Arabic Gum Could Alleviate the Aflatoxin B1-provoked Hepatic Injury in Rat: The Involvement of Oxidative Stress, Inflammatory, and Apoptotic Pathways

 , , ,
, , ,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
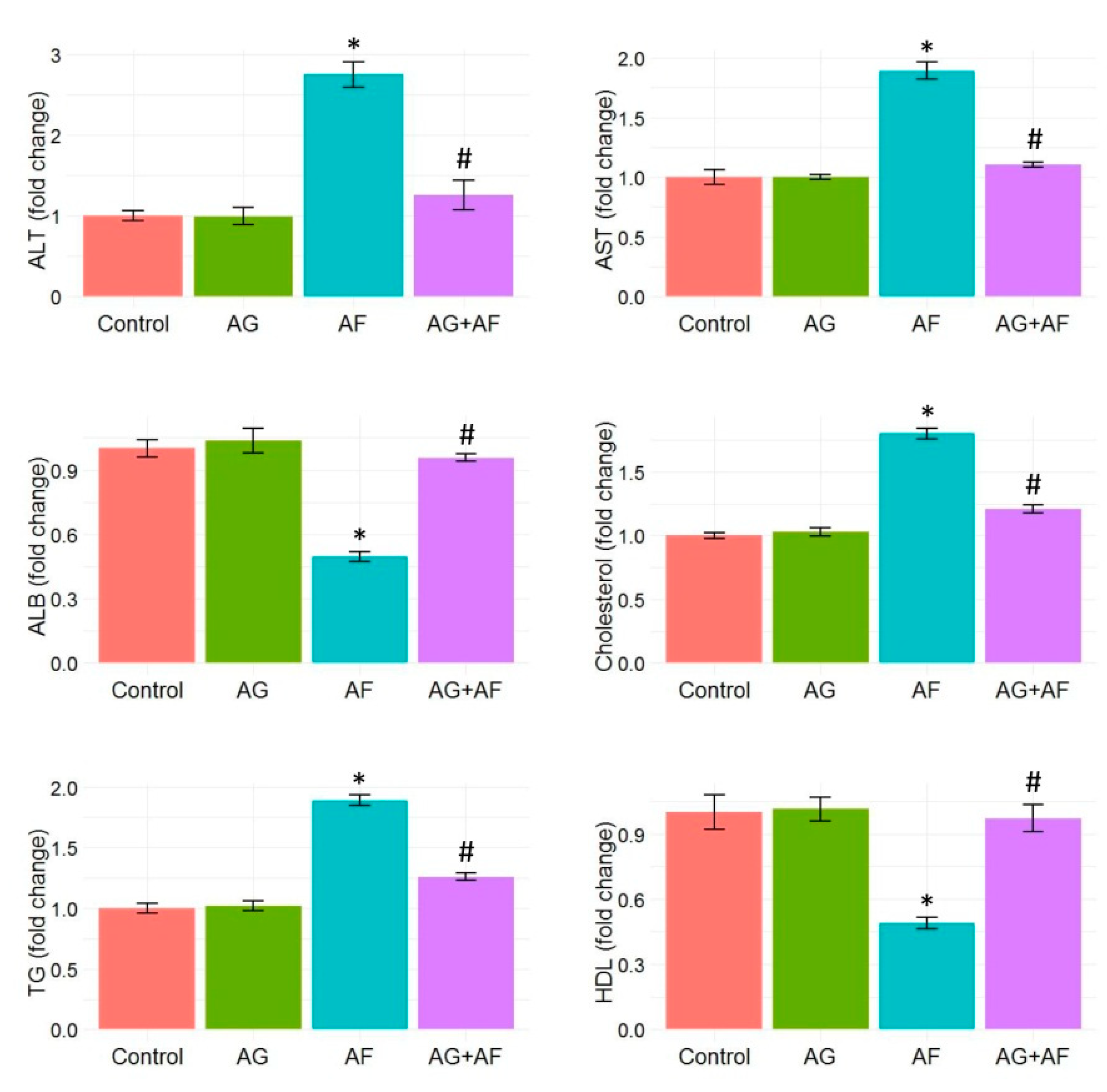
