Production of Aflatoxin B1 by Aspergillus parasiticus Grown on a Novel Meat-Based Media


Abstract
:1. Introduction
2. Results and Discussion
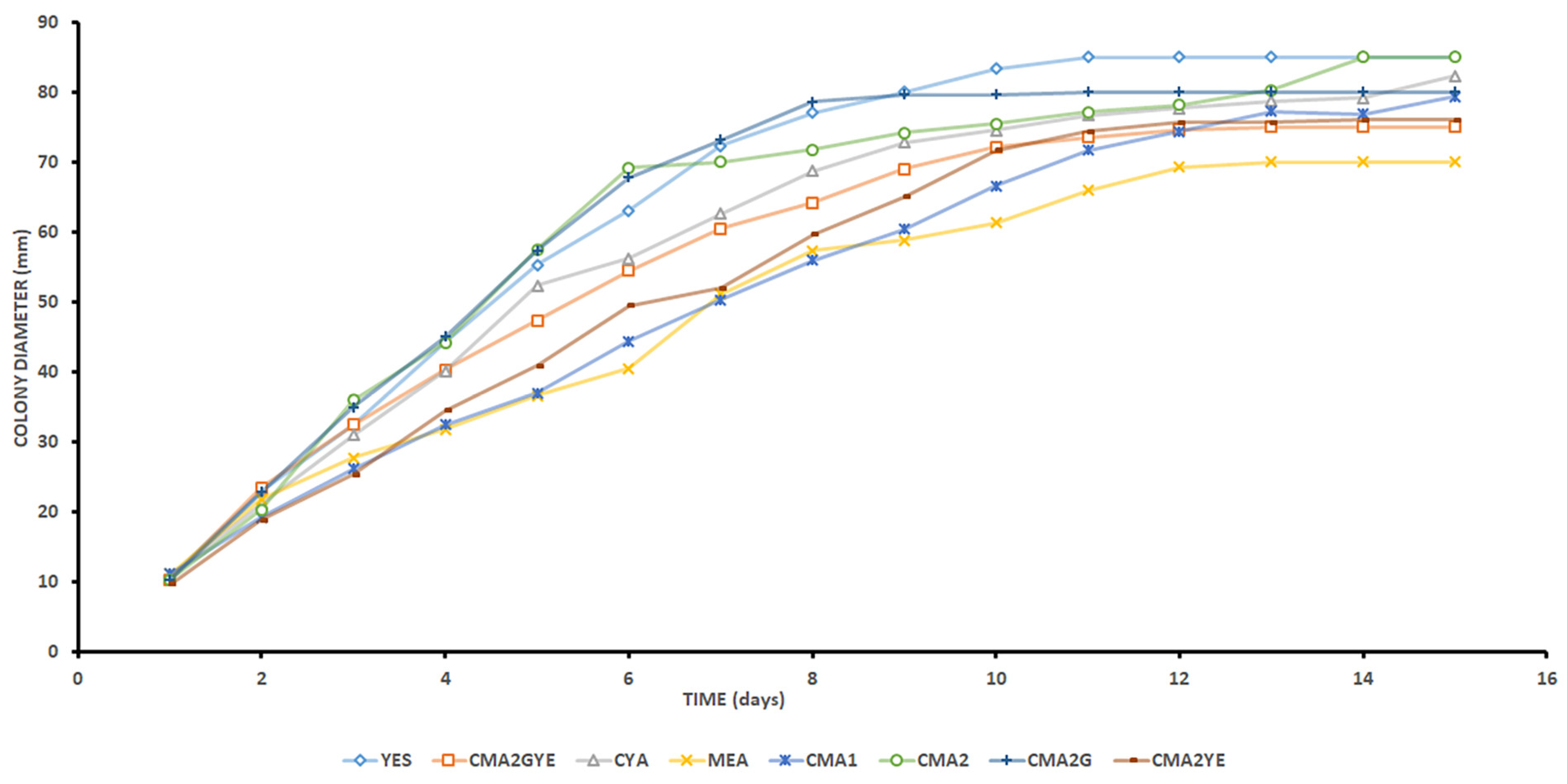
2.1. A. parasiticus Growth Assessment
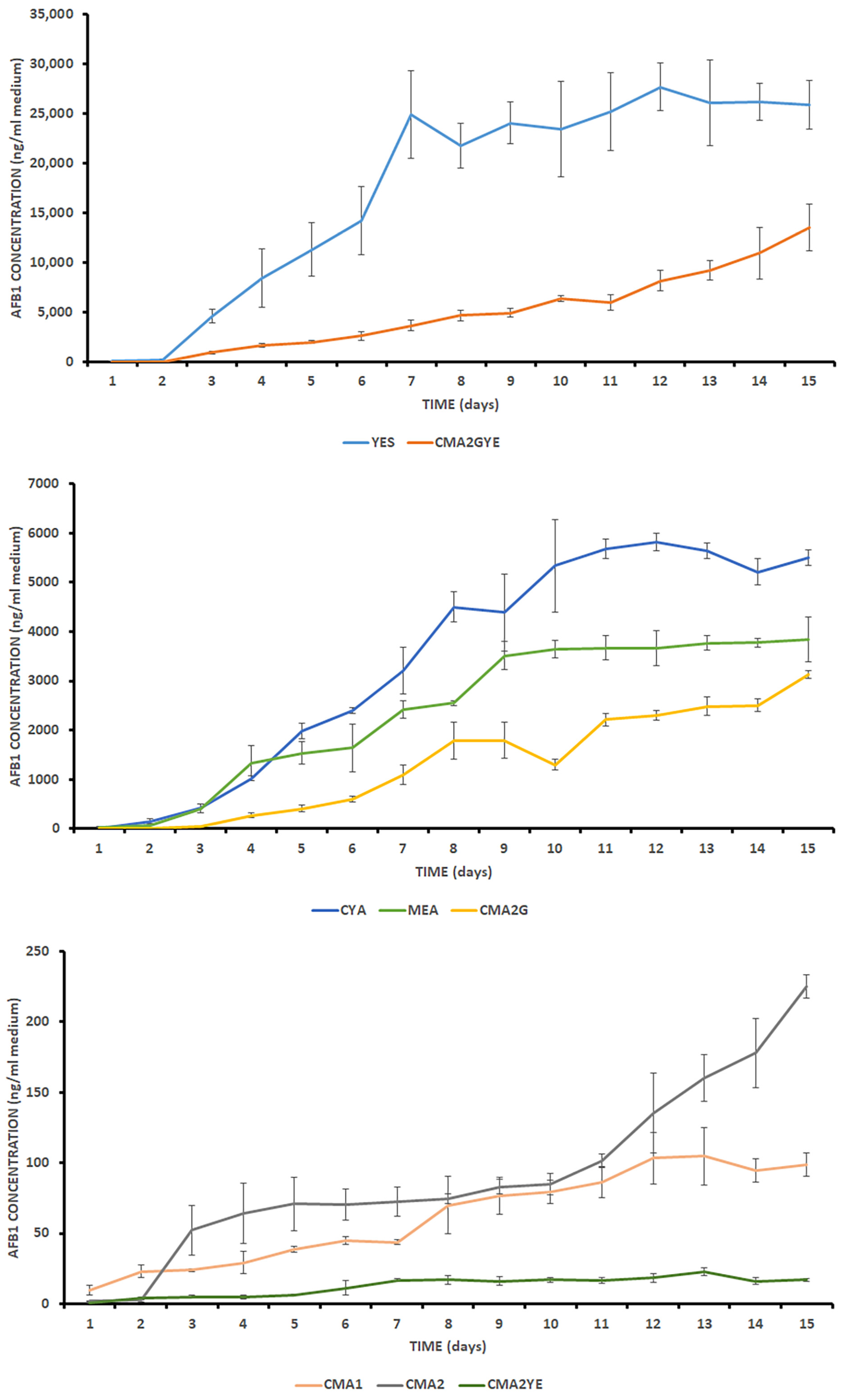
2.2. A. parasiticus AFB1 Production Analysis
2.3. Relationship between Growth and AFB1 Production
3. Conclusions
4. Materials and Methods
4.1. Fungal Strain and Inoculum Preparation
4.2. Media and Experimental Design
- CMA1 was prepared using CMB (Cooked Meat Broth, Merck, Rahway, NJ, USA) with the addition of bacteriological agar (15 g/L);
- CMA2 was prepared as follows: veal bones (4 kg) were purchased from a local market, boiled for 4 h in 4 L of distilled water, and concentrated to 6% dry weight during cooking. The main advantage of this meat-based medium is its ease of preparation, as well as its numerous possibilities for supplementation with different compounds used in meat technologies. The resulting mixture was filtered through a double layer of muslin, and 20 g/L bacteriological agar was added prior to sterilization. The composition of CMA2 was chemically determined (water 94%, ash 1%, fats 0.7%, proteins 4.3%).
