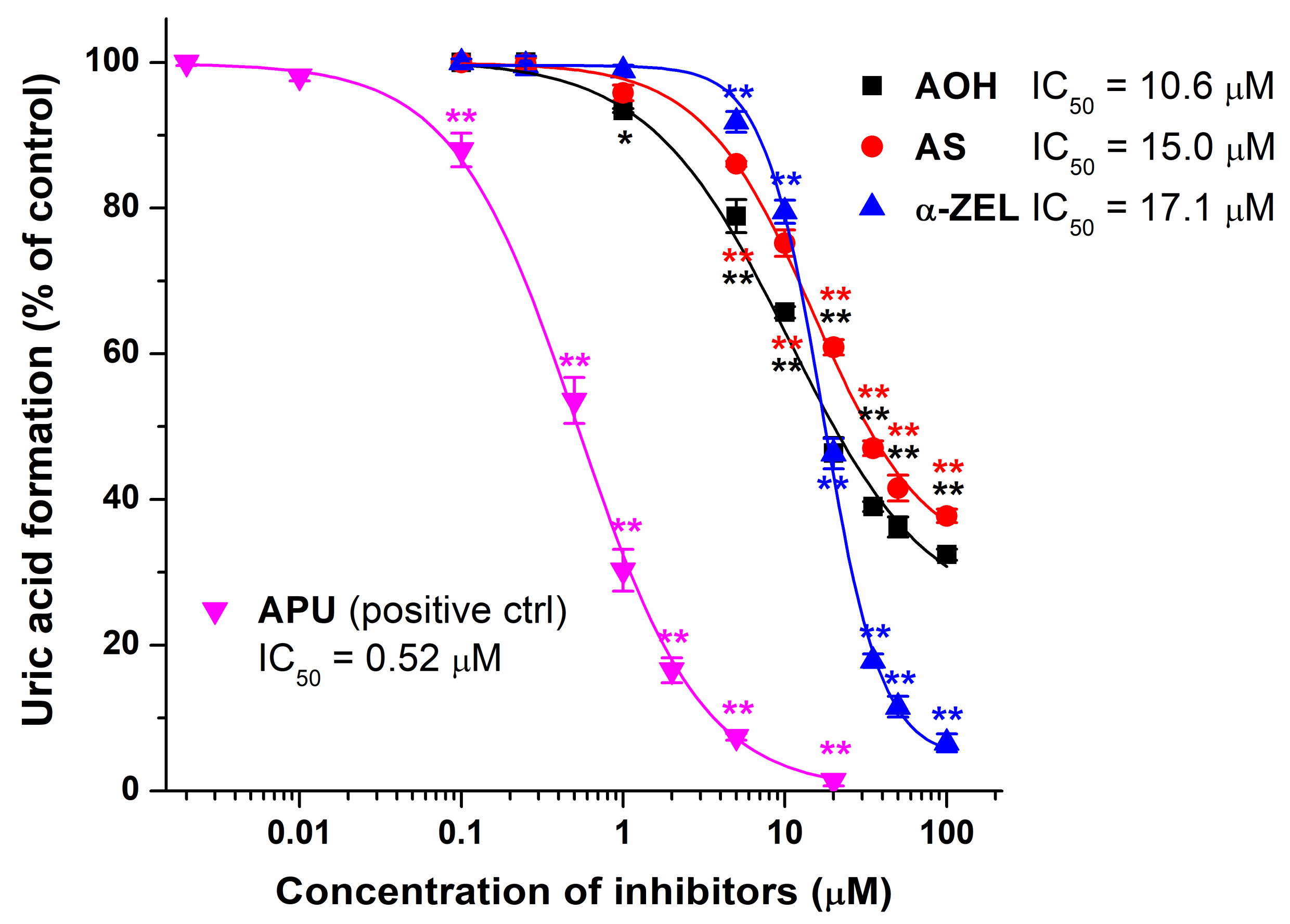
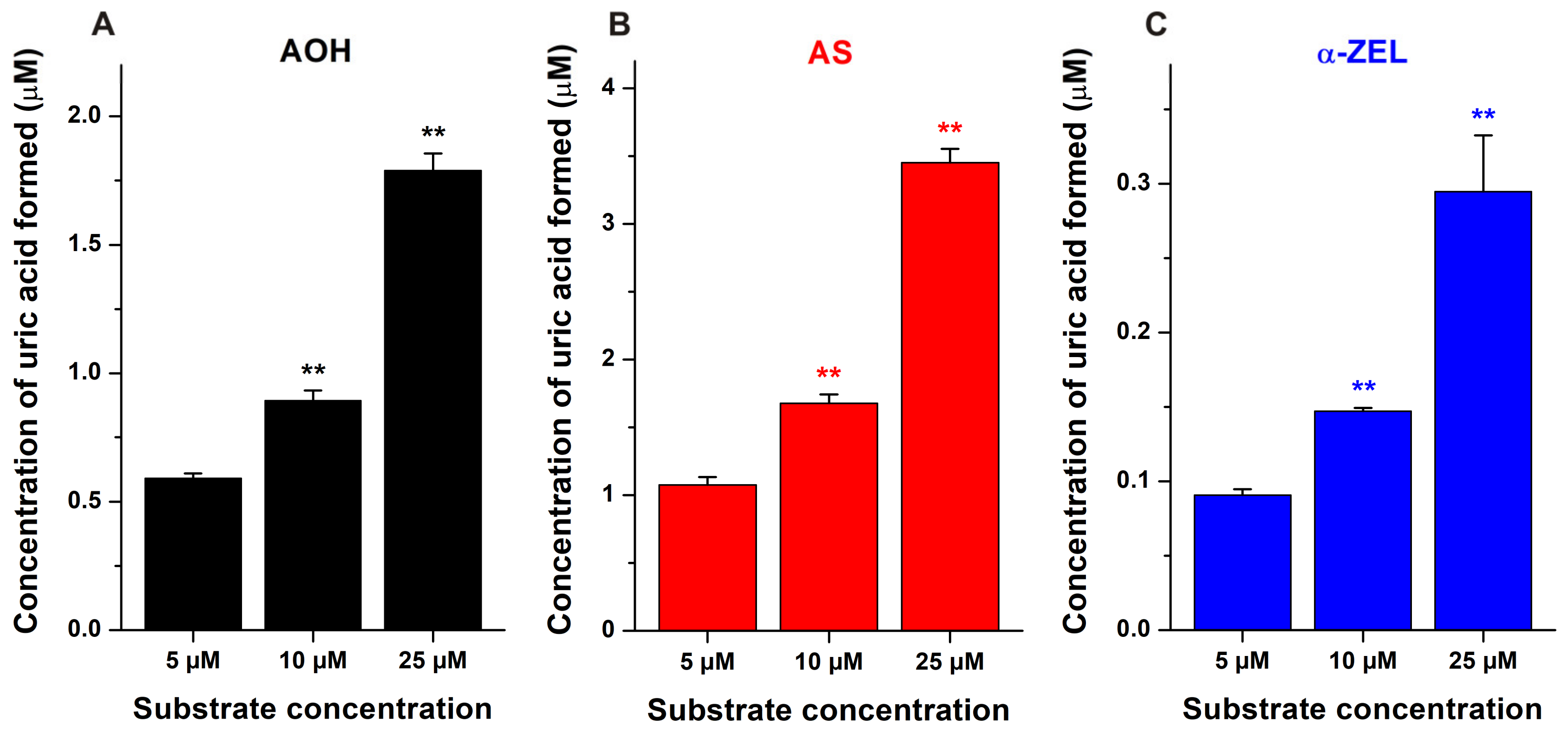
2.1. Inhibitory Effects of Mycotoxins on Xanthine Oxidase Enzyme
2.2. Mycotoxin Depletion Assays
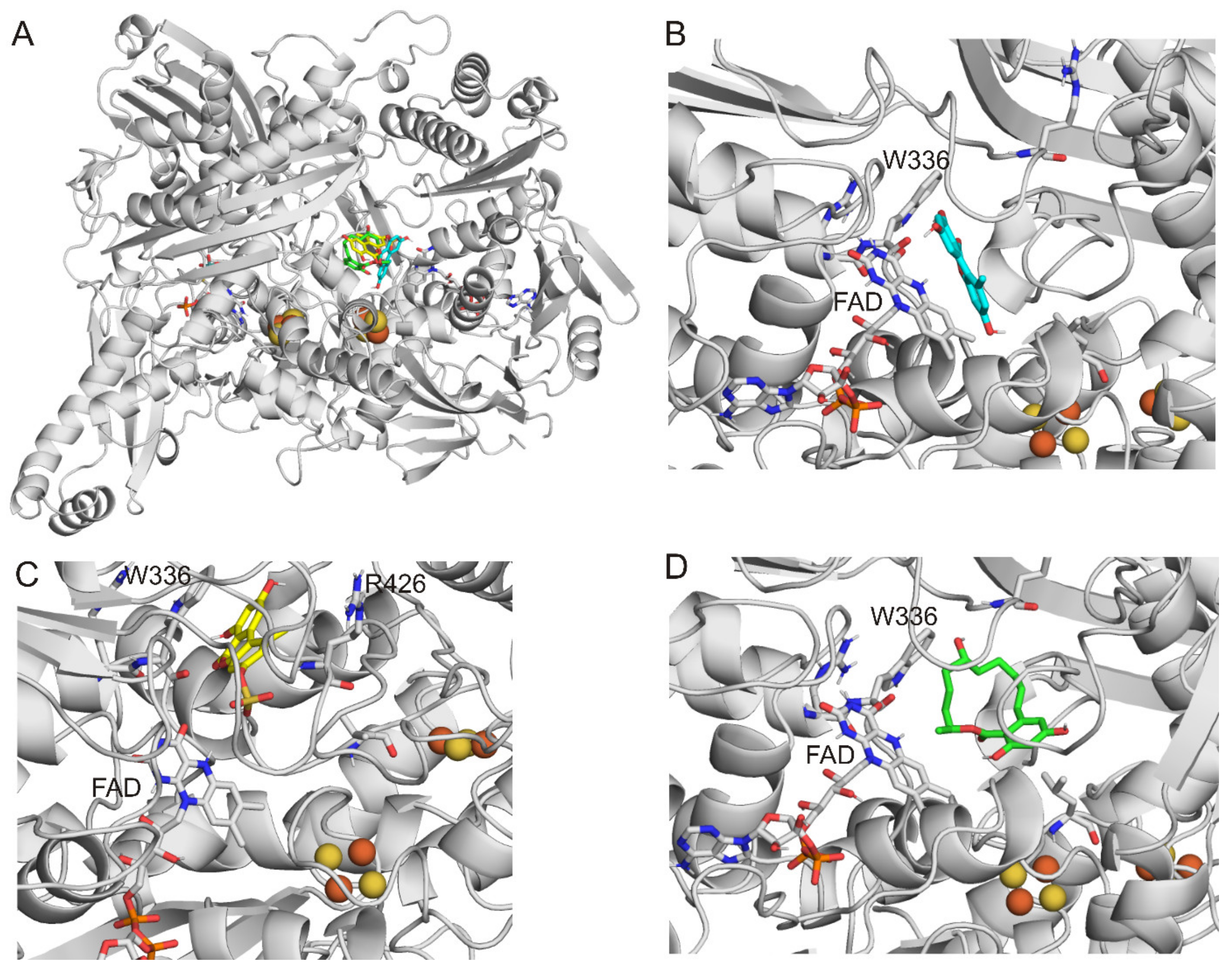
2.3. Modeling Studies
3. Conclusions
4. Materials and Methods
4.1. Reagents
4.2. Xanthine Oxidase Assay
4.3. Mycotoxin Depletion Assays
4.4. HPLC Analyses
4.5. Modeling Studies
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da Rocha, M.E.B.; Freire, F.C.O.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 59–165. [Google Scholar]
- Kabak, B. The fate of mycotoxins during thermal food processing. J. Sci. Food Agric. 2009, 89, 549–554. [Google Scholar] [CrossRef]
- Milani, J.; Maleki, G. Effects of processing on mycotoxin stability in cereals. J. Sci. Food Agric. 2014, 94, 2372–2375. [Google Scholar] [CrossRef]
- Lootens, O.; Vermeulen, A.; Croubels, S.; De Saeger, S.; Van Bocxlaer, J.; De Boevre, M. Possible Mechanisms of the Interplay between Drugs and Mycotoxins—Is There a Possible Impact? Toxins 2022, 14, 873. [Google Scholar] [CrossRef]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asam, S.; Devos, T.; Njumbe Ediage, E.; Diana Di Mavungu, J.; Jacxsens, L.; Van Landschoot, A.; Vanhaecke, L.; et al. Validated UPLC-MS/MS Methods to Quantitate Free and Conjugated Alternaria Toxins in Commercially Available Tomato Products and Fruit and Vegetable Juices in Belgium. J. Agric. Food Chem. 2016, 64, 5101–5109. [Google Scholar] [CrossRef] [Green Version]
- Puntscher, H.; Cobankovic, I.; Marko, D.; Warth, B. Quantitation of Free and Modified Alternaria Mycotoxins in European Food Products by LC-MS/MS. Food Control 2019, 102, 157–165. [Google Scholar] [CrossRef]
