Defensive Molecules Momilactones A and B: Function, Biosynthesis, Induction and Occurrence


Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, Miki, Kagawa 761-0795, Japan
Toxins 2023, 15(4), 241; https://doi.org/10.3390/toxins15040241
Submission received: 24 February 2023
/
Revised: 22 March 2023
/
Accepted: 24 March 2023
/
Published: 25 March 2023
(This article belongs to the Special Issue Biological Activities and Potential Applications of Phytotoxins)
Abstract
:Labdane-related diterpenoids, momilactones A and B were isolated and identified in rice husks in 1973 and later found in rice leaves, straws, roots, root exudate, other several Poaceae species and the moss species Calohypnum plumiforme. The functions of momilactones in rice are well documented. Momilactones in rice plants suppressed the growth of fungal pathogens, indicating the defense function against pathogen attacks. Rice plants also inhibited the growth of adjacent competitive plants through the root secretion of momilactones into their rhizosphere due to the potent growth-inhibitory activity of momilactones, indicating a function in allelopathy. Momilactone-deficient mutants of rice lost their tolerance to pathogens and allelopathic activity, which verifies the involvement of momilactones in both functions. Momilactones also showed pharmacological functions such as anti-leukemia and anti-diabetic activities. Momilactones are synthesized from geranylgeranyl diphosphate through cyclization steps, and the biosynthetic gene cluster is located on chromosome 4 of the rice genome. Pathogen attacks, biotic elicitors such as chitosan and cantharidin, and abiotic elicitors such as UV irradiation and CuCl2 elevated momilactone production through jasmonic acid-dependent and independent signaling pathways. Rice allelopathy was also elevated by jasmonic acid, UV irradiation and nutrient deficiency due to nutrient competition with neighboring plants with the increased production and secretion of momilactones. Rice allelopathic activity and the secretion of momilactones into the rice rhizosphere were also induced by either nearby Echinochloa crus-galli plants or their root exudates. Certain compounds from Echinochloa crus-galli may stimulate the production and secretion of momilactones. This article focuses on the functions, biosynthesis and induction of momilactones and their occurrence in plant species.
Keywords:
allelopathy; biosynthesis; diterpenoid; Echinochloa crus-galli; elicitation; momilactone; Oryza sativa; pathogen; rice blastKey Contribution: The allelopathic and defense functions of momilactones may play important ecological roles in rice evolution because of the existence of a dedicated biosynthetic gene cluster in the rice genome. The potential of momilactones to serve as natural fungicides and herbicides provides significant benefits when applied to other important crops.
1. Introduction
Labdane-related diterpenoids, momilactones A and B (Figure 1) were first isolated and identified in rice husks as potent germination and growth-inhibitory substances in 1973 [1]. Momilactones were later isolated from rice leaves as phytoalexins against fungal pathogens such as the rice blast fungus Magnaporthe oryzae [2,3]. The concentrations of momilactones increased 2 days after infection with Magnaporthe oryzae, and momilactones suppressed the further growth of the fungus [4,5]. The fungal elicitors chitosan and cholic acid also induced the accumulation of momilactone A in rice leaves and suspension-cultured rice cells [6,7].
The function of momilactones, especially momilactone A, as phytoalexins has been extensively studied, and the evidence suggests that momilactones may play a role in the rice defense function against fungal pathogens [8,9,10].
The first finding of rice allelopathy was made in field examinations in Arkansas, U.S.A., where 191 of over 5000 rice accessions suppressed the growth of the aquatic weed Heteranthera limosa [11]. Allelopathy is defined as the chemical interactions among various plant species [12]. Certain plants release some secondary metabolites, termed allelochemicals, into their immediate environment, and these allelochemicals affect the growth and development of other plant species nearby [13,14,15,16,17]. The observation of rice allelopathy led to large field screening programs. Among over 16,000 rice germplasm collections of the USDA-ARS from 99 countries, 412 rice accessions suppressed the growth of Heteranthera limosa, and 145 rice accessions suppressed the growth of Ammannia coccinea [18,19]. More than 40 rice cultivars among 1000 rice collections inhibited the growth of Echinochloa crus-galli and Cyperus difformis [20]. Screening programs in the field and/or laboratories have also been carried out in several other countries, and it was found that certain rice cultivars released allelochemicals from their root systems into their immediate environments, such as rhizosphere soil, cultural solutions and other incubation media [21,22,23,24]. Thereafter, momilactones A and B were again isolated and identified in rice root exudates as rice allelochemicals [25,26]. It was also found that rice plants released momilactones throughout their life cycles with sufficient amounts of momilactones for allelopathy [27,28].
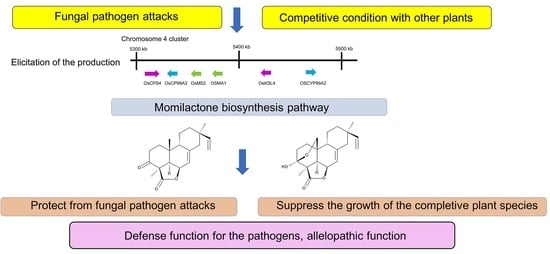
Momilactones are synthesized in rice plants from geranylgeranyl diphosphate, which is also a precursor of other phytoalexins and a plant hormone, gibberellic acid [29]. Momilactones are synthesized and accumulated in rice leaves as phytoalexins and secreted into their root zones as allelochemicals [30,31]. A gene cluster related to momilactone synthesis was found on chromosome 4 of the rice genome. Momilactones were later found in some other Gramineae plant species and the moss species Calohypnum plumiforme (syn. Hypnum plumaeform) as allelochemicals [32,33,34,35]. This review provides an overview of the functions, biosynthesis, induction and occurrence of momilactones in plant species and highlights the importance of momilactones.
2. Defense Function against Pathogens, Microbes and Insects
2.1. Rice Blast Fungal Pathogen
Infection with the rice blast pathogen Magnaporthe oryzae (syn. Pyricularia oryzae; renamed from Magnaporthe grisea) induced momilactone A accumulation in rice leaves. The accumulation was abundant at the edges of necrotic lesions, which are symptoms of the infection of leaves [36]. Blast fungus susceptibility diffed among rice cultivars, and tolerance to the fungus correlated positively with momilactone A accumulation in rice leaves [37]. Blast-fungus-resistant rice mutants accumulated momilactone A 2 days after fungus inoculation, and the concentration of momilactone A was 100–400-fold greater than that in wild-type rice and suppressed the further growth of the fungus [4,5]. Exogenously applied momilactone A also suppressed the growth of the fungus on agar media [5]. In addition, the susceptibility of momilactone-deficient rice mutants to the blast fungus was high compared to wild-type rice [38]. These observations suggest that momilactone A may prevent the subsequent spread of the fungus infection through the increased production of momilactone A after pathogen infection.
2.2. Other Fungal Pathogens
Momilactones A and B inhibited the growth of the pathogenic fungi Rhizoctonia solani, Blumeria graminis, Fusarium oxysporum, Fusarium solani, Botrytis cinereal and the Colletrichum gloesporides complex [39,40]. Infection with Xanthomonas oryzae pv. oryzae, which causes bacterial blight, increased jasmonic acid and momilactone A concentrations in rice leaves [41]. Jasmonic acid is a plant defense signaling hormone and induces several defense responses for protection [42,43,44].
2.3. Anti-Microbe Activity
Momilactone A inhibited the mycelia growth of the mushroom Coprinus cinereus [45] and the cyanobacteria Microcystis aeruginosa [46]. Momilactones A and B inhibited the growth of the bacteria Escherichia coli, Pseudomonas putida (former name, Pseudomonas ovalis), Bacillus cereus and Bacillus pumilus [39].
2.4. Insect Attack
An insect attack by the white-back planthopper (Sogatella furcifera) induced the accumulation of momilactone A in rice leaves through a jasmonic acid-mediated pathway [47]. The jasmonic acid-mediated pathway is described in Section 7. The digestive waste of the rice brown planthopper (Nilaparvata lugens) induced momilactone A and B accumulation in rice leaves. Filtration and heat treatments of digestive wastes reduced their accumulation. A symbiont of the insect, Serratia marcescens, in the digestive waste also induced the accumulation of momilactones A and B [48]. The function of momilactones A and B against insect attacks is not clear.
3. Function in Allelopathy
A considerable number of rice accessions or cultivars have been found to suppress the growth of several other plant species, including weed species, when these rice and other plants were grown together under field and/or laboratory conditions [11,21,22,23,24,49]. These observations suggest that rice is allelopathic and contains allelochemicals. A compound causing the growth-inhibitory effect of rice was later isolated from its root exudates and identified as momilactone B [25]. Momilactone A was also identified in rice secretory fluid [26]. These investigations suggest that momilactones A and B may function as rice allelochemicals.
3.1. Activities of Momilactones A and B as Allelochemicals
Momilactones A and B inhibited the growth of several plant species, including weed species such as Echinochloa crus-galli and Echinochloa colonum. Both Echinochloa species are known as the most noxious weeds in rice fields because of their potential to significantly disturb rice production [50,51]. Momilactones A and B inhibited the root and shoot growth of Echinochloa crus-galli at concentrations greater than 3 μM and 1 μM, respectively, and the root and shoot growth of Echinochloa colonum at concentrations greater than 10 μM and 1 μM, respectively [52]. Table 1 shows the concentrations of momilactones A and B required for 50% growth inhibition (defined as IC50) of target plant species. Smaller values of IC50 indicate the higher susceptibly of the target plants to momilactones. On the basis of IC50 values, monocotyledonous weed plant species (Echinochloa crus-galli, Echinochloa colonum, Phleum pretense, Digitaria sanguinalis and Lolium multiflorum) showed higher susceptibly compared to dicotyledonous plant species (Arabidopsis thaliana, Lepidium sativum, Lactuca sativa and Medicago sativa) [52,53,54,55]. In addition, momilactone B showed much higher growth-inhibitory activity than momilactone A, which has also been confirmed by other bioassay systems [55,56,57,58,59].