2.1. Biochemical Parameters Screening
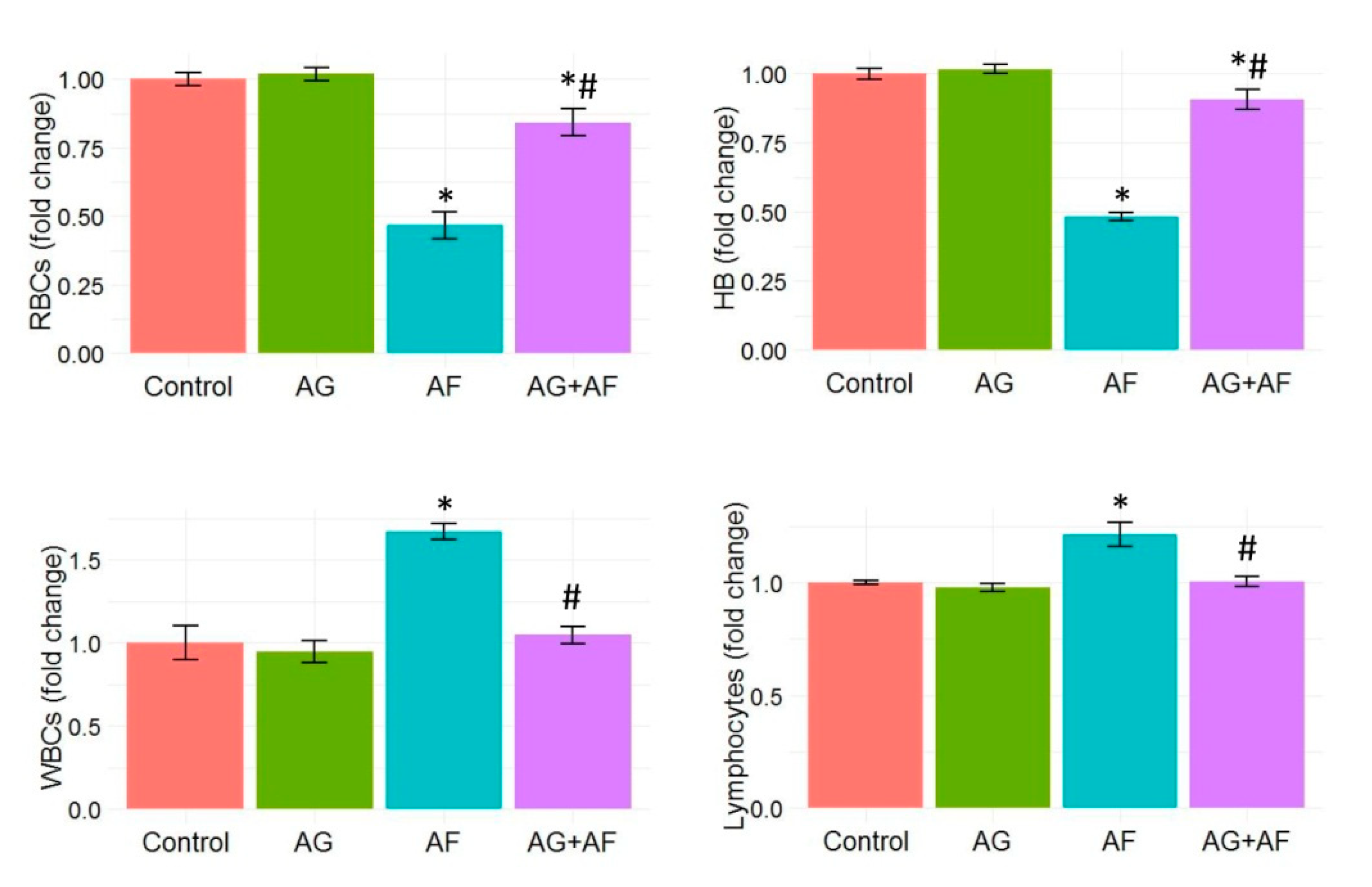
2.2. Hematological Profile
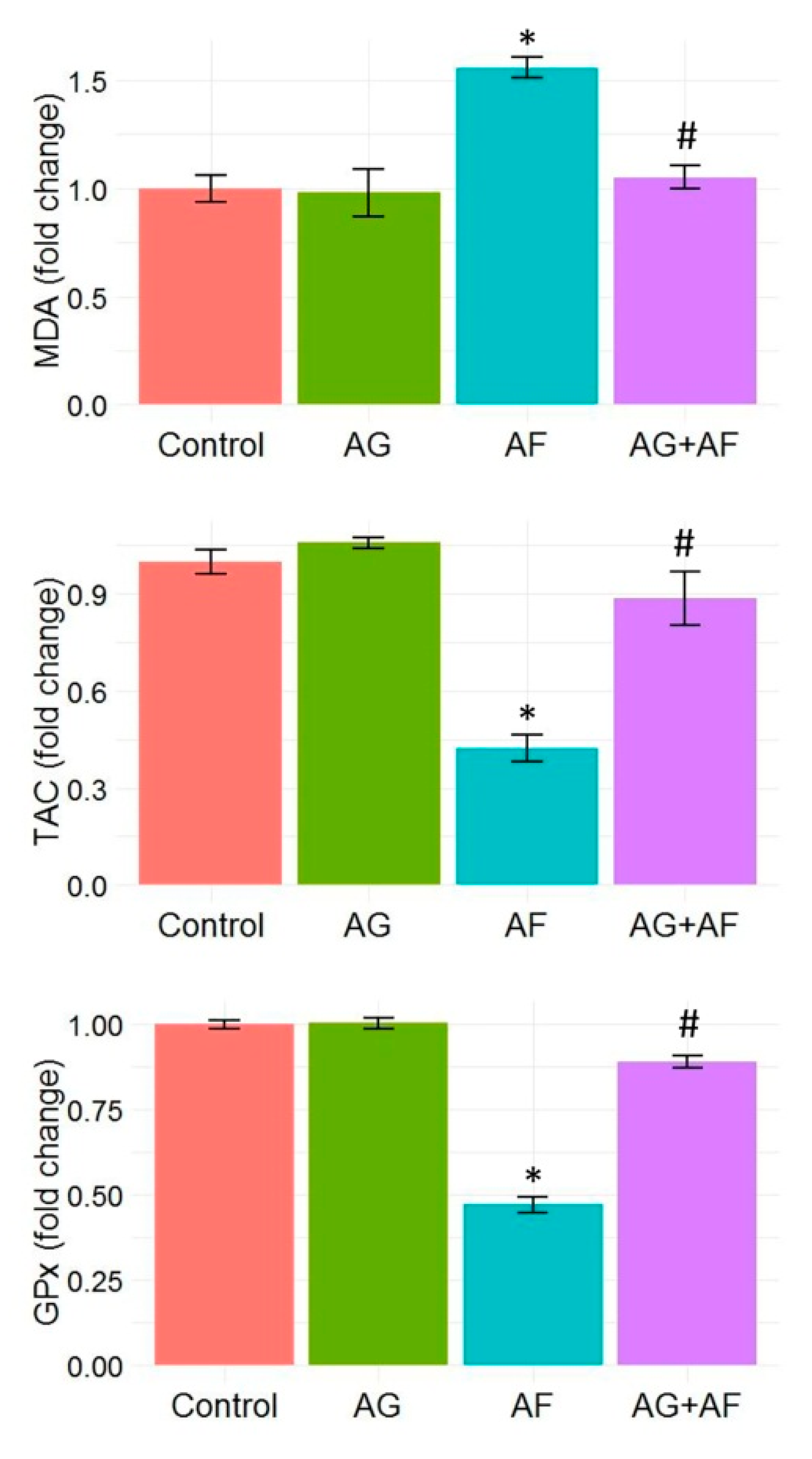