- Qin, Q.; Fan, Y.; Jia, Q.; Duan, S.; Liu, F.; Jia, B.; Wang, G.; Guo, W.; Wang, C. The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes. Toxins 2022, 14, 827. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the Risks for Animal and Public Health Related to the Presence of Alternaria Toxins in Feed and Food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Aichinger, G.; Del Favero, G.; Warth, B.; Marko, D. Alternaria toxins—Still emerging? Compr. Rev. Food Sci. Food Saf. 2021, 20, 4390–4406. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Klingelhöfer, D.; Zhu, Y.; Braun, M.; Bendels, M.H.K.; Brüggmann, D.; Groneberg, D.A. Aflatoxin—Publication analysis of a global health threat. Food Control 2018, 89, 280–290. [Google Scholar] [CrossRef]
- Nieto, C.H.D.; Granero, A.M.; Zon, M.A.; Fernández, H. Sterigmatocystin: A mycotoxin to be seriously considered. Food Chem. Toxicol. 2018, 118, 460–470. [Google Scholar] [CrossRef]
- Ostry, V.; Toman, J.; Grosse, Y.; Malir, F. Cyclopiazonic acid: 50th anniversary of its discovery. World Mycotoxin J. 2018, 11, 135–148. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA). Scientific opinion on the risks for public and animal health related to the presence of citrinin in food and feed. EFSA J. 2012, 10, 2605. [Google Scholar]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef]
- Degen, G.H.; Ali, N.; Gundert-Remy, U. Preliminary data on citrinin kinetics in humans and their use to estimate citrinin exposure based on biomarkers. Toxicol. Lett. 2018, 282, 43–48. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, 6113. [Google Scholar]
- Di Stefano, V.; Avellone, G.; Pitonzo, R.; Capocchiano, V.G.; Mazza, A.; Cicero, N.; Dugo, G. Natural co-occurrence of ochratoxin A, ochratoxin B and aflatoxins in Sicilian red wines. Food Addit. Contam. A 2015, 32, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Remiro, R.; Gonzalez-Penas, E.; Lizarraga, E.; Lopez de Cerain, A. Quantification of ochratoxin A and five analogs in Navarra red wines. Food Control 2012, 27, 139–145. [Google Scholar] [CrossRef]
