Zearalenone and Its Emerging Metabolites Promptly Affect the Rumen Microbiota in Holstein Cows Fed a Forage-Rich Diet

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Feed Intake and Zearalenone Exposure Level
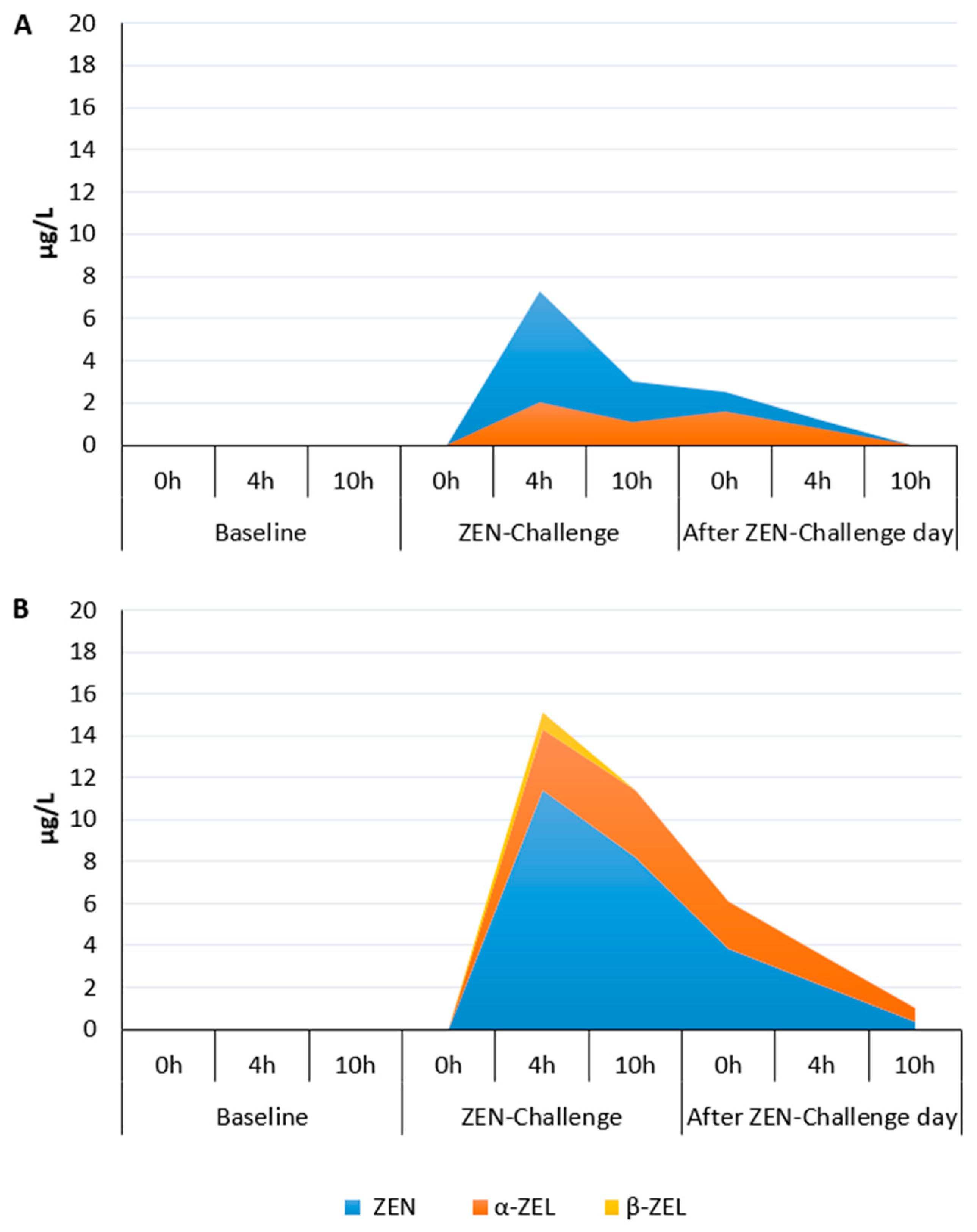
2.2. Changes in Concentrations of Zearalenone and Its Metabolites in the Rumen
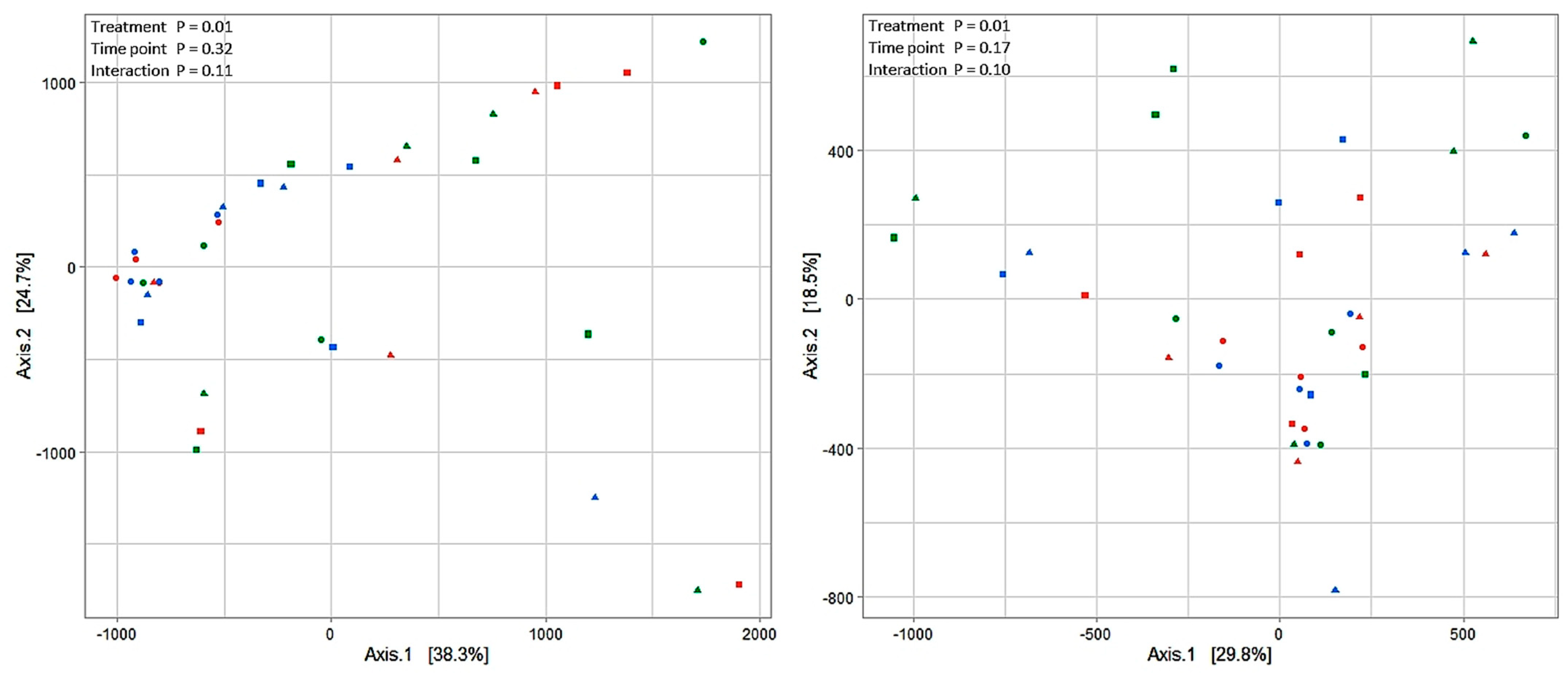
2.3. Prokaryotic Microbiota Composition in the Rumen
2.4. Absolute Abundances of Microbial Groups in the Rumen
2.5. Short-Chain Fatty Acid Profiles in the Rumen
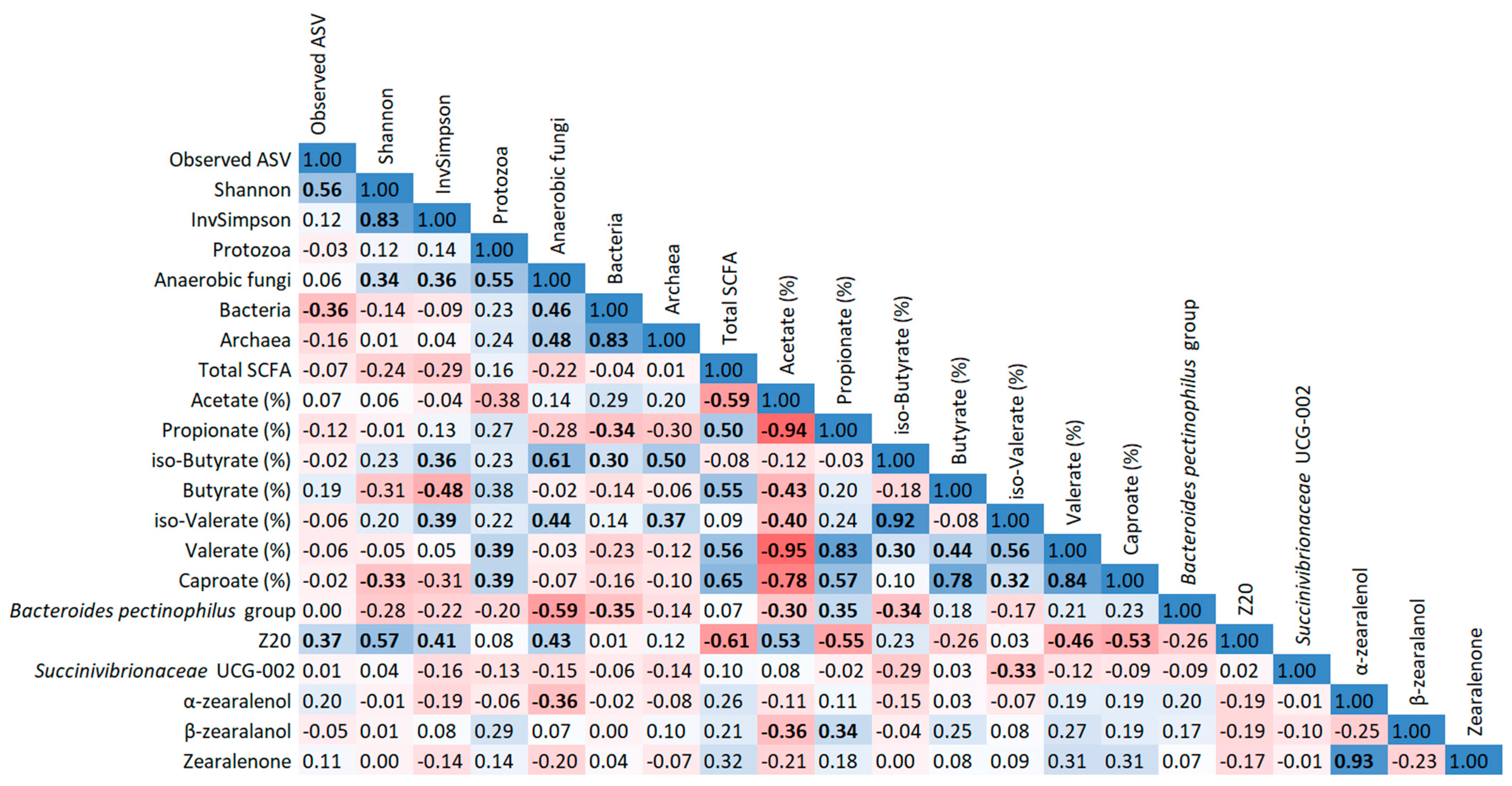
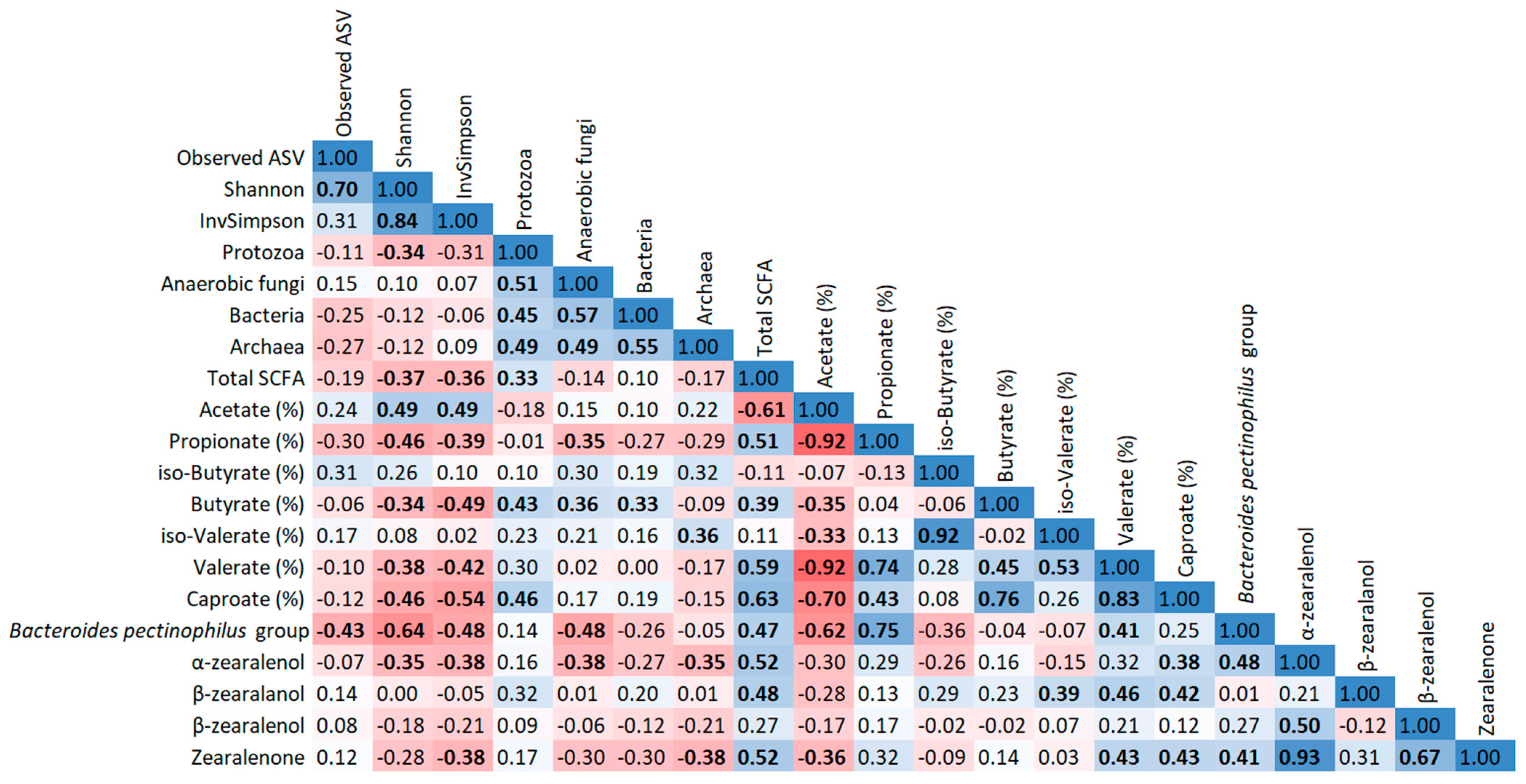
2.6. Correlation Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Feeding
5.2. Experimental Design and Oral ZEN Challenge
5.3. Ruminal Samplings
5.4. Analysis of Feedstuffs
5.5. DNA Extraction and Prokaryotic 16S rRNA Gene Sequencing
5.6. qPCR Analysis
5.7. Analysis of Short-Chain Fatty Acids, Zearalenone and Its Metabolites
5.8. Bioinformatic and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemboi, D.C.; Antonissen, G.; Ochieng, P.E.; Croubels, S.; Okoth, S.; Kangethe, E.K.; Faas, J.; Lindahl, J.F.; Gathumbi, J.K. A review of the impact of mycotoxins on dairy cattle health: Challenges for food safety and dairy production in Sub-Saharan Africa. Toxins 2020, 12, 222. [Google Scholar] [CrossRef] [Green Version]
- Seeling, K.; Dänicke, S. Relevance of the Fusarium toxins deoxynivalenol and zearalenone in ruminant nutrition. A review. J. Anim. Feed Sci. 2005, 14, 3–40. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef]
- Hartinger, T.; Grabher, L.; Pacífico, C.; Angelmayr, B.; Faas, J.; Zebeli, Q. Short-term exposure to the mycotoxins zearalenone or fumonisins affects rumen fermentation and microbiota, and health variables in cattle. Food Chem. Toxicol. 2022, 162, 112900. [Google Scholar] [CrossRef]