- CMA2G: CMA2 medium was supplemented with 1% glucose (D-(+)-glucose, Sigma–Aldrich Chemie GmbH).
- CMA2YE: CMA2 medium was supplemented with 0.2% yeast extract (Sigma–Aldrich Chemie GmbH).
- CMA2GYE: CMA2 medium was supplemented with 1% glucose and 0.2% yeast extract, as these components can be used in the standard production of dry-fermented meat products (and are elements of various microbial culture media).
- YES (20 g/L yeast extract, 150 g/L sucrose, 20 g/L agar, 1 g/L MgSO4 · 7H2O);
- MEA (30 g/L malt extract, 5 g/L mycological peptone, 15 g/L agar);
- CYA (5 g/L yeast extract, 30 g/L saccharose, 15 g/L agar, 10 mL/L concentrated Czapek, 1 g/L K2HPO4).
4.3. A. parasiticus Growth Assessment
4.4. AFB1 Analysis in Growth Media
4.4.1. Purification of Samples
4.4.2. UPLC–MS/MS Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dzuman, Z.; Zachariasova, M.; Veprikova, Z.; Godula, M.; Hajslova, J. Multi-Analyte High Performance Liquid Chromatography Coupled to High Resolution Tandem Mass Spectrometry Method for Control of Pesticide Residues, Mycotoxins, and Pyrrolizidine Alkaloids. Anal. Chim. Acta 2015, 863, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisen, R.; Touhami, N.; Schmidt-Heydt, M. Mycotoxins as Adaptation Factors to Food Related Environments. Curr. Opin. Food Sci. 2017, 17, 1–8. [Google Scholar] [CrossRef]
- Bragulat, M.R.; Abarca, M.L.; Cabañes, F.J. An Easy Screening Method for Fungi Producing Ochratoxin A in Pure Culture. Int. J. Food Microbiol. 2001, 71, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J. Aflatoxin and Sclerotical Production by Aspergillus flavus: Influence of PH. Physiol. Biochem. 1988, 78, 1250–1253. [Google Scholar]
- Adye, J.; Mateles, R.I. Incorporation of Labelled Compounds into Aflatoxins. Biochim. Biophys. Acta Gen. Subj. 1964, 86, 418–420. [Google Scholar] [CrossRef]
- Aziz, N.H.; el-Zeany, S.A.; Moussa, L.A.A. Influence of Gamma-Irradiation and Maize Lipids on the Production of Aflatoxin B1 by Aspergillus flavus. Nahrung 2002, 46, 327–331. [Google Scholar] [CrossRef]
- Luchese, R.H.; Harrigan, W.F. Biosynthesis of Aflatoxin—The Role of Nutritional Factors. J. Appl. Bacteriol. 1993, 74, 5–14. [Google Scholar] [CrossRef]
- Hussein, H.S.; Brasel, J.M. Toxicity, Metabolism, and Impact of Mycotoxins on Humans and Animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Richard, J.L. Some Major Mycotoxins and Their Mycotoxicoses—An Overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef]
- Jaimez, J.; Fente, C.A.; Vazquez, B.I.; Franco, C.M.; Cepeda, A.; Mahuzier, G.; Prognon, P. Application of the Assay of Aflatoxins by Liquid Chromatography with Fluorescence Detection in Food Analysis. J. Chromatogr. A 2000, 882, 1–10. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Aflatoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Press: Lyon, France, 2002; Volume 82. [Google Scholar]
- Vulić, A.; Vahčić, N.; Hengl, B.; Gross-Bošković, A.; Jurković, M.; Kudumija, N.; Pleadin, J. Assessment of Possible Human Exposure to Ochratoxin A in Croatia Due to the Consumption of Dry-Cured and Fermented Meat Products. Food Addit. Contam. Part A 2016, 33, 1428–1434. [Google Scholar] [CrossRef]