- Bryła, M.; Pierzgalski, A.; Zapaśnik, A.; Uwineza, P.A.; Ksieniewicz-Woźniak, E.; Modrzewska, M.; Waśkiewicz, A. Recent Research on Fusarium Mycotoxins in Maize—A Review. Foods 2022, 11, 3465. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef]
- Harrison, R. Structure and function of xanthine oxidoreductase: Where are we now? Free Radic. Biol. Med. 2002, 33, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Glantzounis, G.K.; Tsimoyiannis, E.C.; Kappas, A.M.; Galaris, D.A. Uric acid and oxidative stress. Curr. Pharm. Des. 2005, 11, 4145–4151. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.M.; Kelley, E.E.; Straub, A.C. The impact of xanthine oxidase (XO) on hemolytic diseases. Redox. Biol. 2019, 21, 101072. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Y.; Cheng, J.; Huangfu, N.; Zhao, R.; Xu, Z.; Zhang, F.; Zheng, W.; Zhang, D. Hyperuricemia and Cardiovascular Disease. Curr. Pharm. Des. 2019, 25, 700–709. [Google Scholar] [CrossRef]
- Galbusera, C.; Orth, P.; Fedida, D.; Spector, T. Superoxide radical production by allopurinol and xanthine oxidase. Biochem. Pharmacol. 2006, 71, 1747–1752. [Google Scholar] [CrossRef]
- Day, R.O.; Graham, G.G.; Hicks, M.; McLachlan, A.J.; Stocker, S.L.; Williams, K.M. Clinical Pharmacokinetics and Pharmacodynamics of Allopurinol and Oxypurinol. Clin. Pharmacokinet. 2007, 46, 623–644. [Google Scholar] [CrossRef]
- Leong, R.W.; Gearry, R.B.; Sparrow, M.P. Thiopurine hepatotoxicity in inflammatory bowel disease: The role for adding allopurinol. Expert Opin. Drug Saf. 2008, 7, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Costantini, A.V. The fungal etiology of gout and hyperuricemia: The antifungal mode of action of colchicine. Biomed. Rev. 1992, 1, 47–52. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Doleski, P.H.; Zeppenfeld, C.C.; Descovi, S.; Da Silva, A.S.; Baldisserotto, B. Xanthine oxidase activity exerts pro-oxidative and pro-inflammatory effects in serum of silver catfish fed with a diet contaminated with aflatoxin B1. J. Fish Dis. 2018, 41, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Owumi, S.E.; Najophe, S.E.; Idowu, T.B.; Nwozo, S.O. Protective mechanisms of gallic acid on hepatorenal dysfunction of zearalenone treated rat. Biologia 2021, 76, 3123–3135. [Google Scholar] [CrossRef]
- Osselaere, A.; Santos, R.; Hautekiet, V.; De Backer, P.; Chiers, K.; Ducatelle, R.; Croubels, S. Deoxynivalenol impairs hepatic and intestinal gene expression of selected oxidative stress, tight junction and inflammation proteins in broiler chickens, but addition of an adsorbing agent shifts the effects to the distal parts of the small intestine. PLoS ONE 2013, 8, e69014. [Google Scholar] [CrossRef] [Green Version]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.J.; De Baere, S.; Mountzouris, K.C.; Su, S.; Wong, E.A.; De Meulenaer, B.; et al. Mycotoxins Deoxynivalenol and Fumonisins Alter the Extrinsic Component of Intestinal Barrier in Broiler Chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Sun, S.; Lv, C.; Li, Z.; Guo, M.; Yin, Y.; Wang, H.; Wang, W. Discovery of mycotoxin alternariol as a potential lead compound targeting xanthine oxidase. Chem. Biol. Interact. 2022, 360, 109948. [Google Scholar] [CrossRef]
- Al-Harbi, S.A.; Abdulrahman, A.O.; Zamzami, M.A.; Khan, M.I. Urolithins: The Gut Based Polyphenol Metabolites of Ellagitannins in Cancer Prevention, a Review. Front. Nutr. 2021, 8, 647582. [Google Scholar] [CrossRef] [PubMed]
- Gu, W. Bioactive metabolites from Alternaria brassicicola ML-P08, an endophytic fungus residing in Malus halliana. World J. Microbiol. Biotechnol. 2009, 25, 1677–1683. [Google Scholar] [CrossRef]