- Debevere, S.; Cools, A.; de Baere, S.; Haesaert, G.; Rychlik, M.; Croubels, S.; Fievez, V. In vitro rumen simulations show a reduced disappearance of deoxynivalenol, nivalenol and enniatin B at conditions of rumen acidosis and lower microbial activity. Toxins 2020, 12, 101. [Google Scholar] [CrossRef] [Green Version]
- Hua, C.; Tian, J.; Tian, P.; Cong, R.; Luo, Y.; Geng, Y.; Tao, S.; Ni, Y.; Zhao, R. Feeding a high concentration diet induces unhealthy alterations in the composition and metabolism of ruminal microbiota and host response in a goat model. Front. Microbiol. 2017, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Gruber-Dorninger, C.; Faas, J.; Doupovec, B.; Aleschko, M.; Stoiber, C.; Höbartner-Gußl, A.; Schöndorfer, K.; Killinger, M.; Zebeli, Q.; Schatzmayr, D. Metabolism of zearalenone in the rumen of dairy cows with and without application of a zearalenone-degrading enzyme. Toxins 2021, 13, 84. [Google Scholar] [CrossRef]
- Scheurich, A.; Penicka, A.; Hörtenhuber, S.; Lindenthal, T.; Quendler, E.; Zollitsch, W. Elements of social sustainability among Austrian hay milk farmers: Between satisfaction and stress. Sustainability 2021, 13, 13010. [Google Scholar] [CrossRef]
- Daou, R.; Joubrane, K.; Maroun, R.G.; Khabbaz, L.R.; Ismail, A.; Khoury, A.E. Mycotoxins: Factors influencing production and control strategies. AIMS Agric. Food 2021, 6, 416–447. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Khiaosa-Ard, R.; Nagl, V.; Faas, J.; Jenkins, T.; Sulyok, M.; Zebeli, Q. Mycotoxins, phytoestrogens and other secondary metabolites in Austrian pastures: Occurrences, contamination levels and implications of geo-climatic factors. Toxins 2021, 13, 460. [Google Scholar] [CrossRef]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [CrossRef] [Green Version]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef]