On the other hand, momilactones A and B showed relatively weak inhibitory activity on rice growth compared to Echinochloa crus-galli. The rice roots and shoots were suppressed by momilactones A and B at concentrations greater than 300 μM and 100 μM, respectively [52,53]. Thus, the effect of momilactones on rice was only 1% of that on Echinochloa crus-galli, which was inhibited at concentrations greater than 3 μM and 1 μM for roots and shoots, respectively, as described above [52,53]. In addition, momilactones A and B did not cause any visible damage to rice plants at concentrations that were phytotoxic to other plant species [52,53,54,55]. These observations suggest that the toxicity of momilactones A and B to rice plants is much less than that to other plant species. The resistance mechanism of rice to momilactones is unknown. This tolerance may possibly involve either rapid secretion, the insensitivity of the molecular target and/or the degradation of momilactones.
3.2. Concentration and Secretion of Momilactones
The endogenous concentrations of momilactones A and B, respectively, in rice were 4.5 μg/g and 3.0 μg/g of rice straw [60] and 4.9 μg/g and 2.9 μg/g of rice husks [61]. Momilactone B was found in rice seedlings 7 days after germination, and the concentrations of momilactones A and B increased until day 80 after germination, which is when flowering is initiated [52,62,63,64]. The 80-day-old rice plants contained momilactones A and B at 140 μg/g and 95 μg/g in rice plants, respectively [52,64]. Considering their reported concentrations, the ratio of momilactone A to momilactone B is 1.5–1.6.
The secretion of momilactone B from rice roots was observed 3 days after germination [62]. The levels of momilactone A and B secretion increased up to day 80 after germination and decreased thereafter [52,63]. The secretion levels of momilactones A and B at day 80 were 1.1 and 2.3 μg per plant per day, respectively [52,63], which indicates that the secretion ratio of momilactone B to momilactone A is 2.1. The observation suggests that rice secretes momilactones A and B into its rhizosphere throughout its entire life cycle, and the secretion increases until flowering initiation. Thus, it may be possible that rice allelopathy increases over this time frame. In addition, momilactone B was secreted at a higher rate than momilactone A, even though the concentration of momilactone A is higher than that of momilactone B in rice plants, which suggests that momilactone B may be preferentially secreted into the rhizosphere over momilactone A. Plants are reported to secrete a wide range of compounds from their roots through their cell membranes, for example, by proton-pumping mechanisms, plasmalemma-derived exudation and endoplasmic-derived exudation [65,66,67]. However, the mechanism of the exudation of momilactones from rice roots is unknown.
3.3. Contribution of Momilactones to Rice Allelopathy
When eight cultivars of rice seedlings (7 days old) were incubated for four days with Echinochloa crus-galli seedlings (4 days old) in a buffered bioassay medium, all rice cultivars suppressed the growth of Echinochloa crus-galli with different suppression levels. All rice cultivars produced and secreted momilactones A and B into the media, and the concentrations of momilactones A and B in the media were 0.21–1.45 μM and 0.66–3.84 μM, respectively [53]. Based on the growth-inhibitory activity and secreted amounts of momilactones A and B in the media, momilactone A may only account for 1.0–4.9% of the observed growth inhibition of Echinochloa crus-galli by the respective rice cultivars. By contrast, momilactone B may account for 58.8–81.9% of the observed growth inhibition. In addition, the momilactone B concentration in the media was significantly (p < 0.01) correlated with the extent of the growth suppression of Echinochloa crus-galli by these eight rice cultivars [53]. A similar correlation was also found between the level of momilactone B secretion and the extent of the growth suppression of Lactuca sativa by these rice cultivars [68,69]. The observations suggest that momilactone B may be a major contributor to the allelopathic activity of rice, and the secretion levels of momilactone B reflect the variation in allelopathic activity observed rice cultivars. The leaf, straw and husk extracts of 41 rice cultivars differed in their growth-inhibitory activity against Alisma plantago-aquatica. The concentration of momilactone B in the extracts was also correlated with the inhibitory activity of the extracts [70].
3.4. Genetic Evidence for Momilactones in Rice Allelopathy
Momilactone-biosynthesis-deficient mutants (cps4 and ksl4) were obtained through insertion gene knockouts for OsCPS4 and OsKSL4 [71,72], which is described in Section 5. Allelopathic activity after removing all syn-copalyl diphosphate-derived labdane-related diterpenoids (cps4 mutant) or, more selectively, only momilactones (ksl4 mutant) was compared to the respective wild-type rice. The wild types showed allelopathic activity, whereas both mutants lost this activity [73]. The investigation suggests that the loss of allelopathic activity may be attributed to the specific loss of momilactones, which verifies the involvement of momilactones in rice allelopathy.
3.5. Inhibitory Mechanism
Molecular targets of momilactone B were investigated through SDS-PAGE and two-dimensional gel electrophoresis with MALDI-TOF-MS. Momilactone B suppressed the germination of Arabidopsis thaliana and inhibited the breakdown of the storage proteins cruciferina, cruciferin 2 and cruciferin 3 during germination [74]. The breakdown of these proteins is essential to construct cell structures for germination and seedling growth [75,76,77]. The application of momilactone B to Arabidopsis thaliana seedlings inhibited the accumulation of amyrin synthase LUP2, subtilisin-like serine protease, β-glucosidase and malate synthase [78]. Those proteins are involved in the production of intermediates and metabolic turnover for cell structures [79,80,81,82]. On the contrary, momilactone B induced the accumulation of translationally controlled tumor protein, 1-cysteine peroxiredoxin 1 and glutathione-S-transferase [75]. These proteins elevate the tolerance to drought and oxidative stress conditions [83,84,85]. In addition, glutathione-S-transferase showed herbicide detoxification activity [86], and 1-cysteine peroxiredoxin 1 showed germination-inhibitory activity under unfavorable conditions [87]. These observations suggest that momilactone B may cause growth inhibition through the suppression of metabolic turnover and the production of intermediates and induce tolerance to stress conditions.
3.6. Induction of Rice Allelopathy and Momilactone
The allelopathic activity of rice was increased by nutrient deficiency, which is often caused by competition with neighboring plants [88,89,90]. The nutrient-deficient condition also increased the production and secretion of momilactone B from rice [91]. In addition, the allelopathic activity of rice was also elevated by either nearby Echinochloa crus-galli plants or their root exudates [91,92,93,94]. This elevation was not only owing to nutrient competition between rice and Echinochloa crus-galli [95,96]. The momilactone B concentration in rice and its secretion level from rice were also increased by either Echinochloa crus-galli or its root exudates. Rice may recognize certain components of the root exudation of Echinochloa crus-galli, and the compounds trigger the increased production and secretion of momilactone B [91,95,96]. Other weed species, namely, Eclipta prostate and Leptochola chinensis, also increased the secretion of momilactone B [97].
Rice allelopathic activity was also elevated by jasmonic acid [98]. The application of jasmonic acid and cantharidin with UV irradiation also increased the concentration of momilactone B in rice and the secretion levels of momilactones from rice roots into its rhizosphere [99]. As momilactones, especially momilactone B, have strong allelopathic activity, as described previously, such increasing secretion levels of momilactones may provide a competitive advantage for rice through the suppression of the growth of nearby competing plant species.
4. Pharmacological Activity
4.1. Anticancer Activity
Momilactones A and B showed growth suppression activity in the murine leukemia P399 cell line [100]. Momilactones A and B induced apoptosis in acute promyelocytic leukemia HL-60 and multiple myeloma U266 cell lines through the activation of apoptosis-inducing factors such as caspase-3 [101]. Momilactone B also induced G1 arrest in the cell cycle and apoptosis in the human leukemia U937 cell line through the suppression of pRB phosphorylation and the induction of the kinase inhibitor p21 [102], and it induced apoptosis in human leukemia T cells through the activation of caspase [103] and in human breast cancer cells through signal transducer and activator of transcription 5 and a caspase-3-dependent pathway [104]. Momilactone B showed cytotoxic activity in the human colon cancer HT-29 and SW620 cell lines [105].
4.2. Anti-Inflammatory Activity
Momilactone A suppressed the inflammatory response in mouse macrophage RAW264.7 cells through a reduction in NO production and iNOS mRNA expression [106].
4.3. Anti-Diabetic Activity
4.4. Anti-Ketosis Activity
Momilactone B inhibited ketosis in vitro through the suppression of the mitochondrial enzyme 3-hydroxy-3-methylglutaryl-CoA synthase-2, which converts acetyl-CoA to ketone bodies [109].
4.5. Anti-Melanogenic Activity
Momilactone B inhibited the accumulation of melanin in B16 melanocytes through the suppression of protein kinase A signaling and tyrosinase-related proteins [110].
5. Biosynthesis and Related Genes
Geranylgeranyl diphosphate (GGDP) is the precursor of the plant hormone gibberellin and rice diterpenoid phytoalexins such as oryzalexins and phytocassanes, including momilactones [29]. GGDP is synthesized by GGDP synthase (GGPS) from two five-carbon isoprenoids, isopentyl diphosphate or dimethylallyl diphosphate, which are synthesized through the methylerythritol phosphate pathway from pyruvate and glyceraldehyde-3-phosphate [111] (Figure 2).