- Puvaca, N.; Tanaskovic, S.; Bursic, V.; Petrovic, A.; Merkuri, J.; Kika, T.S.; Marinkovic, D.; Vukovic, G.; Cara, M. Optical Characterization of Alternaria spp. Contaminated Wheat Grain and Its Influence in Early Broilers Nutrition on Oxidative Stress. Sustainability 2021, 13, 4005. [Google Scholar] [CrossRef]
- Harrison, R. Milk xanthine oxidase: Properties and physiological roles. Int. Dairy J. 2006, 16, 546–554. [Google Scholar] [CrossRef]
- Weidenbörner, M. Natural Mycotoxin Contamination in Humans and Animals; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Mohos, V.; Pánovics, A.; Fliszár-Nyúl, E.; Schilli, G.; Hetényi, C.; Mladěnka, P.; Needs, P.W.; Kroon, P.A.; Pethő, G.; Poór, M. Inhibitory Effects of Quercetin and Its Human and Microbial Metabolites on Xanthine Oxidase Enzyme. Int. J. Mol. Sci. 2019, 20, 2681. [Google Scholar] [CrossRef] [Green Version]
- Mohos, V.; Fliszár-Nyúl, E.; Poór, M. Inhibition of Xanthine Oxidase-Catalyzed Xanthine and 6-Mercaptopurine Oxidation by Flavonoid Aglycones and Some of Their Conjugates. Int. J. Mol. Sci. 2020, 21, 3256. [Google Scholar] [CrossRef]
- Turnheim, K.; Krivanek, P.; Oberbauer, R. Pharmacokinetics and pharmacodynamics of allopurinol in elderly and young subjects. Br. J. Clin. Pharmacol. 1999, 48, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Pauff, J.M.; Cao, H.; Hille, R. Substrate Orientation and Catalysis at the Molybdenum Site in Xanthine Oxidase. J. Biol. Chem. 2009, 284, 8760–8767. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, Y.; Nishino, T.; Okamoto, K.; Matsumura, T.; Eger, B.T.; Pai, E.F.; Nishino, T. Unique Amino Acids Cluster for Switching from the Dehydrogenase to Oxidase Form of Xanthine Oxidoreductase. Proc. Natl. Acad. Sci. USA 2003, 100, 8170–8175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fliszár-Nyúl, E.; Lemli, B.; Kunsági-Máté, S.; Szente, L.; Poór, M. Interactions of Mycotoxin Alternariol with Cyclodextrins and Its