- European Commission. Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (2006/576/EC). Off. J. Eur. Union 2006, L 229, 7–9. [Google Scholar]
- Blackburn, T.H.; Hungate, R.E. Succinic acid turnover and propionate production in the bovine rumen. Appl. Microbiol. 1963, 11, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yau, Y.; Zhu, J.; Wang, Y.; Dai, Z.; Gan, H.; Qian, L.; Yang, Z. Effect of electroacupuncture at zusanli (ST36) on intestinal microbiota in rats with chronic atrophic gastritis. Front. Genet. 2022, 13, 824739. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain. Appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, e04425. [Google Scholar] [CrossRef]
- Zhang, Y.; Tao, X.; Liu, Q.; Zhang, Y.J.; Xu, J.; Zhang, W.; Wang, J.; Zhang, D.; Li, B.; Wang, L.; et al. Succession changes of fermentation parameters, nutrient components and bacterial community of sorghum stalk silage. Front. Microbiol. 2022, 13, 982489. [Google Scholar] [CrossRef]
- Edwards, J.E.; Kingston-Smith, A.H.; Jimenez, H.R.; Huws, S.A.; Skøt, K.P.; Griffith, G.W.; McEwan, N.R.; Theodorou, M.K. Dynamics of initial colonization of nonconserved perennial ryegrass by anaerobic fungi in the bovine rumen. FEMS Microbiol. Ecol. 2008, 66, 537–545. [Google Scholar] [CrossRef]
- Westlake, K.; Mackie, R.I.; Dutton, M.F. In vitro metabolism of mycotoxins by bacterial, protozoal and ovine ruminal fluid preparations. Anim. Feed Sci. Technol. 1989, 25, 169–178. [Google Scholar] [CrossRef]