2.3. Cellular Antioxidants and Lipid Peroxidation Indices
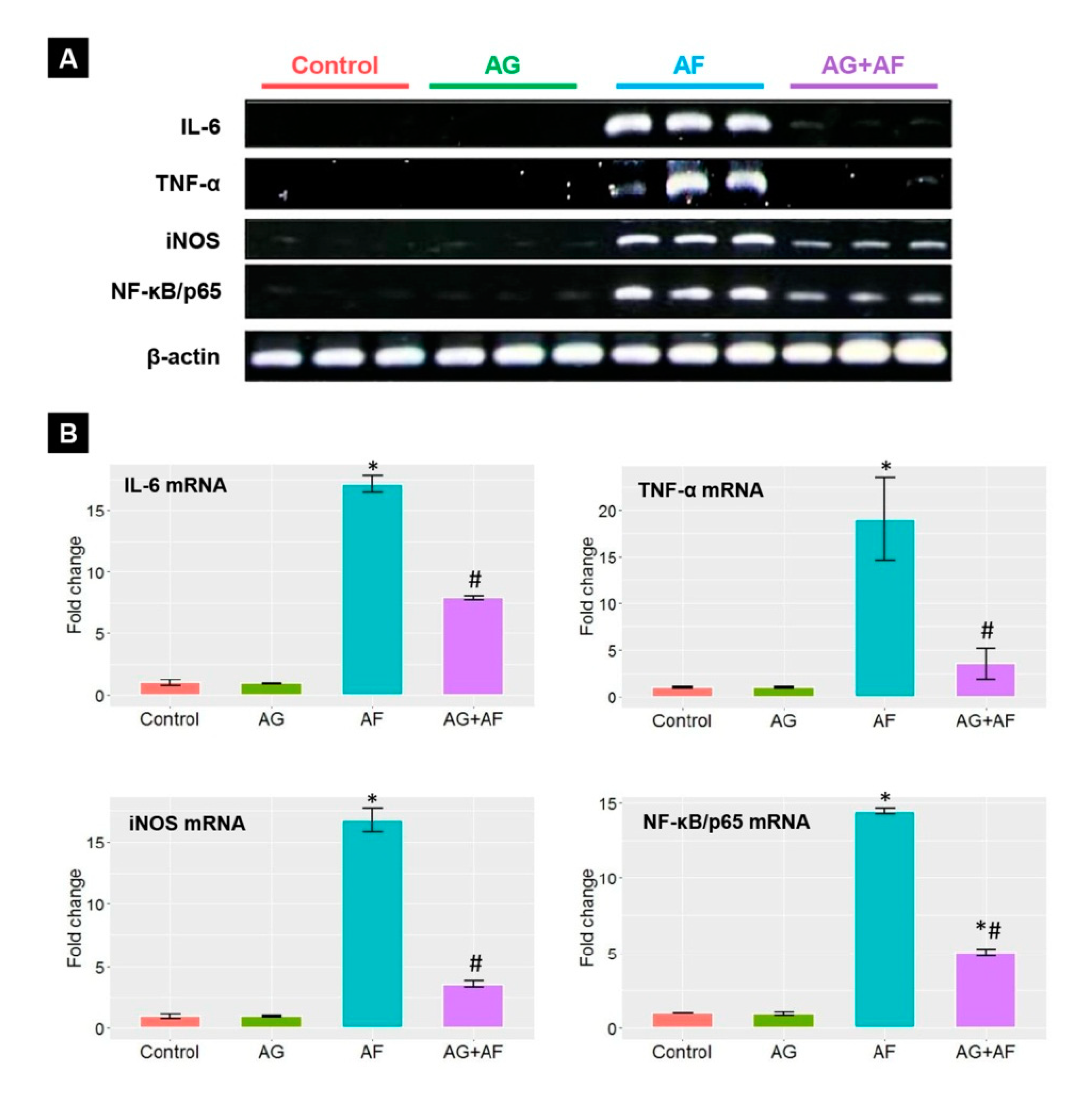
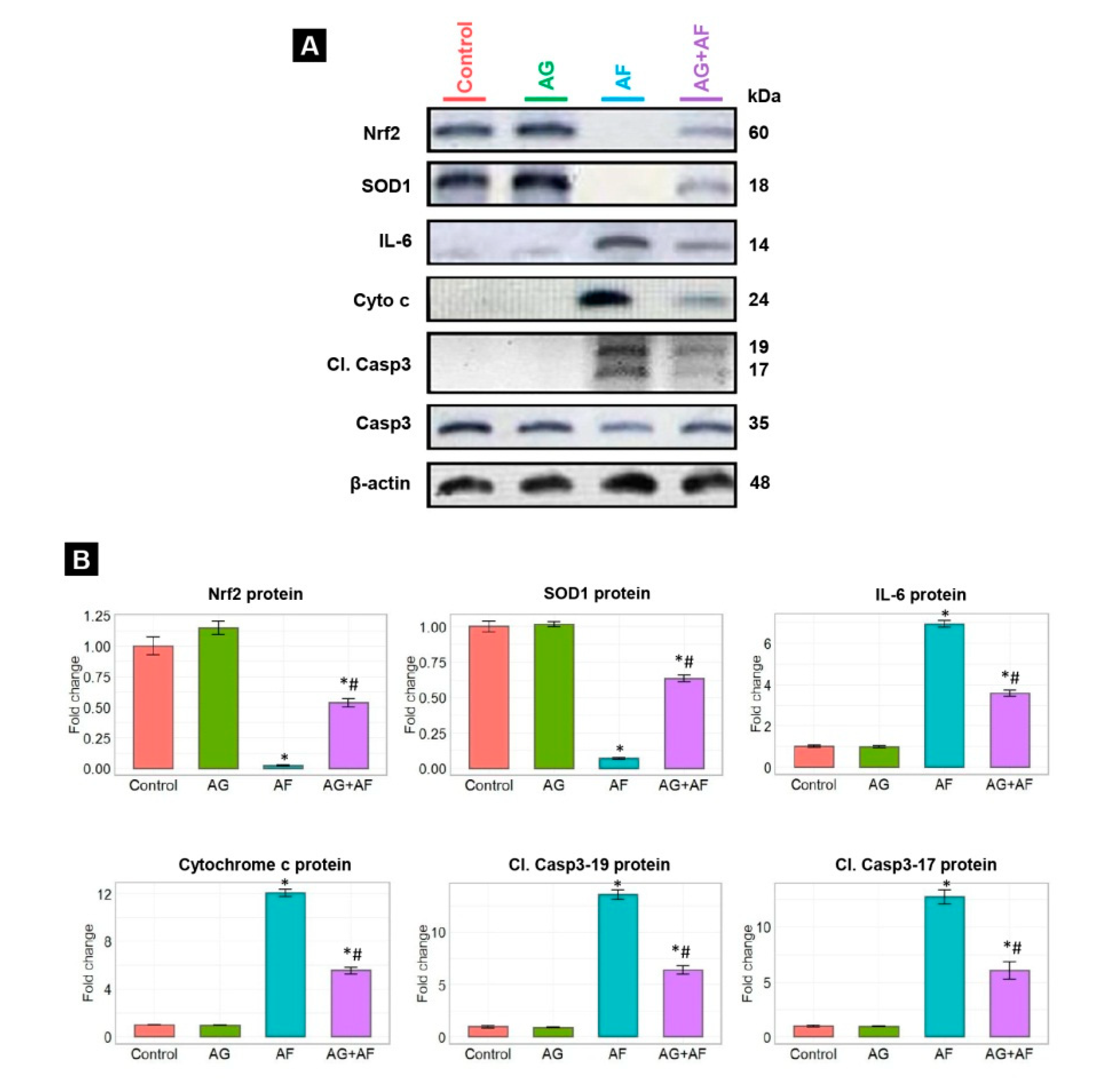