Removal from Aqueous Solution by Beta-Cyclodextrin Bead Polymer. Biomolecules 2019, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemli, B.; Vilmányi, P.; Fliszár-Nyúl, E.; Zsidó, B.Z.; Hetényi, C.; Szente, L.; Poór, M. Testing Serum Albumins and Cyclodextrins as Potential Binders of the Mycotoxin Metabolites Alternariol-3-Sulfate, Alternariol-9-Monomethylether and Alternariol-9-Monomethylether-3-Sulfate. Int. J. Mol. Sci. 2022, 23, 14353. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Faisal, Z.; Zand, A.; Bencsik, T.; Lemli, B.; Kunsági-Máté, S.; Szente, L. Removal of Zearalenone and Zearalenols from Aqueous Solutions Using Insoluble Beta-Cyclodextrin Bead Polymer. Toxins 2018, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, J.; Marsili, M. Iterative Partial Equalization of Orbital Electronegativity-a Rapid Access to Atomic Charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated Docking Using a Lamarckian Genetic Algorithm and Empirical Binding Free Energy Function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Stewart, J.J.P. MOPAC; Stewart Computational Chemistry: Colorado Springs, CO, USA, 2016. [Google Scholar]
- Stewart, J.J.P. Optimization of Parameters for Semiempirical Methods VI: More Modifications to the NDDO Approximations and Re-Optimization of Parameters. J. Mol. Model. 2013, 19, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetényi, C.; Van Der Spoel, D. Blind Docking of Drug-Sized Compounds to Proteins with up to a Thousand Residues. FEBS Lett. 2006, 580, 1447–1450. [Google Scholar] [CrossRef] [Green Version]
- Hetényi, C.; Van Der Spoel, D. Toward Prediction of Functional Protein Pockets Using Blind Docking and Pocket Search Algorithms. Protein Sci. 2011, 20, 880–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zsidó, B.Z.; Börzsei, R.; Szél, V.; Hetényi, C. Determination of Ligand Binding Modes in Hydrated Viral Ion Channels to Foster Drug Design and Repositioning. J. Chem. Inf. Model. 2021, 61, 4011–4022. [Google Scholar] [CrossRef] [PubMed]