- Firkins, J.L.; Yu, Z.; Park, T.; Plank, J.E. Extending Burk Dehority’s perspectives on the role of ciliate protozoa in the rumen. Front. Microbiol. 2020, 11, 123. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Puniya, A.K.; Callaghan, T.M.; Edwards, J.E.; Youssef, N.; Dagar, S.S.; Fliegerova, K.; Griffith, G.W.; Forster, R.; Tsang, A.; et al. Anaerobic fungi (phylum Neocallimastigomycota): Advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiol. Ecol. 2014, 90, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Elliott, R.; Ash, A.J.; Calderon-Cortes, F.; Norton, B.W.; Bauchop, T. The influence of anaerobic fungi on rumen volatile fatty acid concentrations in vivo. J. Agric. Sci. 1987, 109, 13–17. [Google Scholar] [CrossRef]
- VDLUFA. VDLUFA-Methodenbuch Bd. III Die chemische Untersuchung von Futtermitteln, 3rd ed.; VDLUFA-Verlag: Darmstadt, Germany, 2012; ISBN 9783941273146. [Google Scholar]
- Klevenhusen, F.; Petri, R.M.; Kleefisch, M.-T.; Khiaosa-Ard, R.; Metzler-Zebeli, B.U.; Zebeli, Q. Changes in fibre-adherent and fluid-associated microbial communities and fermentation profiles in the rumen of cattle fed diets differing in hay quality and concentrate amount. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, M.; Adamowicz, E.; Basarab, J.A.; Le Guan, L. Characterization of bovine ruminal epithelial bacterial communities using 16S rRNA sequencing, PCR-DGGE, and qRT-PCR analysis. Vet. Microbiol. 2012, 155, 72–80. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Hernandez-Sanabria, E.; Le Guan, L. Assessment of the microbial ecology of ruminal methanogens in cattle with different feed efficiencies. Appl. Environ. Microbiol. 2009, 75, 6524–6533. [Google Scholar] [CrossRef] [Green Version]
- Sylvester, J.T.; Karnati, S.K.R.; Yu, Z.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef]