- Lešić, T.; Zadravec, M.; Zdolec, N.; Vulić, A.; Perković, I.; Škrivanko, M.; Kudumija, N.; Jakopović, Ž.; Pleadin, J. Mycobiota and Mycotoxin Contamination of Traditional and Industrial Dry-Fermented Sausage Kulen. Toxins 2021, 13, 798. [Google Scholar] [CrossRef]
- Montanha, F.P.; Anater, A.; Burchard, J.F.; Luciano, F.B.; Meca, G.; Manyes, L.; Pimpao, C.T. Mycotoxins in Dry-Cured Meats: A Review. Food Chem. Toxicol. 2018, 111, 494–502. [Google Scholar] [CrossRef]
- Perrone, G.; Rodriguez, A.; Magista, D.; Magan, N. Insights into Existing and Future Fungal and Mycotoxin Contamination of Cured Meats. Curr. Opin. Food Sci. 2019, 29, 20–27. [Google Scholar] [CrossRef]
- World Health Organization; International Agency for Research on Cancer. Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. In IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans; World Health Organization: Lyon, France, 1993; Volume 56. [Google Scholar]
- Iacumin, L.; Chiesa, L.; Boscolo, D.; Manzano, M.; Cantoni, C.; Orlic, S.; Comi, G. Moulds and Ochratoxin A on Surfaces of Artisanal and Industrial Dry Sausages. Food Microbiol. 2009, 26, 65–70. [Google Scholar] [CrossRef]
- Iacumin, L.; Milesi, S.; Pirani, S.; Comi, G.; Chiesa, L.M. Ochratoxigenic Mold and Ochratoxin a in Fermented Sausages from Different Areas in Northern Italy: Occurrence, Reduction or Prevention with Ozonated Air. J. Food Saf. 2011, 31, 538–545. [Google Scholar] [CrossRef]
- Markov, K.; Pleadin, J.; Bevardi, M.; Vahčić, N.; Sokolić-Mihalak, D.; Frece, J. Natural Occurrence of Aflatoxin B1, Ochratoxin A and Citrinin in Croatian Fermented Meat Products. Food Control 2013, 34, 312–317. [Google Scholar] [CrossRef]
- Zadravec, M.; Vahčić, N.; Brnić, D.; Markov, K.; Frece, J.; Beck, R.; Lešić, T.; Pleadin, J. A Study of Surface Moulds and Mycotoxins in Croatian Traditional Dry-Cured Meat Products. Int. J. Food Microbiol. 2020, 317, 108459. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Galaverna, G.; Bertuzzi, T.; Moseriti, A.; Pietri, A.; Dossena, A.; Marchelli, R. Occurrence of Ochratoxin A in Raw Ham Muscle, Salami and Dry-Cured Ham from Pigs Fed with Contaminated Diet. Food Chem. 2010, 120, 978–983. [Google Scholar] [CrossRef]
- Rodríguez, A.; Rodríguez, M.; Martín, A.; Delgado, J.; Córdoba, J.J. Presence of Ochratoxin A on the Surface of Dry-Cured Iberian Ham after Initial Fungal Growth in the Drying Stage. Meat Sci. 2012, 92, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Andrade, M.J.; Delgado, J.; Sánchez-Montero, L.; Núñez, F. Biocontrol of Aflatoxigenic Aspergillus parasiticus by Native Debaryomyces hansenii in Dry-Cured Meat Products. Food Microbiol. 2019, 82, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Roncada, P.; Altafini, A.; Fedrizzi, G.; Guerrini, A.; Polonini, G.L.; Caprai, E. Ochratoxin A Contamination of the Casing and the Edible Portion of Artisan Salamis Produced in Two Italian Regions. World Mycotoxin J. 2020, 13, 553–562. [Google Scholar] [CrossRef]