- Zsidó, B.Z.; Balog, M.; Erős, N.; Poór, M.; Mohos, V.; Fliszár-Nyúl, E.; Hetényi, C.; Nagane, M.; Hideg, K.; Kálai, T.; et al. Synthesis of Spin-Labelled Bergamottin: A Potent CYP3A4 Inhibitor with Antiproliferative Activity. Int. J. Mol. Sci. 2020, 21, 508. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | Abbreviation | Occurrence | Fungi | Toxic Effects | Reference |
|---|---|---|---|---|---|
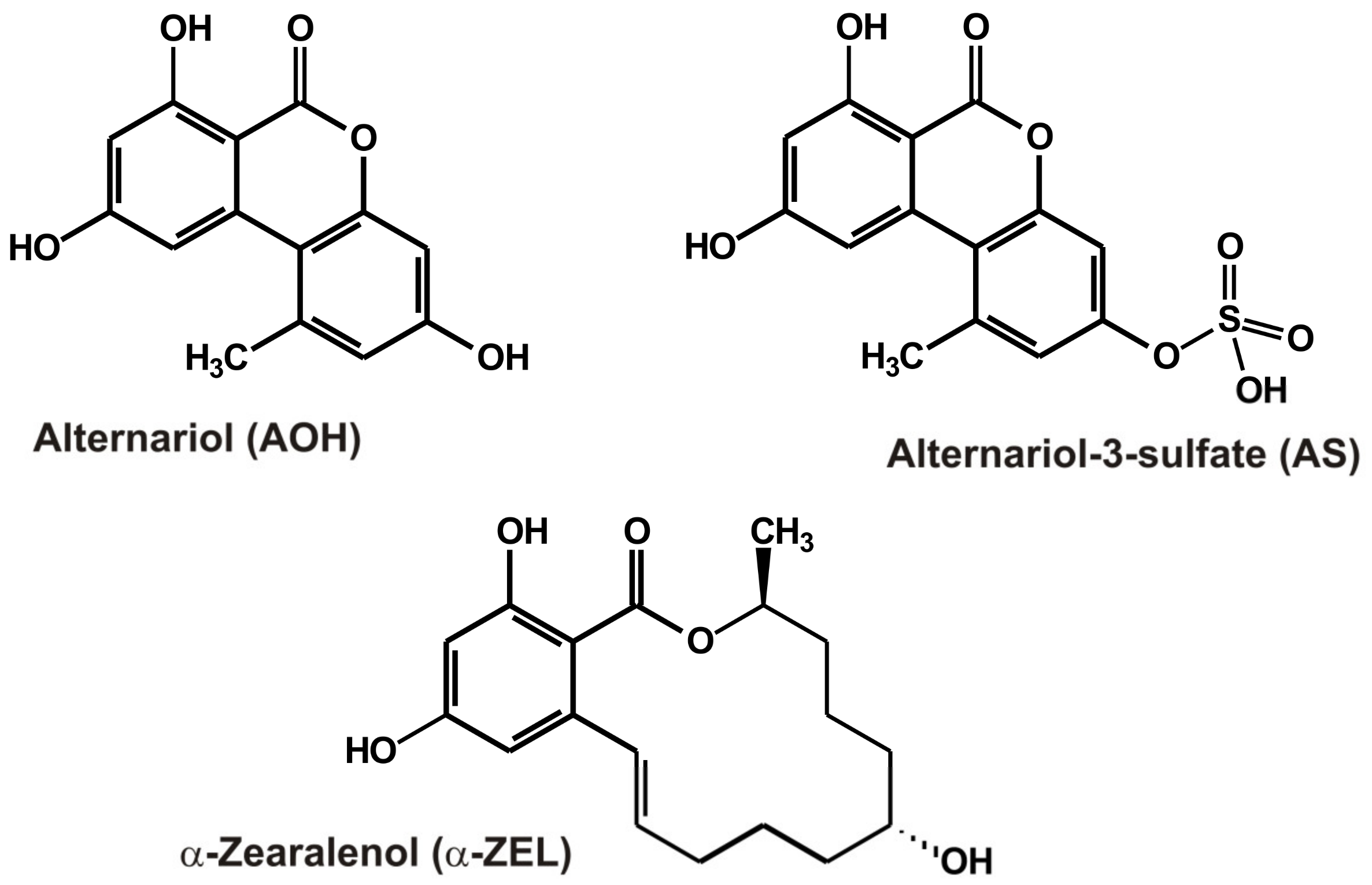
| Alternariol Alternariol-9-methylether Alternariol-3-sulfate Alternariol-3-glucoside AME-3-sulfate AME-3-glucoside | AOH AME AS AG AMS AMG | Tomatoes, grapes, and corresponding products | Alternaria sp. | Endocrine disruptor and carcinogenicity | [5,6,7,8,9] |
| Aflatoxin B1 Aflatoxin B2 Aflatoxin G1 Aflatoxin G2 Aflatoxin M1 | AFB1 AFB2 AFG1 AFG2 AFM1 | Cereals, nuts, figs, vegetables, meat, milk, and dairy products | Aspergillus sp. | Hepatotoxicity | [10,11] |
| Sterigmatocystin | STC | Grains, coffee beans, cheese, spices, and soybeans | Aspergillus sp. | Hepatotoxicity and nephrotoxicity | [12] |
| Cyclopiazonic acid | CPA | Oilseeds, cereals, meat, and milk | Aspergillus sp. Penicillium sp. | Gastrointestinal toxicity and neurotoxicity | [13] |
| Citrinin Dihydrocitrinone | CIT DHC | Grains, rice, fruits, and spices | Aspergillus sp. Penicillium sp. Monascus sp. | Nephrotoxicity | [14,16] |
| Ochratoxin A Ochratoxin B Ochratoxin C | OTA OTB OTC | Cereals, fruits, meat, dairy products, and beverages | Aspergillus sp. Penicillium sp. | Nephrotoxicity | [17,18,19] |
| Patulin | PAT | Apple, pear, and corresponding products | Aspergillus sp. Penicillium sp. | Gastrointestinal toxicity and immunotoxicity | [15] |
| Beauvericin | BEA | Cereals and corresponding products | Fusarium sp. | Low toxicity | [20] |