- Poier, G.; Terler, G.; Klevenhusen, F.; Sharma, S.; Zebeli, Q. Replacing concentrates with a high-quality hay in the starter feed of dairy calves: II. Effects on the development of chewing and gut fermentation, and selected systemic health variables. J. Dairy Sci. 2022, 105, 3113–3128. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech Xu, Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur rapidly resolves single-nucleotide community sequence patterns. mSystems 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable association discovery in population-scale meta-omics studies. BioRxiv, 2021; preprint. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 9783319242774. [Google Scholar]
- Bisanz, J.E. Qiime2r: Importing QIIME2 Artifacts and Associated Data into R Sessions. 2018. Available online: https://github.com/jbisanz/qiime2R (accessed on 25 October 2022).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Baseline | ZEN-Challenge Day | After ZEN-Challenge Day | p-Values 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time after Feeding | 0 h | 4 h | 10 h | Mean | 0 h | 4 h | 10 h | Mean | 0 h | 4 h | 10 h | Mean | SEM 1 | Trt | Time | Trt × Time |
| FRL | ||||||||||||||||
| Observed ASV | 3918 A ab | 3983 a | 3556 b | 3819 | 3136 Bb | 3760 a | 3720 a | 3539 | 4182 Aa | 3683 b | 3813 ab | 3892 | 187 | 0.23 | 0.61 | 0.01 |
| Shannon | 7.32 | 7.34 | 7.18 | 7.28 X | 7.03 | 7.21 | 7.08 | 7.11 Y | 7.13 | 7.16 | 7.03 | 7.11 Y | 0.07 | 0.03 | 0.03 | 0.54 |
| InvSimpson | 464 | 456 | 352 | 424 X | 241 | 352 | 207 | 267 Y | 226 | 367 | 234 | 276 Y | 49.1 | <0.01 | 0.01 | 0.60 |
| PARL | ||||||||||||||||
| Observed ASV | 3236 | 3165 | 3190 | 3197 | 3337 | 3509 | 3089 | 3312 | 3067 | 3189 | 2958 | 3071 | 114 | 0.15 | 0.03 | 0.26 |
| Shannon | 7.13 a | 7.08 b | 7.08 b | 7.10 | 7.11 a | 7.07 a | 6.97 b | 7.05 | 7.09 a | 7.06 a | 6.92 b | 7.02 | 0.04 | 0.30 | <0.01 | 0.03 |
| InvSimpson | 383 | 359 | 343 | 362 | 352 | 314 | 254 | 307 | 385 | 343 | 247 | 325 | 22.9 | 0.10 | <0.01 | 0.09 |