- Pleadin, J.; Zadravec, M.; Brnic, D.; Perkovic, I.; Skrivanko, M.; Kovacevic, D. Moulds and Mycotoxins Detected in the Regional Speciality Fermented Sausage “slavonski Kulen’ during a 1-Year Production Period. Food Addit. Contam. Part A—Chem. 2017, 34, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Parussolo, G.; Oliveira, M.S.; Garcia, M.V.; Bernardi, A.O.; Lemos, J.G.; Stefanello, A.; Mallmann, C.A.; Copetti, M.V. Ochratoxin A Production by Aspergillus westerdijkiae in Italian-Type Salami. Food Microbiol. 2019, 83, 134–140. [Google Scholar] [CrossRef]
- Algammal, A.M.; Elsayed, M.E.; Hashem, H.R.; Ramadan, H.; Sheraba, N.S.; El-Diasty, E.M.; Abbas, S.M.; Hetta, H.F. Molecular and HPLC-Based Approaches for Detection of Aflatoxin B1 and Ochratoxin A Released from Toxigenic Aspergillus Species in Processed Meat. BMC Microbiol. 2021, 21, 82. [Google Scholar] [CrossRef]
- Kudumija, N.; Vulić, A.; Lešić, T.; Vahčić, N.; Pleadin, J. Aflatoxins and Ochratoxin A in Dry-Fermented Sausages in Croatia, by LC-MS/MS. Food Addit. Contam. Part B Surveill. 2020, 13, 225–232. [Google Scholar] [CrossRef]
- Pleadin, J.; Staver, M.M.; Vahčić, N.; Kovačević, D.; Milone, S.; Saftić, L.; Scortichini, G. Survey of Aflatoxin B1 and Ochratoxin A Occurrence in Traditional Meat Products Coming from Croatian Households and Markets. Food Control 2015, 52, 71–77. [Google Scholar] [CrossRef]
- Aziz, N.H.; Youssef, Y.A. Occurrence of Aflatoxins and Aflatoxin-producing Moulds in Fresh and Processed Meat in Egypt. Food Addit. Contam. 1991, 8, 321–331. [Google Scholar] [CrossRef]
- Cvetnić, Z.; Pepeljnjak, S. Aflatoxin-Producing Potential of Aspergillus flavus and Aspergillus parasiticus Isolated from Samples of Smoked-Dried Meat. Food/Nahr. 1995, 39, 302–307. [Google Scholar] [CrossRef]
- Klich, M.A. Environmental and Developmental Factors Influencing Aflatoxin Production by Aspergillus flavus and Aspergillus parasiticus. Mycoscience 2007, 48, 71–80. [Google Scholar] [CrossRef]
- Lozano-Ojalvo, D.; Rodríguez, A.; Bernáldez, V.; Córdoba, J.J.; Rodríguez, M. Influence of Temperature and Substrate Conditions on the Omt-1 Gene Expression of Aspergillus parasiticus in Relation to Its Aflatoxin Production. Int. J. Food Microbiol. 2013, 166, 263–269. [Google Scholar] [CrossRef]
- Muñoz, K.; Vega, M.; Rios, G.; Geisen, R.; Degen, G.H. Mycotoxin Production by Different Ochratoxigenic Aspergillus and Penicillium Species on Coffee- and Wheat-Based Media. Mycotoxin Res. 2011, 27, 239–247. [Google Scholar] [CrossRef]
- Ritter, A.C.; Hoeltz, M.; Noll, I.B. Toxigenic Potential of Aspergillus flavus Tested in Different Culture Conditions. Cienc. Tecnol. Aliment. 2011, 31, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, M.; Rodríguez, A.; Núñez, F.; Silva, A.; Andrade, M.J. In Vitro Antifungal Effects of Spices on Ochratoxin A Production and Related Gene Expression in Penicillium nordicum on a Dry-Cured Fermented Sausage Medium. Food Control 2020, 114, 107222. [Google Scholar] [CrossRef]