GGDP is cyclized into syn-copalyl diphosphate (syn-CDP) by CDP synthases (OsCPS4). syn-CDP is further cyclized into syn-pimaradiene by ent-kaurene synthase-like 4 (OsKSL4) [112,113,114]. cDNA encoding OsCPS4 was obtained from UV-irradiated rice leaves [115]. OsCOS4 and OsKSL4 are located close to each other on chromosome 4 and were demonstrated to have sequential activity producing syn-CDP and syn-pimaradiene [116,117].
Cytochrome P450 enzymes (CYPs) are involved in the further metabolism of syn-pimaradiene. OsCYP99A3 oxidizes the C19 methyl of syn-pimaradiene into syn-pimaradien-19-oic acid [118,119], and OsCPY76M8 then hydroxylates its C6 position into 6β-hydroxy-syn-pimaradienon-19-oic acid, followed by the spontaneous closure of the ring between C19 and C6, which forms syn-pimaradienon-19,6β-hemiacetal [120]. Momilactone synthase (OsMS1 or OsMS2) converts the C19 hydroxyl group into a ketone to form syn-pimaradienon-19,6β-olide [120]. OsMS2 (or OsCPY701A8) then catalyzes C3 hydroxy into a ketone, forming momilactone A [120]. C20 hydroxylation of momilactone A by OsCP76M14 leads to the spontaneous closure of the hemiacetal ring and forms momilactone B [120,121]. The momilactone-synthesis-related genes OsCPS4, OsKSL4, CYP99A2, CYP99A3, OsMS1 and OsMS2 were reported to be located on chromosome 4 in plastids of rice cells [32,122] (Figure 3), which indicates that momilactones may play an important ecological role in rice evolution because of the presence of a dedicated biosynthetic gene cluster in the rice genome.
6. Momilactone Induction
Plants often respond by increasing their production of certain phytoalexins when they are attacked by pathogens and insects. The reaction involves the induction of active oxygen species, lignification, protease inhibitors and some enzymes, such as chitinase and β-glucanase. Plant defense reactions are also induced by a variety of biological, chemical and physical elicitors, such as oligosaccharides, cantharidin and UV irradiation [123,124,125]. Momilactone A and B production and accumulation were also induced by these elicitors.
6.1. Biotic Elicitors
Chitosan (oligosaccharide) is a deacetylated derivative of chitin, which is a long-chain polymer of N-acetylglucosamine and a primary component of fungal cell walls, arthropod exoskeletons and insect exuviae [126,127]. Chitosan increased the accumulation of momilactone A in rice leaves and suspension-cultured rice cells [6,7] and increased the tolerance of rice to the rice blight pathogen Fusarium oxysporum [128]. N-Acetylchitooligosaccharides, which are released from the cell walls of pathogenic fungi, also induced the accumulation of momilactones A and B in suspension-cultured rice cells, and their accumulation was 100–500 g/g of cultured cells, which is a sufficient concentration to prevent the growth of pathogenic fungi [129].
Tetraglucosyl glucitol [β-(1,3/1,6)-derived glucan] increased momilactone A production in rice cells [130]. Cantharidin, a protein serine/threonine phosphatase inhibitor contained in some insects, has been shown to mimic elicitor action in plants and to activate defense responses [131,132], and it increased the concentrations of momilactones A and B in rice [132,133] and the secretion level of momilactone B [134]. Cerebrosides (monoglycosylceramides), which are important components of animal cell membranes, induced β-glucanase, chitinase and peroxidase-encoding transcripts and enhanced the production of momilactone A [36,135]. The application of methionine also increased the momilactone A concentration in rice leaves. A free radical scavenger, Tiron (disodium 4,5-dihydroxy-1,3-benzenedisulfonate), increased the momilactone A concentration, which suggests that active oxygen species may stimulate methionine-induced momilactone A production [136].
6.2. Abiotic Elicitors
UV irradiation (254 nm, 20 min) increased momilactone A and B concentrations in rice leaves, and the maximum accumulation was found 3 days after UV irradiation [3,137]. The increase in the levels of momilactone A differed among rice varieties, and blast-resistant rice varieties accumulated more momilactone A than susceptible rice varieties [37].
The application of CuCl2 to rice leaves also induced momilactone A accumulation. The accumulation was detected 12 h after application and reached maximum accumulation at 72 h. FeCl2 and HgCl2 also increased momilactone A accumulation by 37% and 20% compared to CuCl2 application, respectively [138]. The application of CuCl2 to rice leaves induced jasmonic acid and momilactone A. Jasmonic acid biosynthesis inhibitors, quinacrine, nordihydroguaiaretic acid and salicylhydroxamic acid, suppressed momilactone A accumulation after the application of CuCl2. However, additional jasmonic acid application induced momilactone A accumulation after the application of CuCl2 and jasmonic acid biosynthesis inhibitors [139]. These observations suggest that CuCl2 increased the concentration of jasmonic acid in the leaves, and jasmonic acid then stimulated the biosynthesis of momilactone A. In addition, the application of CuCl2 and FeCl2 increased the production and secretion levels of momilactone B in rice and its allelopathic activity [133].
Other metal ions, such as silver, potassium, calcium, sodium zinc and magnesium, also increased the accumulation of momilactone A in suspension-cultured rice cells [140]. The air pollutant sulfur dioxide (SO2) induced reddish-brown necrotic spots on rice leaves and increased the momilactone A concentration in the leaves [141]. A fungicide, 2,2-dichloro-3,3-dimethyl cyclopropane carboxylic acid, also induced the accumulation of momilactones A and B in rice leaves [2]. Protein synthesis inhibitor herbicides, pretilachlor and butachlor, increased momilactone A accumulation in rice leaves [142].
7. Induction Signaling
The generation of elicitor fragments after pathogen and insect attacks may occur through the induction of chitinase and β-1,3-glucanase [143,144]. Elicitor fragments such as N-acetylchitooligosaccharide induced the formation of hetero-oligomer complexes of OsCEBiP (chitin elicitor binding protein) and OsCERK1 (chitin elicitor receptor kinase) [145]. OsCERK1 is part of the defensome complex at the plasma membrane (Figure 4). The defensome contains OsHsp70 (heat shock protein 70), OsHps90, OsHop/Sti1 (Hsp70/Hsp90 organizing protein/stress-induced protein 1), OsSGT1 (suppressor of G/two allele of Skp1) and OsRAR1 (required for Mla12 resistance) as molecular chaperone proteins and co-chaperon-like proteins [146,147,148]. OsRac1 (small-specific Rho-type GTPase), which is another important component, may cause mitogen-activated protein kinase (MAPK) signal cascades [149]. The earliest MAPK signaling step is OsACDR1 (accelerated cell death and resistance 1), followed by OsMKK4 and then OsMK3 and/or OsMK6 [150,151]. OsTGAP1 (TGA factor for phytoalexin production 1) may then induce the methylerythritol phosphate pathway and the expression of momilactone biosynthetic genes, including OsKSL4 [152,153].
OsRac1 may interact with OsRbohB (respiratory burst oxidase homolog B) in a Ca2+-dependent manner [154]. The constitutive expression of OsRac1 causes an increase in H2O2 production, OsCP2 transcripts and momilactone A accumulation in rice [155].
Tricoderma viride-derived xylanase (TvX) requires specific receptors [156] and increased cytosolic Ca2+ within minutes [157,158]. Cytosolic Ca2+ induction by TvX is partly mediated by the plasma membrane putative voltage-gated cation channel OsTPC1 [158]. TvX-induced signaling targets Ca2+-sensing calcineurin B-like proteins (OsCBL) and CBL-interacting protein kinases (CIPK14 and 15), which may act as Ca2+ sensors [157]. Increased momilactone production was found 24 h after TvX application [157,158].
Exogenous jasmonic acid (JA) induced the accumulation of momilactones [159]. Jasmonic acid production in plants after exposure to stress factors is initiated by the peroxidation of linolenic acid, followed by allene oxide cyclase-mediated epoxide formation, cyclization by allene oxide cyclase (OsAOC) and β-oxidation [159,160]. The produced jasmonic acid is then conjugated with isoleucine by OsJAR1 (Jasmonate Resistant 1), resulting in the formation of JA-isoleucine (JA-Il). JA-Il may then stimulate momilactone synthesis [159]. The exogenous application of salicylic acid also induced momilactone accumulation in rice. However, the mechanism of salicylic acid induction of momilactones remains unclear [161].
8. Occurrence of Momilactone
Momilactones A and B were first isolated from seed husks of Oryza sativa cv. Koshihikari [1] and then found in the leaves of Oryza sativa [2], but these studies did not clearly mention the cultivar or accession of the rice. Momilactones A and B were found in whole plants of rice, including their roots [60,64], and in multiple rice cultivars [53,68,69,70,162]. The concentrations of momilactones A and B were determined in the leaves of 69 rice cultivars from World Rice Core Collections, and in 64 and 31 cultivars, the presence of momilactones A and B were detected, respectively. The concentrations of momilactones A and B varied among these cultivars. The maximum amount of momilactone A was recorded in the cultivar Urasan at 495 nmol/g leaf, but the exact value for momilactone B was not reported. The concentrations of momilactones A and B in the leaves were greater in Japonica-type cultivars than in Indica-type cultivars [162].
Wild rice species such as Oryza rufipogon, O. burthii, O. glaberrima, O. glumaepatula, O. meridionalis, O. punctatas and O. brachyatha also contained momilactones A and B. The concentrations of momilactone A were 0.97–667 nmol/g leaf [162]. Momilactone biosynthesis genes of O. punctatas (OpCPS4, OpCYP99A, OpMS1, OpMS2, OpKSL4 and OpCYP99A) form a gene cluster on the same chromosome [163]. These genes are equivalent to rice OsCPS4, OsCYP99A, OsMS1, OsMS2, OsKSL4 and OsCYP99A genes, respectively. A gene cluster for momilactone biosynthesis was also found in Echinochloa crus-galli. The gene cluster contains only single copies of EcCY99A and EcMS, and its gene sequence on the chromosome is different from that in rice [164]. However, the endogenous concentration of momilactones in Echinochloa crus-galli has not yet been reported.