| Treatment | Baseline | ZEN-Challenge-Day | After ZEN-Challenge Day | p-Values 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time after Feeding | 0 h | 4 h | 10 h | Mean | 0 h | 4 h | 10 h | Mean | 0 h | 4 h | 10 h | Mean | SEM 1 | Trt | Time | Trt × Time |
| FRL | ||||||||||||||||
| Bacteria | 6.09 | 6.11 | 6.18 | 6.12 | 6.18 | 6.08 | 6.05 | 6.10 | 5.97 | 6.09 | 5.94 | 6.00 | 0.08 | 0.19 | 0.79 | 0.36 |
| Archaea | 4.69 | 4.82 | 4.72 | 4.74 | 4.75 | 4.62 | 4.59 | 4.65 | 4.59 | 4.70 | 4.52 | 4.60 | 0.14 | 0.39 | 0.62 | 0.85 |
| Protozoa | 4.14 | 4.61 | 4.80 | 4.51 | 4.04 | 4.56 | 4.63 | 4.41 | 3.87 | 4.36 | 4.52 | 4.25 | 0.14 | 0.13 | <0.01 | 0.71 |
| Anaerobic fungi | 3.76 | 4.09 | 3.83 | 3.89 X | 3.51 | 3.67 | 3.50 | 3.56 Y | 3.30 | 3.67 | 3.46 | 3.48 Y | 0.11 | <0.01 | <0.01 | 0.76 |
| PARL | ||||||||||||||||
| Bacteria | 6.17 | 6.10 | 6.16 | 6.15 | 6.20 | 6.12 | 6.08 | 6.13 | 6.16 | 6.14 | 6.26 | 6.18 | 0.06 | 0.30 | 0.33 | 0.42 |
| Archaea | 5.29 a | 5.23 Aa | 4.85 Bb | 5.12 | 5.18 a | 4.90 Bab | 4.86 Bb | 4.98 | 5.06 | 5.06 AB | 5.22 A | 5.11 | 0.12 | 0.27 | 0.08 | 0.04 |
| Protozoa | 4.43 | 4.59 | 4.43 | 4.48 Y | 4.61 | 4.72 | 4.52 | 4.61 XY | 4.66 | 4.72 | 4.81 | 4.73 X | 0.09 | 0.02 | 0.27 | 0.34 |
| Anaerobic fungi | 3.82 | 4.11 | 3.90 | 3.95 | 4.03 | 3.94 | 3.68 | 3.88 | 3.77 | 3.95 | 4.02 | 3.91 | 0.13 | 0.80 | 0.28 | 0.10 |
| Treatment | Baseline | ZEN-Challenge-Day | After ZEN-Challenge Day | p-Values 2 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time after Feeding | 0 h | 4 h | 10 h | Mean | 0 h | 4 h | 10 h | Mean | 0 h | 4 h | 10 h | Mean | SEM1 | Trt | Time | Trt × Time |
| FRL | ||||||||||||||||
| Total SCFA | 67.0 | 66.4 | 88.6 | 74.0 Y | 82.7 | 98.1 | 100.5 | 93.8 X | 78.7 | 105.8 | 98.5 | 94.3 X | 6.88 | 0.02 | 0.01 | 0.30 |