- Battilani, P.; Formenti, S.; Toscani, T.; Virgili, R. Influence of Abiotic Parameters on Ochratoxin A Production by a Penicillium nordicum Strain in Dry-Cured Meat Model Systems. Food Control 2010, 21, 1739–1744. [Google Scholar] [CrossRef]
- Rodríguez, A.; Medina, Á.; Córdoba, J.J.; Magan, N. The Influence of Salt (NaCl) on Ochratoxin A Biosynthetic Genes, Growth and Ochratoxin A Production by Three Strains of Penicillium nordicum on a Dry-Cured Ham-Based Medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef]
- Rodríguez, A.; Capela, D.; Medina, Á.; Córdoba, J.J.; Magan, N. Relationship between Ecophysiological Factors, Growth and Ochratoxin A Contamination of Dry-Cured Sausage Based Matrices. Int. J. Food Microbiol. 2015, 194, 71–77. [Google Scholar] [CrossRef]
- Peromingo, B.; Rodríguez, A.; Bernáldez, V.; Delgado, J.; Rodríguez, M. Effect of Temperature and Water Activity on Growth and Aflatoxin Production by Aspergillus flavus and Aspergillus parasiticus on Cured Meat Model Systems. Meat Sci. 2016, 122, 76–83. [Google Scholar] [CrossRef]
- Sánchez-Montero, L.; Córdoba, J.J.; Peromingo, B.; Álvarez, M.; Núñez, F. Effects of Environmental Conditions and Substrate on Growth and Ochratoxin A Production by Penicillium verrucosum and Penicillium nordicum: Relative Risk Assessment of OTA in Dry-Cured Meat Products. Food Res. Int. 2019, 121, 604–611. [Google Scholar] [CrossRef]
- Sánchez-Montero, L.; Córdoba, J.J.; Alía, A.; Peromingo, B.; Núñez, F. Effect of Spanish Smoked Paprika “Pimentón de La Vera” on Control of Ochratoxin A and Aflatoxins Production on a Dry-Cured Meat Model System. Int. J. Food Microbiol. 2019, 308, 108303. [Google Scholar] [CrossRef] [PubMed]
- Yogendrarajah, P.; Vermeulen, A.; Jacxsens, L.; Mavromichali, E.; De Saeger, S.; De Meulenaer, B.; Devlieghere, F. Mycotoxin Production and Predictive Modelling Kinetics on the Growth of Aspergillus flavus and Aspergillus parasiticus Isolates in Whole Black Peppercorns (Piper nigrum L.). Int. J. Food Microbiol. 2016, 228, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Negash, D. A Review of Aflatoxin: Occurrence, Prevention, and Gapsin Both Food and Feed Safety. J. Appl. Microb. Res. 2018, 1, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Tai, B.; Chang, J.; Liu, Y.; Xing, F. Recent Progress of the Effect of Environmental Factors on Aspergillus flavus Growth and Aflatoxins Production on Foods. Food Qual. Saf. 2020, 4, 21–28. [Google Scholar] [CrossRef]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful Effects and Control Strategies of Aflatoxin B1 Produced by Aspergillus flavus and Aspergillus parasiticus Strains on Poultry: Review. Toxins 2019, 11, 176. [Google Scholar] [CrossRef] [Green Version]
- Gizachew, D.; Chang, C.-H.; Szonyi, B.; De La Torre, S.; Ting, W.E. Aflatoxin B1 (AFB1) Production by Aspergillus flavus and Aspergillus parasiticus on Ground Nyjer Seeds: The Effect of Water Activity and Temperature. Int. J. Food Microbiol. 2019, 296, 8–13. [Google Scholar] [CrossRef]
- Park, K.Y.; Bullerman, L.B. Effect of Cycling Temperatures on Aflatoxin Production by Aspergillus parasiticus and Aspergillus flavus in Rice and Cheddar Cheese. J. Food Sci. 1983, 48, 889–896. [Google Scholar] [CrossRef]