Momilactones A and B were also found in the moss species Calohypnum plumiforme (syn. Hypnum plumaeform), which is quite taxonomically distinct from rice [33,34,35]. Calohypnum plumiforme belongs to the Hypnaceae family of the Bryophyta division, often dominates in plant communities and forms large pure colonies in sunny places in lowland to upland areas, including marshy places in eastern Asia [165,166]. Momilactones are also synthesized from GGDP in the moss. GGDP is cyclized to syn-pimaradiene by diterpene cyclase (CpDTC1/HpDTC1) [167]. syn-Pimaradiene is catalyzed into 3β-hydroxy-syn-pimaradienon-19,6β-olide by CpCYP770A14 and CpCYP964A1. 3β-Hydroxy-syn-pimaradienon-19,6β-olide is then metabolized to momilactone A by momilactone synthase (CpMS) [168]. Those genes also form a gene cluster in the order CpMS, CpCYP970A14, CpDTC1/HpDTC1 and CpCYP964A1 on the same chromosome [168].
Momilactone A and B concentrations in the moss were 58.7 μg/g and 23.4 μg/g dry weight of the moss, respectively. The moss also secretes momilactones A and B into the rhizosphere at ratios of 4.0 μg/g and 6.3 μg/g dry weight of the moss, respectively, which were 7.3% and 27% of the endogenous concentrations of momilactones A and B in the moss [35]. The observations suggest that the moss selectively secretes momilactone B into the rhizosphere rather than momilactone A. UV irradiation, jasmonic acid and cantharidin also increased the production and secretion levels of momilactones A and B [169]. These observations suggest that elicitors and/or pathogen attacks may increase the production and secretion levels of momilactones A and B in the moss. Momilactones A and B secreted from moss are also able to suppress the growth of neighboring plant species. Therefore, momilactones in the moss may function in the defense against pathogen attacks and allelopathy.
9. Conclusions
The literature reviewed here demonstrates an important role for momilactones in the defense function and allelopathic function. Momilactones in rice plants may provide resistance to fungal pathogen attacks, and momilactones in rice root exudate may provide rice with the ability to compete with neighboring plant species, which was confirmed with momilactone-deficient mutants. The momilactone biosynthesis pathway and related genes have been investigated by many researchers. The elicitation of momilactone production and secretion and the endogenous signaling cascades involved in the elicitation are also well documented. These findings suggest that the allelopathic and defense functions of momilactones may play important ecological roles in rice evolution because of the existence of a dedicated biosynthetic gene cluster in the genome. However, the mechanism and molecular targets of momilactone functions remain unknown. Momilactones A and B did not cause growth suppression or any visible damage to rice plants at concentrations that were phytotoxic to other plant species. The resistance mechanism of rice to momilactones is also unknown. It is worth investigating the mechanism underlying this tolerance for developing resistant crop plants. The potential of momilactones to serve as endogenous natural fungicides and herbicides provides significant benefits when applied to other important crops. The identification of momilactones may provide a molecular marker for breeding and engineering directed at increasing defense and allelopathic abilities. In addition, momilactones have shown anti-leukemia and anti-diabetic activities. Further investigations are necessary to develop their medical applications.
Funding
This work was supported by JSPS Grants-in-Aid for Scientific Research, Grant Number JP21K05598.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kato, T.; Kabuto, C.; Sasaki, N.; Tsunagawa, M.; Aizawa, H.; Fujita, K.; Kato, Y.; Takahashi, N. Momilactones, growth inhibitors from rice, Oryza sativa L. Tetrahedron Lett. 1973, 39, 3861–3864. [Google Scholar] [CrossRef]
- Cartwright, D.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Chemical activation of host defense mechanisms as a basis for crop protection. Nature 1977, 267, 511–513. [Google Scholar] [CrossRef]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Isolation and characterization of two phytoalexins from rice as momilactones A and B. Phytochemistry 1981, 20, 535–537. [Google Scholar] [CrossRef]
- Takahashi, A.; Kawasaki, T.; Henmi, K.; Shii, K.; Kodama, O.; Satoh, H.; Shimamoto, K. Lesion mimic mutants of rice with alterations in early signaling events of defense. Plant J. 1999, 17, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Imai, T.; Koga, J.; Okada, K.; Yamane, H.; Ohashi, Y. Phytoalexin accumulation in the interaction between rice and the blast fungus. Mol. Plant-Micro. Intrac. 2010, 23, 1000–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, G.K.; Rakwal, R.; Tamogami, S.; Yonekura, M.; Kubo, A.; Saji, H. Chitosan activates defense/stress response(s) in the leaves of Oryza sativa seedlings. Plant Physiol. Biochem. 2002, 40, 1061–1069. [Google Scholar] [CrossRef]
- Shimizu, T.; Jikumaru, Y.; Okada, A.; Okada, K.; Koga, J.; Umemura, K.; Minami, E.; Shibuya, N.; Hasegawa, M.; Kodama, O.; et al. Effects of a bile acid elicitor, cholic acid, on the biosynthesis of diterpenoid phytoalexins in suspension-cultured rice cells. Phytochemistry 2008, 69, 973–981. [Google Scholar] [CrossRef]
- Tamogami, S.; Kodama, O. Coronatine elicits phytoalexin production in rice leaves (Oryza sativa L.) in the same manner as jasmonic acid. Phytochemistry 2000, 54, 689–694. [Google Scholar]
- Jung, Y.H.; Lee, J.H.; Agrawal, G.K.; Rakwal, R.; Kim, J.A.; Shim, J.K.; Lee, S.K.; Jeon, J.S.; Koh, H.J.; Lee, Y.H.; et al. The rice (Oryza sativa) blast lesion mimic mutant, blm, may confer resistance to blast pathogens by triggering multiple defense-associated signaling pathways. Plant Physiol. Biochem. 2005, 43, 397–406. [Google Scholar] [CrossRef]
- Okada, A.; Shimizu, T.; Okada, K.; Kuzuyama, T.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. Elicitor induced activation of the methylerythritol phosphate pathway toward phytoalexins biosynthesis in rice. Plant Mol. Biol. 2007, 65, 177–187. [Google Scholar] [CrossRef]
- Dilday, R.H.; Nastasi, P.; Smith, R.J., Jr. Allelopathic observations in rice (Oryza sativa L.) to ducksalad (Heteranthera limosa). Proc. Arkansas. Acad. Sci. 1989, 43, 21–22. [Google Scholar]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Putnam, A.R.; Tang, C.S. Allelopathy: State of the science. In The Science of Allelopathy; Putnam, A.R., Tang, C.S., Eds.; John Wiley and Sons: Ithaca, NY, USA, 1986; pp. 1–19. [Google Scholar]
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef] [PubMed]
- Dilday, R.H.; Lin, J.; Yan, W. Identification of allelopathy in the USDA-ARS rice germplasm collection. Aust. J. Exp. Agric. 1994, 34, 907–910. [Google Scholar] [CrossRef]
- Dilday, R.H.; Yan, W.G.; Moldenhauer, K.A.K.; Gravois, K.A. Allelopathic activity in rice for controlling major aquatic weeds. In Allelopathy in Rice; Olofsdotter, M., Ed.; International Rice Research Institute: Manila, Philippines, 1998; pp. 7–26. [Google Scholar]
- Hassan, S.M.; Aidy, I.R.; Bastawisi, A.O.; Draz, A.E. Weed management using allelopathic rice varieties in Egypt. In Allelopathy in Rice; Olofsdotter, M., Ed.; International Rice Research Institute: Manila, Philippines, 1998; pp. 27–37. [Google Scholar]
- Kim, K.U.; Shin, D.H. Rice allelopathy research in Korea. In Allelopathy in Rice; Olofsdotter, M., Ed.; International Rice Research Institute: Manila, Philippines, 1998; pp. 39–43. [Google Scholar]
- Olofsdotter, M.; Navarez, D.; Rebulanan, M.; Streibig, J.C. Weed-suppressing rice cultivars: Does allelopathy play a role? Weed Res. 1999, 39, 441–454. [Google Scholar] [CrossRef]
- Pheng, S.; Adkins, S.; Olofsdotter, M.; Jahn, G. Allelopathic effects of rice (Oryza sativa L.) on the growth of awnless barnyardgrass (Echinochloa colona (L.) Link): A new form for weed management. Cambodian J. Agri. 1999, 2, 42–49. [Google Scholar]