| Acetate | 72.3 | 69.4 | 69.0 | 70.2 | 72.2 | 68.3 | 67.7 | 69.4 | 71.7 | 67.8 | 67.1 | 68.9 | 0.74 | 0.06 | <0.01 | 0.66 |
| Propionate | 16.2 | 17.9 | 19.1 | 17.7 | 16.3 | 18.8 | 19.0 | 18.0 | 16.8 | 19.2 | 19.7 | 18.5 | 0.73 | 0.06 | <0.01 | 0.71 |
| n-Butyrate | 8.54 | 8.89 | 8.86 | 8.76 | 8.82 | 8.98 | 9.62 | 9.14 | 8.92 | 9.18 | 9.60 | 9.23 | 0.24 | 0.08 | <0.01 | 0.07 |
| iso-Butyrate | 0.83 Aa | 0.91 Aa | 0.73 b | 0.82 X | 0.71 Ab | 0.83 Aba | 0.75 b | 0.76 XY | 0.64 Bb | 0.78 Ba | 0.71 b | 0.71 Y | 0.04 | 0.03 | <0.01 | 0.02 |
| n-Valerate | 0.80 | 1.26 | 1.10 | 1.05 | 0.80 | 1.43 | 1.43 | 1.22 | 0.85 | 1.42 | 1.40 | 1.22 | 0.08 | 0.06 | <0.01 | 0.17 |
| iso-Valerate | 1.08 Ab | 1.28 a | 0.89 c | 1.08 | 0.88 Bb | 1.19 a | 0.98 b | 1.02 | 0.77 Bb | 1.15 a | 0.99 a | 0.97 | 0.08 | 0.26 | <0.01 | 0.04 |
| Caproate | 0.30 b | 0.39 Ba | 0.37 Ba | 0.35 Y | 0.32 c | 0.48 Ab | 0.57 Aa | 0.46 X | 0.34 b | 0.53 Aa | 0.55 Aa | 0.47 X | 0.03 | <0.01 | <0.01 | 0.01 |
| PARL | ||||||||||||||||
| Total SCFA | 79.7 b | 75.3 Bb | 94.1 a | 83.0 Y | 85.8 b | 102.9 Aa | 97.7 a | 95.5 X | 91.0 b | 108.3 Aa | 99.1 ab | 99.5 X | 6.04 | 0.02 | 0.01 | 0.03 |
| Acetate | 72.1 | 69.2 | 68.7 | 70.0 | 71.9 | 68.3 | 67.9 | 69.4 | 71.6 | 67.9 | 67.1 | 68.9 | 0.67 | 0.06 | <0.01 | 0.69 |
| Propionate | 16.3 | 17.9 | 19.1 | 17.8 | 16.4 | 18.6 | 19.3 | 18.1 | 16.9 | 19.0 | 19.6 | 18.5 | 0.73 | 0.08 | <0.01 | 0.89 |
| n-Butyrate | 8.64 | 9.01 | 8.92 | 8.86 | 8.88 | 9.12 | 9.29 | 9.10 | 8.85 | 9.16 | 9.64 | 9.21 | 0.25 | 0.29 | <0.01 | 0.25 |
| iso-Butyrate | 0.82 Ab | 0.91 Aa | 0.74 c | 0.82 X | 0.71 Bb | 0.83 Ba | 0.72 b | 0.75 Y | 0.65 Bb | 0.78 Ba | 0.71 b | 0.71 Y | 0.04 | 0.01 | <0.01 | 0.05 |
| n-Valerate | 0.84 | 1.30 | 1.16 | 1.10 | 0.85 | 1.48 | 1.38 | 1.24 | 0.89 | 1.46 | 1.43 | 1.26 | 0.07 | 0.08 | <0.01 | 0.47 |