- Yogendrarajah, P.; Devlieghere, F.; Njumbe Ediage, E.; Jacxsens, L.; De Meulenaer, B.; De Saeger, S. Toxigenic Potentiality of Aspergillus flavus and Aspergillus parasiticus Strains Isolated from Black Pepper Assessed by an LC-MS/MS Based Multi-Mycotoxin Method. Food Microbiol. 2015, 52, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Razzaghi-Abyaneh, M.; Shams-Ghahfarokhi, M.; Eslamifar, A.; Schmidt, O.J.; Gharebaghi, R.; Karimian, M.; Naseri, A.; Sheikhi, M. Inhibitory Effects of Akacid®plus on Growth and Aflatoxin Production by Aspergillus parasiticus. Mycopathologia 2006, 161, 245–249. [Google Scholar] [CrossRef]
- Yahyaraeyat, R.; Khosravi, A.R.; Shahbazzadeh, D.; Khalaj, V. The Potential Effects of Zataria multiflora Boiss Essential Oil on Growth, Aflatoxin Production and Transcription of Aflatoxin Biosynthesis Pathway Genes of Toxigenic Aspergillus parasiticus. Braz. J. Microbiol. 2013, 44, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Armando, M.R.; Dogi, C.A.; Rosa, C.A.R.; Dalcero, A.M.; Cavaglieri, L.R. Saccharomyces Cerevisiae Strains and the Reduction of Aspergillus parasiticus Growth and Aflatoxin B1 Production at Different Interacting Environmental Conditions, In Vitro. Food Addit. Contam. Part A 2012, 29, 1443–1449. [Google Scholar] [CrossRef]
- Hugas, M.; Monfort, J.M. Bacterial Starter Cultures for Meat Fermentation. Food Chem. 1997, 59, 547–554. [Google Scholar] [CrossRef]
- González-Fernández, C.; Santos, E.M.; Rovira, J.; Jaime, I. The Effect of Sugar Concentration and Starter Culture on Instrumental and Sensory Textural Properties of Chorizo-Spanish Dry-Cured Sausage. Meat Sci. 2006, 74, 467–475. [Google Scholar] [CrossRef]
- Feiner, G. Salami: Practical Science and Processing Technology; Academic Press: Cambridge, MA, USA, 2016; ISBN 978-0-12-809680-2. [Google Scholar]
- Festring, D.; Hofmann, T. Discovery of N2-(1-Carboxyethyl)Guanosine 5′-Monophosphate as an Umami-Enhancing Maillard-Modified Nucleotide in Yeast Extracts. J. Agric. Food Chem. 2010, 58, 10614–10622. [Google Scholar] [CrossRef]
- Liu, J.; Song, H.; Liu, Y.; Li, P.; Yao, J.; Xiong, J. Discovery of Kokumi Peptide from Yeast Extract by LC-Q-TOF-MS/MS and Sensomics Approach. J. Sci. Food Agric. 2015, 95, 3183–3194. [Google Scholar] [CrossRef]
- Mahadevan, K.; Farmer, L. Key Odor Impact Compounds in Three Yeast Extract Pastes. J. Agric. Food Chem. 2006, 54, 7242–7250. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Patiño, B.; Cortes, L.; Gonzalez-Jaen, M.T.; Vazquez, C. Aspergillus steynii and Aspergillus westerdijkiae as Potential Risk of OTA Contamination in Food Products in Warm Climates. Food Microbiol. 2015, 46, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Alapont, C.; López-Mendoza, M.C.; Gil, J.V.; Martínez-Culebras, P.V. Mycobiota and Toxigenic Penicillium Species on Two Spanish Dry-Cured Ham Manufacturing Plants. Food Addit. Contam. Part A 2014, 31, 93–104. [Google Scholar] [CrossRef]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S.A. Analytical Methods for Determination of Mycotoxins: A Review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Modeling Kinetics of Aflatoxin Production by Aspergillus flavus in Maize-Based Medium and Maize Grain. Int. J. Food Microbiol. 2013, 162, 182–189. [Google Scholar] [CrossRef]