- Kato-Noguchi, H.; Ino, T. Assessment of allelopathic potential of root exudate of rice seedlings. Biol. Plant. 2001, 44, 635–638. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T.; Sata, N.; Yamamura, S. Isolation and identification of a potent allelopathic substance in rice root exudates. Physiol. Plant. 2002, 115, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Ino, T.; Ota, K. Secretion of momilactone A from rice roots to the rhizosphere. J. Plant Physiol. 2008, 165, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Allelopathic substance in rice root exudates: Rediscovery of momilactone B as an allelochemical. J. Plant Physiol. 2004, 161, 271–276. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Peters, R.J. The role of momilactones in rice allelopathy. J. Chem. Ecol. 2013, 39, 175–185. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Huffaker, A.; Sims, J.W.; Christensen, S.A.; Lu, X.; Okada, K.; Peters, R.J. Biosynthesis, elicitation and roles of monocot terpenoid phytoalexins. Plant J. 2014, 79, 659–678. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H. Convergent or parallel molecular evolution of momilactone A and B: Potent allelochemicals, momilactones have been found only in rice and the moss Hypnum plumaeforme. J. Plant Physiol. 2011, 168, 1511–1516. [Google Scholar] [CrossRef]
- Serra, N.S.; Shanmuganathan, R.; Becker, C. Allelopathy in rice: A story of momilactones, kin recognition, and weed management. J. Exp. Bot. 2021, 72, 4022–4037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Peters, R.J. Why are momilactones always associated with biosynthetic gene clusters in plants? Proc. Natl. Acad. Sci. USA 2020, 117, 13867–13869. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Shigemori, H.; Kato-Noguch, H. Allelopathic potential of Hypnum plumaeforme L. and its allelopathic substances. In Proceedings of the 4th Asia-Pacific Conference on Chemical Ecology, from Biomolecules to Ecosystems an Interactive Chemical Message for our Future, Tsukuba, Japan, 10–14 September 2007; p. 77. [Google Scholar]
- Nozaki, H.; Hayashi, K.I.; Nishimura, N.; Kawaide, H.; Matsuo, A.; Takaoka, D. Momilactone A and B as allelochemicals from moss Hypnum plumaeforme: First occurrence in bryophytes. Biosci. Biotech. Biochem. 2007, 71, 3127–3130. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kobayashi, K.; Shigemori, H. Allelopathy of the moss Hypnum plumaeforme by the production of momilactone A and B. Weed Res. 2009, 49, 621–627. [Google Scholar] [CrossRef]
- Umemura, K.; Ogawa, N.; Shimura, M.; Koga, J.; Usami, H.; Kono, T. Possible role of phytocassane, rice phytoalexin, in disease resistance of rice against the blast fungus Magnaporthe grisea. Biosci. Biotech. Biochem. 2003, 67, 899–902. [Google Scholar] [CrossRef] [Green Version]
- Dillon, V.M.; Overton, J.; Grayer, R.J.; Harborne, J.B. Differences in phytoalexin response among rice cultivars of different resistance to blast. Phytochemistry 1997, 44, 599–603. [Google Scholar] [CrossRef]
- Toyomasu, T.; Usui, M.; Sugawara, C.; Otomo, K.; Hirose, Y.; Miyao, A.; Hirochik, H.; Okad, K.; Shimizu, T.; Koga, J.; et al. Reverse-genetic approach to verify physiological roles of rice phytoalexins: Characterization of a knockdown mutant of OsCPS4 phytoalexin biosynthetic gene in rice. Physiol. Plant. 2014, 150, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, M.; Xuan, T.D.; Deba, F.; Tawata, S.; Dang Khanh, T.; Chung, M.I. Comparative efficacies in vitro of antibacterial, fungicidal, antioxidant, and herbicidal activities of momilatones A and B. J. Plant Interac. 2007, 2, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.Z.; Xia, X.M.; Lv, J.; Tan, J.W.; Baerson, S.R.; Pan, Z.Q.; Song, Y.Y.; Zeng, R.S. Diterpenoids with herbicidal and antifungal activities from hulls of rice (Oryza sativa). Fitoterapia 2019, 136, 104183. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, X.; Xiao, J.; Wang, S. A convenient method for simultaneous quantification of multiple phytohormones and metabolites: Application in study of rice-bacterium interaction. Plant Methods 2012, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Howe, G.A.; Major, I.T.; Koo, A.J. Modularity in jasmonate signaling for multistress resilience. Ann. Rev. Plant Biol. 2018, 69, 387–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Plant responses to herbivory, wounding, and infection. Int. J. Mol. Sci. 2022, 23, 7031. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Goossens, A.; Lacchini, E. Jasmonate: A hormone of primary importance for plant metabolism. Curr. Opin. Plant Biol. 2022, 67, 102197. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Ishida, S.; Saito, C.; Maita, T.; Kusano, M.; Tamogami, S.; Noma, M. Stimulation of mycelia growth in several mushroom species by rice husks. Biosci. Biotech. Biochem. 2005, 69, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.M.; Ali, M.; Ahmad, A.; Chun, S.C.; Kim, J.T.; Sultana, S.; Kim, J.S.; Seo, B.R. Steroidal constituents of rice (Oryza sativa) hulls with Algicidal and Herbicidal activity against blue-green algae and duckweed. Phytochem. Anal. 2007, 18, 133–145. [Google Scholar] [CrossRef]
- Kanno, H.; Hasegawa, M.; Kodama, O. Accumulation of salicylic acid, jasmonic acid and phytoalexins in rice, Oryza sativa, infested by the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). Appl. Entomol. Zool. 2012, 47, 27–34. [Google Scholar] [CrossRef]
- Wari, D.; Alamgir, K.M.; Mujiono, K.; Hojo, Y.; Tani, A.; Shinya, T.; Nakatania, H.; Galis, I. Brown planthopper honeydew-associated symbiotic microbes elicit momilactones in rice. Plant Signal. Behav. 2019, 14, 1655335. [Google Scholar] [CrossRef] [PubMed]
- Azmi, M.; Abdullah, M.Z.; Fujii, Y. Exploratory study on allelopathic effect of selected Malaysian rice varieties and rice field weed species. J. Trop. Agric. Food Sci. 2000, 28, 39–54. [Google Scholar]
- Rao, A.N.; Johnson, D.E.; Sivaprasad, B.; Ladaha, J.K.; Mortimer, A.M. Weed management in direct-seeded rice. Adv. Agron. 2007, 93, 153–255. [Google Scholar]
- Kong, C.H. Rice allelopathy. Allelopathy J. 2008, 22, 261–278. [Google Scholar]
- Kato-Noguchi, H.; Ota, K.; Ino, T. Release of momilactone A and B from rice plants into the rhizosphere and its bioactivities. Allelopathy J. 2008, 22, 321–328. [Google Scholar]
- Kato-Noguchi, H.; Hasegawa, M.; Ino, T.; Ota, K.; Kujime, H. Contribution of momilactone A and B to rice allelopathy. J. Plant Physiol. 2010, 167, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Ota, K.; Kujime, H. Absorption of momilactone A and B by Arabidopsis thaliana L. and the growth inhibitory effects. J. Plant Physiol. 2012, 169, 1471–1476. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ota, K. Biological activities of rice allelochemicals momilactone A and B. Rice Res. 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Kato, T.; Tsunagawa, M.; Sasaki, N.; Kitahara, Y. Mechanisms of dormancy in rice seeds. II. New growth inhibitors, momilactone-A and -B isolated from the hulls of rice seeds. Jpn. J. Breed. 1976, 26, 91–98. [Google Scholar] [CrossRef]
- Kato, T.; Tsunakawa, M.; Sasaki, N.; Aizawa, H.; Fujita, K.; Kitahara, Y.; Takahashi, N. Growth and germination inhibitors in rice husks. Phytochemistry 1977, 16, 45–48. [Google Scholar] [CrossRef]
- Chung, I.M.; Hahh, S.J.; Ahmad, A. Confirmation of potential herbicidal agents in hulls of rice, Oryza sativa. J. Chem. Ecol. 2005, 31, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Toyomasu, T.; Kagahara, T.; Okada, K.; Koga, J.; Hasegawa, M.; Mitsuhashi, W.; Sassa, T.; Yamane, H. Diterpene phytoalexins are biosynthesized in and exuded from the roots of rice seedlings. Biosci. Biotechnol. Biochem. 2008, 72, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Yoneyama, K.; Takeuchi, Y.; Konnai, M.; Tamogami, S.; Kodama, O. Momilactones A and B in rice straw harvested at different growth stages. Biosci. Biotechnol. Biochem. 1999, 63, 1318–1320. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.-M.; Kim, T.K.; Kim, S.H. Evaluation of allelopathic potential and quantification of momilactone A, B from rice hull extracts and assessment of inhibitory bioactivity on paddy field weeds. J. Agric. Food. Chem. 2006, 54, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Ino, T. Rice seedlings release momilactone B into the environment. Phytochemistry 2003, 63, 551–554. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T.; Ichii, M. Changes in release level of momilactone B into the environment from rice throughout its life cycle. Func. Plant Biol. 2003, 30, 995–997. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T. Possible involvement of momilactone B in rice allelopathy. J. Plant Physiol. 2005, 162, 718–721. [Google Scholar] [CrossRef]
- Hawes, M.C.; Gunawardena, U.; Miyasaka, S.; Zhao, X. The role of root border cells in plant defense. Trends Plant Sci. 2000, 5, 128–133. [Google Scholar] [CrossRef]