| Deoxynivalenol | DON | Cereals and corresponding products | Fusarium sp. | Gastrointestinal toxicity | [20] |
| Fumonisin B1 | FB1 | Cereals and corresponding products | Fusarium sp. | Neural tube defects | [20] |
| T-2 toxin | T2 | Cereals and corresponding products | Fusarium sp. | Gastrointestinal toxicity and toxic aleukia | [20] |
| Zearalenone α-Zearalenol β-Zearalenol Zearalanone α-Zearalanol β-Zearalanol Zearalenone-14-sulfate Zearalenone-14-glucoside | ZEN α-ZEL β-ZEL ZAN α-ZAL β-ZAL Z14S Z14Glz | Cereals and corresponding products | Fusarium sp. | Xenoestrogen, endocrine disruptor | [20,21,22] |
| Incubation Time (min) | AOH Concentration (%) | AS Concentration (%) | α-ZEL Concentration (%) |
|---|---|---|---|
| 0 | 100.0 ± 1.0 | 100.0 ± 0.1 | 100.0 ± 0.2 |
| 60 | 102.0 ± 0.9 | 100.3 ± 1.4 | 98.9 ± 0.9 |
| 120 | 96.4 ± 1.0 | 99.9 ± 1.0 | 100.0 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balázs, O.; Dombi, Á.; Zsidó, B.Z.; Hetényi, C.; Vida, R.G.; Poór, M. Probing the Interactions of 31 Mycotoxins with Xanthine Oxidase: Alternariol, Alternariol-3-Sulfate, and α-Zearalenol Are Allosteric Inhibitors of the Enzyme. Toxins 2023, 15, 250. https://doi.org/10.3390/toxins15040250
Balázs O, Dombi Á, Zsidó BZ, Hetényi C, Vida RG, Poór M. Probing the Interactions of 31 Mycotoxins with Xanthine Oxidase: Alternariol, Alternariol-3-Sulfate, and α-Zearalenol Are Allosteric Inhibitors of the Enzyme. Toxins. 2023; 15(4):250. https://doi.org/10.3390/toxins15040250
Chicago/Turabian StyleBalázs, Orsolya, Ágnes Dombi, Balázs Zoltán Zsidó, Csaba Hetényi, Róbert György Vida, and Miklós Poór. 2023. "Probing the Interactions of 31 Mycotoxins with Xanthine Oxidase: Alternariol, Alternariol-3-Sulfate, and α-Zearalenol Are Allosteric Inhibitors of the Enzyme" Toxins 15, no. 4: 250. https://doi.org/10.3390/toxins15040250