| iso-Valerate | 1.08 Ab | 1.28 a | 0.91 c | 1.09 | 0.88 Bb | 1.17 a | 0.94 b | 1.00 | 0.79 Bc | 1.13 a | 0.98 b | 0.96 | 0.07 | 0.14 | <0.01 | 0.03 |
| Caproate | 0.31 | 0.41 | 0.41 | 0.37 Y | 0.35 | 0.53 | 0.54 | 0.47 X | 0.37 | 0.57 | 0.58 | 0.51 X | 0.04 | 0.01 | <0.01 | 0.27 |
| Target | Item | Primer Sequence (5′–3′) | Annealing Temperature (°C) | Primer Concentration (nmol) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|---|
| Bacteria | F 1 | CCTACGGGAGGCAGCAG | 61 | 100 | 189 | [27] |
| R 2 | ATTACCGCGGCTGCTGG | |||||
| Archaea | F | CCGGAGATGGAACCTGAGAC | 60 | 100 | 160 | [28] |
| R | CGGTCTTGCCCAGCTCTTATTC | |||||
| Protozoa | F | GCTTTCGWTGGTAGTGTATT | 60 | 400 | 233 | [29] |
| R | CTTGCCCTCYAATCGTWCT | |||||
| Anaerobic fungi | F | GAGGAAGTAAAAGTCGTAACAAGGTTTC | 60 | 200 | 110–115 | [30] |
| R | CAAATTCACAAAGGGTAGGATGATT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hartinger, T.; Kröger, I.; Neubauer, V.; Faas, J.; Doupovec, B.; Schatzmayr, D.; Zebeli, Q. Zearalenone and Its Emerging Metabolites Promptly Affect the Rumen Microbiota in Holstein Cows Fed a Forage-Rich Diet. Toxins 2023, 15, 185. https://doi.org/10.3390/toxins15030185
Hartinger T, Kröger I, Neubauer V, Faas J, Doupovec B, Schatzmayr D, Zebeli Q. Zearalenone and Its Emerging Metabolites Promptly Affect the Rumen Microbiota in Holstein Cows Fed a Forage-Rich Diet. Toxins. 2023; 15(3):185. https://doi.org/10.3390/toxins15030185
Chicago/Turabian StyleHartinger, Thomas, Iris Kröger, Viktoria Neubauer, Johannes Faas, Barbara Doupovec, Dian Schatzmayr, and Qendrim Zebeli. 2023. "Zearalenone and Its Emerging Metabolites Promptly Affect the Rumen Microbiota in Holstein Cows Fed a Forage-Rich Diet" Toxins 15, no. 3: 185. https://doi.org/10.3390/toxins15030185