- Bais, H.P.; Park, S.W.; Weir, T.L.; Callaway, R.M.; Vivanco, J.M. How plants communicate using the underground information superhighway. Trends Plant Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Ino, T. Concentration and release level of momilactone B in the seedlings of eight rice cultivars. J. Plant Physiol. 2005, 162, 965–969. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T.; Kujime, H. The relation between growth inhibition and secretion level of momilactone B from rice root. J. Plant Interact. 2010, 5, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Mennan, H.; Ngouajio, M.; Sahin, M.; Isik, D.; Altop, E.K. Quantification of momilactone B in rice hulls and the phytotoxic potential of rice extracts on the seed germination of Alisma plantago-aquatica. Weed Biol. Manag. 2012, 12, 29–39. [Google Scholar] [CrossRef]
- Zhang, J.; Li, C.; Wu, C.; Xiong, L.; Chen, G.; Zhang, Q.; Wang, S. RMD: A rice mutant database for functional analysis of the rice genome. Nucleic Acids Res. 2006, 34, D745–D748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, J.S.; Lee, S.; Jung, K.H.; Jun, S.H.; Jeong, D.H.; Lee, J.; Kim, C.; Jang, S.; Yang, K.; Nam, J.; et al. T-DNA insertional mutagenesis for functional genomics in rice. Plant J. 2000, 22, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Galhano, R.; Wiemann, P.; Bueno, E.; Tiernan, M.; Wu, W.; Chung, I.M.; Gershenzon, J.; Sesma, A.T.; Peters, R.J. Genetic evidence for natural product-mediated plant-plant allelopathy in rice (Oryza sativa). New Phytol. 2012, 193, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H.; Ota, K.; Kujime, H.; Ogawa, M. Effects of momilactone on the protein expression in Arabidopsis germination: Arabidopsis and momilactone. Weed Biol. Manag. 2013, 13, 19–23. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Tenbarge, K.M.; Shumway, J.E.; Crouch, M.L. Role of ABA in maturation of rapeseed embryos. Plant Physiol. 1985, 78, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed; Springer: New York, NY, USA, 2012; pp. 1–408. [Google Scholar]
- Job, C.; Rajjou, L.; Lovigny, Y.; Belghazi, M.; Job, D. Patterns of protein oxidation in Arabidopsis seeds and during germination. Plant Physiol. 2005, 138, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H.; Kitajima, S. Momilactone sensitive proteins in Arabidopsis thaliana. Nat. Prod. Commun. 2015, 10, 729–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohyama, K.; Suzuki, M.; Kikuchi, J.; Saito, K.; Muranaka, T. Dual biosynthetic pathways to phytosterol via cycloartenol and lanosterol in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Kuroha, T.; Okuda, A.; Arai, M.; Komatsu, Y.; Sato, S.; Kato, T.; Tabata, S.; Satoh, S. Identification of Arabidopsis subtilisin-like serine protease specifically expressed in root stele by gene trapping. Physiol. Plant. 2009, 137, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, K.; Job, C.; Groot, S.P.C.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomics of Arabidopsis seed germination. A comparative study of wild-type and gibberellin-deficient seeds. Plant Physiol. 2002, 129, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Eastmond, P.J.; Graham, I.A. Re-examining the role of the glyoxylate cycle in oilseeds. Trends Plant Sci. 2001, 6, 72–77. [Google Scholar] [CrossRef]
- Stacy, R.A.P.; Nordeng, T.W.; Culianez-Macia, F.A.; Reidunn, B.; Aalen, R.B. The dormancy-related peroxiredoxin anti-oxidant, PER1, is localized to the nucleus of barley embryo and aleurone cells. Plant J. 1999, 19, 1–8. [Google Scholar] [CrossRef]
- Kim, S.Y.; Paeng, S.K.; Nawkar, G.M.; Maibam, P.; Lee, E.S.; Kim, K.S.; Lee, D.H.; Park, D.J.; Kang, S.B.; Kim, M.R.; et al. The 1-cys peroxiredoxin, a regulator of seed dormancy, functions as a molecular chaperone under oxidative stress conditions. Plant Sci. 2011, 181, 119–124. [Google Scholar] [CrossRef]
- Fanucchi, F.; Alpi, E.; Olivieri, S.; Cannistraci, C.V.; Bachi, A.; Alpi, A.; Alessio, M. Acclimation increases freezing stress response of Arabidopsis thaliana at proteome level. Biochim. Biophys. Acta 2012, 1824, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Neuefeind, T.; Reinemer, P.; Bieseler, B. Plant glutathione S-transferases and herbicide detoxification (Review). Biol. Chem. 1997, 378, 199–205. [Google Scholar] [PubMed]
- Haslekås, C.; Viken, M.K.; Grini, P.E.; Nygaard, V.; Nordgard, S.H.; Meza, T.J.; Aalen, R.B. Seed 1-cysteine peroxiredoxin antioxidants are not involved in dormancy, but contribute to inhibition of germination during stress. Plant Physiol. 2003, 133, 1148–1157. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.U.; Shin, D.H.; Lee, I.J.; Kim, H.Y.; Kim, K.U.; Shin, D.H. Rice allelopathy in Korea. In Rice Allelopathy; Kim, K.U., Shin, D.H., Eds.; Kyungpook National University: Taegu, Korea, 2000; pp. 57–82. [Google Scholar]
- Song, B.; Xiong, J.; Fang, C.; Qiu, L.; Lin, R.; Liang, Y.; Lin, W. Allelopathic enhancement and differential gene expression in rice under low nitrogen treatment. J. Chem. Ecol. 2008, 34, 688–695. [Google Scholar] [CrossRef]
- Shen, L.; Lin, W. Effects of phosphorus levels on allelopathic potential of rice co-cultured with barnyardgrass. Allelopathy J. 2007, 19, 393–402. [Google Scholar]
- Kato-Noguchi, H. Barnyard grass-induced rice allelopathy and momilactone B. J. Plant Physiol. 2011, 168, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, H.; Kong, C.; Xu, X.; Liang, W. Chemical response of allelopathic rice seedlings under varying environmental conditions. Allelopathy J. 2005, 15, 105–110. [Google Scholar]
- Kong, C.H.; Li, H.B.; Hu, F.; Xu, X.H.; Wang, P. Allelochemicals released by rice roots and residues in soil. Plant Soil 2006, 288, 47–56. [Google Scholar] [CrossRef]
- Li, L.L.; Zhao, H.H.; Kong, C.H. (–)-Loliolide, the most ubiquitous lactone, is involved in barnyardgrass-induced rice allelopathy. J. Exp. Bot. 2020, 71, 1540–1550. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. The chemical cross talk between rice and barnyardgrass. Plant Signal. Behav. 2011, 6, 1207–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato-Noguchi, H.; Ino, T. The chemical-mediated allelopathic interaction between rice and barnyard grass. Plant Soil 2013, 370, 267–275. [Google Scholar] [CrossRef]
- Yang, X.F.; Kong, C.H. Interference of allelopathic rice with paddy weeds at the root level. Plant Biol. 2017, 19, 584–591. [Google Scholar] [CrossRef]
- Bi, H.H.; Zeng, R.Z.; Su, L.M.; An, M.; Luo, S.H. Rice allelopathy induced by methyl jasmonate and methyl salicylate. J. Chem. Ecol. 2007, 33, 1089–1103. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kujime, H.; Ino, T. UV-induced momilactone B accumulation in rice rhizosphere. J. Plant Physiol. 2007, 164, 1548–1551. [Google Scholar] [CrossRef]
- Chung, I.M.; Ali, M.; Hahn, S.J.; Siddiqui, N.A.; Lim, Y.H.; Ahmad, A. Chemical constituents from the hulls of Oryza sativa with cytotoxic activity. Chem. Nat. Compd. 2005, 41, 182–189. [Google Scholar] [CrossRef]
- Anh, L.H.; Lam, V.Q.; Takami, A.; Khanh, T.D.; Quan, N.V.; Xuan, T.D. Cytotoxic mechanism of momilactones A and B against acute promyelocytic leukemia and multiple myeloma cell Lines. Cancers 2022, 14, 4848. [Google Scholar] [CrossRef]
- Park, C.; Jeong, N.Y.; Kim, G.Y.; Han, M.H.; Chung, I.M.; Kim, W.J.; Yoo, Y.Y.; Choi, Y.H. Momilactone B induces apoptosis and G1 arrest of the cell cycle in human monocytic leukemia U937 cells through downregulation of pRB phosphorylation and induction of the cyclin-dependent kinase inhibitor p21Waf1/Cip1. Oncol. Rep. 2014, 31, 1653–1660. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Chung, I.M.; Jin, Y.J.; Song, Y.S.; Seo, S.Y.; Park, B.S.; Vho, K.H.; Yoo, K.S.; Yee, S.B.; Yoo, Y.H. Momilactone B, an allelochemical of rice hulls, induces apoptosis on human lymphoma cells (Jurkat) in a micromolar concentration. Nutr. Cancer 2008, 60, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Joung, Y.H.; Lim, E.J.; Kim, M.S.; Lim, S.D.; Yoon, S.Y.; Lim, Y.C.; Yoo, Y.B.; Ye, S.K.; Park, T.; Chung, I.M.; et al. Enhancement of hypoxia-induced apoptosis of human breast cancer cells via STAT5b by momilactone B. Int. J. Oncol. 2008, 33, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Park, H.R.; Park, E.; Lee, S.C. Cytotoxic and antitumor activity of momilactone B from rice hulls. J. Agric. Food Chem. 2007, 55, 1702–1706. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.G.; Cha, B.J.; Lee, S.M.; Shrestha, S.; Jeong, R.H.; Lee, D.S.; Kim, Y.C.; Lee, D.G.; Kang, H.C.; Jiyoung Kima, J.; et al. Diterpenes from the roots of Oryza sativa L. and their inhibition activity on NO production in LPS-stimulated RAW264. 7 macrophages. Chem. Biodivers. 2015, 12, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Quan, N.V.; Tran, H.D.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef] [Green Version]