- Le Bars, J. Toxigenesis as a Function of the Ecological Conditions of the Grain/Microorganisms System; Lavoisier: Paris, France, 1988. [Google Scholar]
- Pitt, R.E. A descriptive model of mold growth and aflatoxin formation as affected by environmental conditions. J. Food Prot. 1993, 56, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Yu, J. Current Understanding on Aflatoxin Biosynthesis and Future Perspective in Reducing Aflatoxin Contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honikel, K.-O. The Use and Control of Nitrate and Nitrite for the Processing of Meat Products. Meat Sci. 2008, 78, 68–76. [Google Scholar] [CrossRef] [PubMed]
- López-Díaz, T.-M.; Santos, J.-A.; García-López, M.-L.; Otero, A. Surface Mycoflora of a Spanish Fermented Meat Sausage and Toxigenicity of Penicillium Isolates. Int. J. Food Microbiol. 2001, 68, 69–74. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, H.; Neng, J.; Gao, J.; Yang, B.; Liu, Y. The Influence of NaCl and Glucose Content on Growth and Ochratoxin A Production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum. Toxins 2020, 12, 515. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Medium | λ (Day) | μ (mm/Day) | k AFB1 (ng/mL/Day) | R2 (DIAM/AFB1) | Spearman |
|---|---|---|---|---|---|
| CMA1 | 1.02 a ± 0.04 | 6.11 bc ± 0.60 | 7.82 f ± 2.02 | 0.90 b ± 0.01 | 0.904 |
| CMA2 | 0.82 b ± 0.10 | 10.38 a ± 0.29 | 7.43 f ± 1.55 | 0.63 c ± 0.10 | 0.928 |
| CMA2* | 32.39 e ± 12.37 | ||||
| CMA2G | 0.99 ab ± 0.16 | 10.35 a ± 0.08 | 248.49 d ± 23.08 | 0.79 d ± 0.01 | 0.924 |
| CMA2YE | 0.54 bc ± 0.55 | 7.64 b ± 0.57 | 1.76 g ± 0.31 | 0.77 c ± 0.10 | 0.795 |
| CMA2GYE | 0.67 b ± 0.60 | 7.37 b ± 0.07 | 629.07 b ± 36.86 | 0.93 a ± 0.01 | 0.979 |
| CMA2GYE* | 2147.63 a ± 780.32 | ||||
| YES | 0.45 c ± 0.18 | 10.03 a ± 1.30 | 2895.00 a ± 368.42 | 0.91 b ± 0.02 | 0.895 |
| MEA | 0.36 c ± 0.35 | 5.42 c ± 0.42 | 424.23 c ± 15.43 | 0.93 a ± 0.01 | 0.895 |
| CYA | 0.81 ab ± 0.62 | 8.43 b ± 0.59 | 663.30 b ± 68.17 | 0.93 a ± 0.05 | 0.912 |
| Kruskal- Wallis test (p) | 0.032 | 0.004 | 0.002 | 0.021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahija, I.; Jeršek, B.; Demšar, L.; Polak, M.L.; Polak, T. Production of Aflatoxin B1 by Aspergillus parasiticus Grown on a Novel Meat-Based Media. Toxins 2023, 15, 25. https://doi.org/10.3390/toxins15010025
Zahija I, Jeršek B, Demšar L, Polak ML, Polak T. Production of Aflatoxin B1 by Aspergillus parasiticus Grown on a Novel Meat-Based Media. Toxins. 2023; 15(1):25. https://doi.org/10.3390/toxins15010025
Chicago/Turabian StyleZahija, Iva, Barbka Jeršek, Lea Demšar, Mateja Lušnic Polak, and Tomaž Polak. 2023. "Production of Aflatoxin B1 by Aspergillus parasiticus Grown on a Novel Meat-Based Media" Toxins 15, no. 1: 25. https://doi.org/10.3390/toxins15010025