- Quan, N.; Xuan, T.D.; Tran, H.D.; Ahmad, A.; Khanh, T.D.; Dat, T.D. Contribution of momilactones A and B to diabetes inhibitory potential of rice bran: Evidence from in vitro assays. Saudi Pharm. J. 2019, 27, 643–649. [Google Scholar] [CrossRef]
- Kang, D.Y.; SP, N.; Darvin, P.; Joung, Y.H.; Byun, H.J.; Do, C.H.; Park, K.D.; Cho, K.H.; Yang, Y.M. Momilactone B inhibits ketosis in vitro by regulating the ANGPTL3-LPL pathway and inhibiting HMGCS2. Anim. Biotechnol. 2017, 28, 189–197. [Google Scholar] [CrossRef]
- Lee, J.H.; Cho, B.; Jun, H.J.; Seo, W.D.; Kim, D.W.; Cho, K.J.; Lee, S.J. Momilactione B inhibits protein kinase A signaling and reduces tyrosinase-related proteins 1 and 2 expression in melanocytes. Biotechnol. Let. 2012, 4, 805–812. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Network analysis of the VA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 4, 665–700. [Google Scholar] [CrossRef]
- Nemoto, T.; Cho, E.M.; Okada, A.; Okada, K.; Otomo, K.; Kanno, Y.; Toyomasu, T.; Mitsuhashi, W.; Sassa, T.; Minami, E.; et al. Stemar-13-ene synthase, a diterpene cyclase involved in the biosynthesis of the phytoalexin oryzalexin S in rice. FEBS Lett. 2004, 571, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Otomo, K.; Kanno, Y.; Motegi, A.; Kenmoku, H.; Yamane, H.; Mitsuhashi, W.; Okikawa, H.; Toshima, H.; Itoh, H.; Matsuoka, M.; et al. Diterpene cyclases responsible or the biosynthesis of phytoalexins, momilactones A, B, and oryzalexins A-F in rice. Biosci. Biotechnol. Biochem. 2004, 68, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Hillwig, M.L.; Prisic, S.; Coates, R.M.; Peters, R.J. Functional identification of rice syn-copalyl diphosphate synthase and its role in initiating biosynthesis of diterpenoid phytoalexin/allelopathic natural products. Plant J. 2004, 39, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Otomo, K.; Kenmoku, H.; Oikawa, H.; Konig, W.A.; Toshima, H.; Mitsuhashi, W.; Yamane, H.; Sassa, T.; Toyomasu, T. Biological functions of ent- and syn-copalyl diphosphate synthases in rice: Key enzymes for the branch point of gibberellin and phytoalexin biosynthesis. Plant J. 2004, 39, 886–893. [Google Scholar] [CrossRef]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol. 2004, 134, 1642–1653. [Google Scholar] [CrossRef] [Green Version]
- Wilderman, P.R.; Xu, M.; Jin, Y.; Coates, R.M.; Peters, R.J. Identification of syn-pimara-7,15-diene synthase reveals functional clustering of terpene synthases involved in rice phytoalexin/allelochemical biosynthesis. Plant Physiol. 2004, 135, 2098–2105. [Google Scholar] [CrossRef] [Green Version]
- Shimura, K.; Okada, A.; Okada, K.; Jikumaru, Y.; Ko, K.W.; Toyomasu, T.; Sassa, T.; Hasegawa, M.; Kodama, O.; Shibuya, N.; et al. Identification of a biosynthetic gene cluster in rice for momilactones. J. Biol. Chem. 2007, 282, 34013–34018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Hillwig, M.L.; Peters, R.J. CYP99A3: Functional identification of a diterpene oxidase from the momilactone biosynthetic gene cluster in rice. Plant J. 2011, 65, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Kitaoka, N.; Zhang, J.; Oyagbenro, R.K.; Brown, B.; Wu, Y.; Yang, B.; Li, Z.; Peters, R.J. Interdependent evolution of biosynthetic gene clusters for momilactone production in rice. Plant Cell 2021, 33, 290–305. [Google Scholar] [CrossRef] [PubMed]
- De La Peña, R.; Sattely, E.S. Rerouting plant terpene biosynthesis enables momilactone pathway elucidation. Nat. Chem. Biol. 2020, 17, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, J.; Li, Z.; Peters, R.J.; Yang, B. Dissecting the labdane-related diterpenoid biosynthetic gene clusters in rice reveals directional cross-cluster phytotoxicity. New Phytologist. 2022, 233, 878–889. [Google Scholar] [CrossRef]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced resistance for plant disease control: Maximizing the effcacy of resistance elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langner, T.; Kamoun, S.; Belhaj, K. CRISPR crops: Plant genome editing toward disease resistance. Annu. Rev. Phytopathol. 2018, 56, 479–512. [Google Scholar] [CrossRef] [Green Version]
- Van, D.E.; Koornneef, A.; Ton, J.; Pieterse, C.M. Induced resistance-orchestrating defence mechanisms through crosstalk and priming. Annu. Plant Rev. 2018, 34, 334–370. [Google Scholar]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [Green Version]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Ma, B.; Wang, J.; Liu, C.; Hu, J.; Tan, K.; Zhao, F.; Yuan, M.; Zhang, J.; Gai, Z. Preventive effects of fluoro-substituted benzothiadiazole derivatives and chitosan oligosaccharide against the rice seedling blight induced by Fusarium oxysporum. Plants 2019, 8, 538. [Google Scholar] [CrossRef] [Green Version]
- Yamada, A.; Shibuya, N.; Kodama, O.; Akatsuka, T. Induction of phytoalexin formation in suspension-cultured rice cells by N-acetylchitooligosaccharides. Biosci. Biotech. Biochem. 1993, 57, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Yamada, A.; Hong, N.; Ogawa, T.; Ishii, T.; Shibuya, N. Differences in the recognition of glucan elicitor signals between rice and soybean: B-glucan fragments from the rice blast disease fungus Pyricularia oryzae that elicit phytoalexin biosynthesis in suspension-cultured rice cells. Plant Cell 2000, 12, 817–826. [Google Scholar] [PubMed] [Green Version]
- MacKintosh, C.; Lyon, G.D.; MacKintosh, R.W. Protein phosphatase inhibitors activate anti-fungal defense responses of soybean cotyledons and cell cultures. Plant J. 1994, 5, 137–147. [Google Scholar] [CrossRef]
- Rakwal, R.; Shii, K.; Agrawal, G.K.; Yonekura, M. Protein phosphatase inhibitors activate defense responses in rice (Oryza sativa) leaves. Physiol. Plant. 2001, 111, 151–157. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kobayashi, K. Jasmonic acid, protein phosphatase inhibitor, metals and UV-irradiation increased momilactone A and B concentrations in the moss Hypnum plumaeforme. J. Plant Physiol. 2009, 166, 1118–1122. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Stress-induced allelopathic activity and momilactone B in rice. Plant Growth Regul. 2009, 59, 153–158. [Google Scholar] [CrossRef]
- Koga, J.; Yamauchi, T.; Shimura, M.; Ogawa, N.; Oshima, K.; Umemura, K.; Kikuchi, M.; Ogasawara, N. Cerebrosides A and C, sphingolipid elicitors of hypersensitive cell death and phytoalexin accumulation in rice plants. J. Biol. Chem. 1998, 273, 31985–31991. [Google Scholar] [CrossRef] [Green Version]
- Nakazato, Y.; Tamogami, S.; Kawai, H.; Hasegawa, M.; Kodama, O. Methionine-induced phytoalexin production in rice leaves. Biosci. Biotech. Biochem. 2000, 64, 577–583. [Google Scholar] [CrossRef]
- Kodama, O.; Suzuki, T.; Miyakawa, J.; Akatsuka, T. Ultraviolet-induced accumulation of phytoalexins in rice leaves. Agric. Biol. Chem. 1988, 52, 2469–2473. [Google Scholar]
- Kodama, O.; Yamada, A.; Yamamoto, A.; Takemoto, T.; Akatsuka, T. Induction of phytoalexins with heavy metal ions in rice leaves. J. Pesticide Sci. 1988, 13, 615–617. [Google Scholar] [CrossRef] [Green Version]
- Rakwal, R.; Tamogami, S.; Kodama, O. Role of jasmonic acid as a signaling molecule in copper chloride-elicited rice phytoalexin production. Biosci. Biotech. Biochem. 1996, 60, 1046–1048. [Google Scholar] [CrossRef]
- Qin, J.; Wand, Y.; He, G. Induction of phytoalexins (PA) formation in suspension-cultured rice cells by metal and nonmetal Ions. Chin. J. Appl. Environ. Biol. 2006, 12, 322. [Google Scholar]
- Rakwal, R.; Agrawal, G.K.; Kubo, A.; Yonekura, M.; Tamogami, S.; Saji, H.; Iwahashi, H. Defense/stress responses elicited in rice seedlings exposed to the gaseous air pollutant sulfur dioxide. Environ. Exp. Bot. 2003, 49, 223–235. [Google Scholar] [CrossRef]
- Tamogami, S.; Kodama, O.; Hirose, K.; Akatsuka, T. Pretilachlor [2-chloro-N-(2, 6-diethylphenyl)-N-(2-propoxyethyl) acetamide]-and butachlor [N-(butoxymethyl)-2-chloro-N-(2, 6-diethylphenyl) acetamide]-induced accumulation of phytoalexin in rice (Oryza sativa) plants. J. Agric. Food Chem. 1995, 43, 1695–1697. [Google Scholar] [CrossRef]
- Ji, C.; Norton, R.A.; Wicklow, D.T.; Dowd, P.F. Isoform patterns of chitinase and b-1,3-glucanase in maturing corn kernels (Zea mays L.) associated with Aspergillus flavus milk stage infection. J. Agric. Food Chem. 2000, 48, 507–511. [Google Scholar] [CrossRef]
- Rodriguez, V.M.; Santiago, R.; Malvar, R.A.; Butron, A. Inducible maize defense mechanisms against the corn borer Sesamia nonagrioides: A transcriptome and biochemical approach. Mol. Plant-Microbe Interact. 2012, 25, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Nakano, T.; Takamizawa, D.; Desaki, Y.; Ishii-Minami, N.; Nishizawa, Y.; Minami, E.; Okada, K.; Yamane, H.; Kaku, H.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Thao, N.P.; Chen, L.; Nakashima, A.; Hara, S.I.; Umemura, K.; Takahashi, A.; Shirasu, K.; Kawasaki, T.; Shimamoto, K. RAR1 and HSP90 form a complex with Rac/Rop GTPase and function in innate-immune responses in rice. Plant Cell 2007, 19, 4035–4045. [Google Scholar] [CrossRef] [Green Version]
- Seo, N.S.; Lee, S.K.; Song, M.Y.; Suh, J.P.; Hahn, T.R.; Ronald, P.; Jeon, J.S. The HSP90–SGT1–RAR1 molecular chaperone complex: A core modulator in plant immunity. J. Plant Biol. 2008, 51, 1–10. [Google Scholar] [CrossRef]
- Chen, L.; Hamada, S.; Fujiwara, M.; Zhu, T.; Thao, N.P.; Wong, H.L.; Krishna, P.; Ueda, T.; Kaku, H.; Naoto Shibuya, N.; et al. The Hop/Sti1–Hsp90 chaperone complex facilitates the maturation and transport of a PAMP receptor in rice innate immunity. Cell Host Microbe 2010, 7, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Lieberherr, D.; Thao, N.P.; Nakashima, A.; Umemura, K.; Kawasaki, T.; Shimamoto, K. A sphingolipid elicitor-inducible mitogen-activated protein kinase is regulated by the small GTPase OsRac1 and heterotrimeric G-protein in rice. Plant Physiol. 2005, 138, 1644–1652. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Cho, K.; Singh, R.; Jung, Y.H.; Jeong, S.H.; Kim, S.H.; Kim, S.H.; Lee, J.E.; Cho, Y.S.; Agrawal, G.K.; et al. Rice OsACDR1 (Oryza sativa accelerated cell death and resistance 1) is a potential positive regulator of fungal disease resistance. Mol. Cells 2009, 28, 431–439. [Google Scholar] [CrossRef]
- Kishi-Kaboshi, M.; Okada, K.; Kurimoto, L.; Murakami, S.; Umezawa, T.; Shibuya, N.; Yamane, H.; Miyao, A.; Takatsuji, H.; Takahashi, A.; et al. A rice fungal MAMP-responsive MAPK cascade regulates metabolic flow to antimicrobial metabolite synthesis. Plant J. 2010, 63, 599–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, A.; Okada, K.; Miyamoto, K.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. OsTGAP1, a bZIP transcription factor, coordinately regulates the inductive production of diterpenoid phytoalexins in rice. J. Biol. Chem. 2009, 284, 26510–26518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokotani, N.; Sato, Y.; Tanabe, S.; Chujo, T.; Shimizu, T.; Okada, K.; Yamane, H.; Shimono, M.; Sugano, S.; Takatsuji, H.; et al. OsWRKY76 is a rice transcriptional repressor playing opposite roles in blast disease resistance and cold stress tolerance. J. Exp. Bot. 2013, 64, 5085–5097. [Google Scholar] [CrossRef]
- Wong, H.L.; Pinontoan, R.; Hayashi, K.; Tabata, R.; Yaeno, T.; Hasegawa, K.; Kojima, C.; Yoshioka, H.; Iba, L.; Kawasaki, T.; et al. Regulation of rice NADPH oxidase by binding of Rac GTPase to its N-terminal extension. Plant Cell 2007, 19, 4022–4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, E.; Wong, H.L.; Kawasaki, T.; Hasegawa, M.; Kodama, O.; Shimamoto, K. Essential role of the small GTPase Rac in disease resistance of rice. Proc. Natl Acad. Sci. USA 2001, 98, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Ron, M.; Avni, A. The receptor for the fungal elicitor ethylene-inducing xylanase is a member of a resistance-like gene family in tomato. Plant Cell 2004, 16, 1604–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurusu, T.; Hamada, J.; Nokajima, H.; Kitagawa, Y.; Kiyoduka, M.; Takahashi, A.; Hanamata, S.; Ohno, R.; Hayashi, T.; Okada, K.; et al. Regulation of microbe-associated molecular pattern-induced hypersensitive cell death, phytoalexin production, and defense gene expression by calcineurin B-like protein-interacting protein kinases, OsCIPK14/15, in rice cultured cells. Plant Physiol. 2010, 153, 678–692. [Google Scholar] [CrossRef] [Green Version]
- Hamada, H.; Kurusu, T.; Okuma, E.; Nokajima, H.; Kiyoduka, M.; Koyano, T.; Sugiyama, Y.; Okada, K.; Koga, J.; Saji, H.; et al. Regulation of a proteinaceous elicitor-induced Ca2+ influx and production of phytoalexins by a putative voltage-gated cation channel, OsTPC1, in cultured rice cells. J. Biol. Chem. 2012, 287, 9931–9939. [Google Scholar] [CrossRef] [Green Version]
- Riemann, M.; Haga, K.; Shimizu, T.; Okada, K.; Ando, S.; Mochizuki, S.; Nishizawa, Y.; Yamanouchi, U.; Nick, P.; Yano, M.; et al. Identification of rice allene oxide cyclase mutants and the function of jasmonate for defense against Magnaporthe oryzae. Plant J. 2013, 74, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Daw, B.D.; Zhang, L.H.; Wang, Z.Z. Salicylic acid enhances antifungal resistance to Magnaporthe grisea in rice plants. Aust. Plant Pathol. 2008, 37, 637–644. [Google Scholar] [CrossRef]
- Kariya, K.; Ube, N.; Ueno, M.; Teraishi, M.; Okumoto, Y.; Mori, N.; Ueno, L.; Ishihara, A. Natural variation of diterpenoid phytoalexins in cultivated and wild rice species. Phytochemistry 2020, 180, 112518. [Google Scholar] [CrossRef]
- Miyamoto, K.; Fujita, M.; Shenton, M.R.; Akashi, S.; Sugawara, C.; Sakai, A.; Horie, K.; Hasegawa, M.; Kawaide, H.; Mitsuhashi, W.; et al. Evolutionary trajectory of phytoalexin biosynthetic gene clusters in rice. Plant J. 2016, 87, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Qiu, J.; Ye, C.; Jin, G.; Mao, L.; Zhang, H.; Yang, X.; Peng, Q.; Wang, Y.; Jia, L.; et al. Echinochloa crus-galli genome analysis provides insight into its adaptation and invasiveness as a weed. Nat. Commun. 2017, 8, 1031. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Matsuo, A. Applied bryology. In Advance in Bryology, vol. 2.; W. Schultze-Motel, W., Ed.; International Association of Bryologists: Vaduz, Liechtenstein, 1984; pp. 133–224. [Google Scholar]
- Tsubota, H.; Kuroda, A.; Masuzaki, H.; Nakahara, M.; Deguchi, H. Preliminary study on allelopathic activity of bryophytes under laboratory conditions using the sandwich method. J. Hattori Bot. Lab. 2006, 100, 517–525. [Google Scholar]
- Okada, K.; Kawaide, H.; Miyamoto, K.; Miyazaki, S.; Kainuma, R.; Kimura, H.; Fujiwara, K.; Natsume, M.; Nojiri, H.; Nakajima, M.; et al. HpDTC1, a stress-inducible bifunctional diterpene cyclase involved in momilactone biosynthesis, functions in chemical defense in the moss Hypnum plumaeforme. Sci. Rep. 2006, 6, 1–12. [Google Scholar]
- Mao, L.; Kawaide, H.; Higuchi, T.; Chen, M.; Miyamoto, K.; Hirata, Y.; Kimura, H.; Miyazaki, S.; Teruya, M.; Fujiwara, K.; et al. Genomic evidence for convergent evolution of gene clusters for momilactone biosynthesis in land plants. Proc. Natl Acad. Sci. USA 2020, 117, 12472–12480. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Secretion of momilactone A and B by the moss Hypnum plumaeforme. Plant Signal. Behav. 2009, 4, 737–739. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Momilactones.

Figure 2.
Biosynthetic pathway of momilactones in rice.

Figure 3.
Gene cluster on chromosome 4 of rice genome for momilactone biosynthesis.

Figure 4.
Signaling pathway for the elicitation of rice momilactone biosynthesis.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The concentrations (μM) required for 50% growth inhibition (IC50) of various plant species.
Table 1.
The concentrations (μM) required for 50% growth inhibition (IC50) of various plant species.
| Momilactone A | Momilactone B | Reference | |||
|---|---|---|---|---|---|
| Target Plant Species | Roots | Shoots | Roots | Shoots | |
| Echinochola crus-gall | 28.7 | 46.4 | 6.1 | 6.3 | [53] |
| Echinochloa colonum | 65.4 | 240 | 5.04 | 12.5 | [52] |
| Phleum pratense | 76.5 | 157 | 5.6 | 7.9 | [55] |
| Digitaria sanguinalis | 98.5 | 275 | 9.5 | 12.4 | [55] |
| Lolium multiflorum | 91.9 | 138 | 6.9 | 6.5 | [55] |
| Arabidopsis thiliana | 203 | 84.4 | 12 | 6.5 | [54] |
| Lepidium sativum | 425 | 285 | 6.3 | 4.6 | [35] |
| Lactuca sativa | 472 | 395 | 54.3 | 77.9 | [55] |
| Medicago sativa | 379 | 315 | 67.8 | 82.4 | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kato-Noguchi, H. Defensive Molecules Momilactones A and B: Function, Biosynthesis, Induction and Occurrence. Toxins 2023, 15, 241. https://doi.org/10.3390/toxins15040241
AMA Style
Kato-Noguchi H. Defensive Molecules Momilactones A and B: Function, Biosynthesis, Induction and Occurrence. Toxins. 2023; 15(4):241. https://doi.org/10.3390/toxins15040241
Chicago/Turabian StyleKato-Noguchi, Hisashi. 2023. "Defensive Molecules Momilactones A and B: Function, Biosynthesis, Induction and Occurrence" Toxins 15, no. 4: 241. https://doi.org/10.3390/toxins15040241
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.