2.4. Proinflammatory Cytokine Expression
2.5. Apoptotic Biomarker Protein Expression
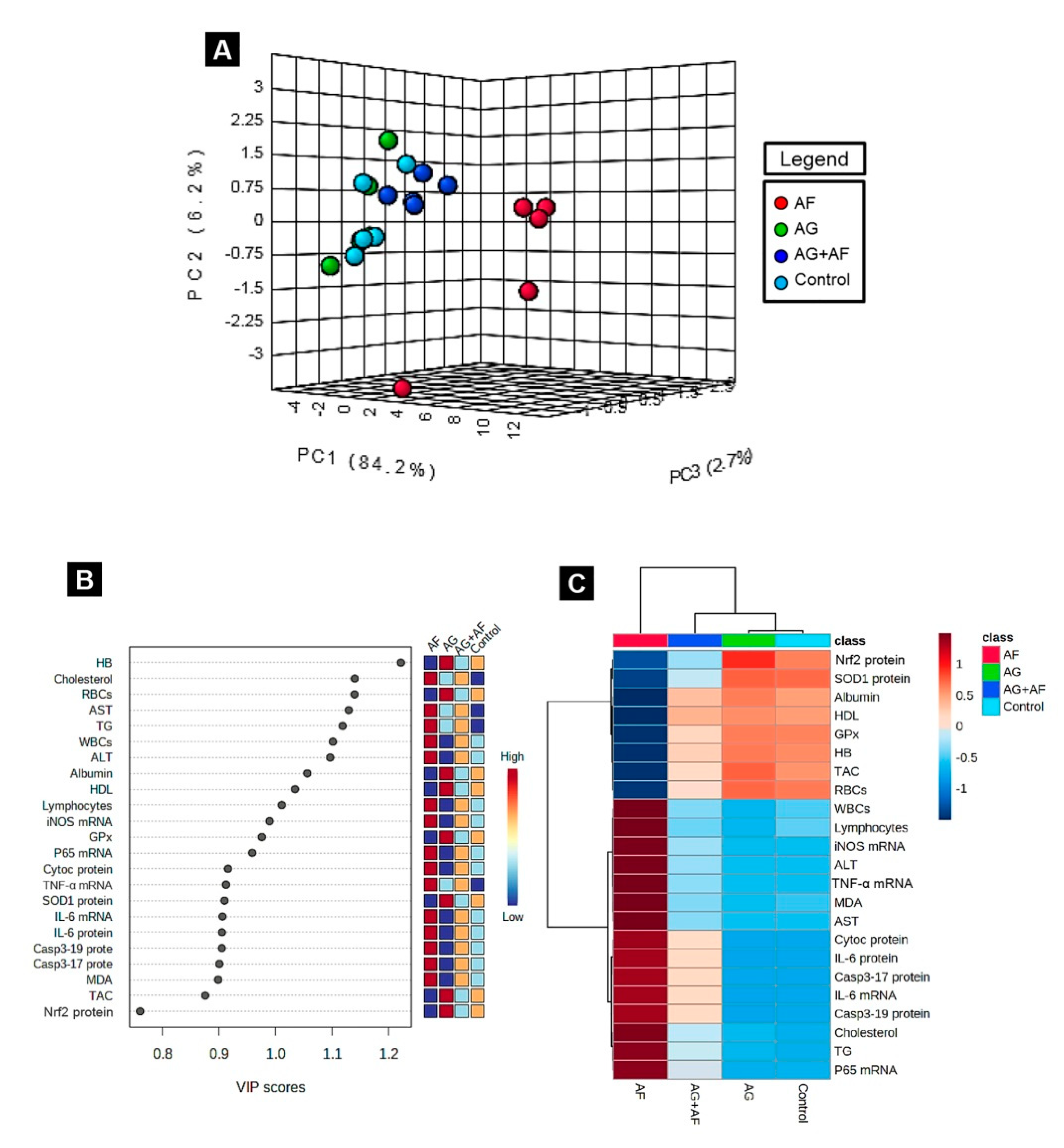
2.6. Principal Component Analysis (PCA), Variable Importance in Projection (VIP) Score, and Hierarchical Clustering Heatmap
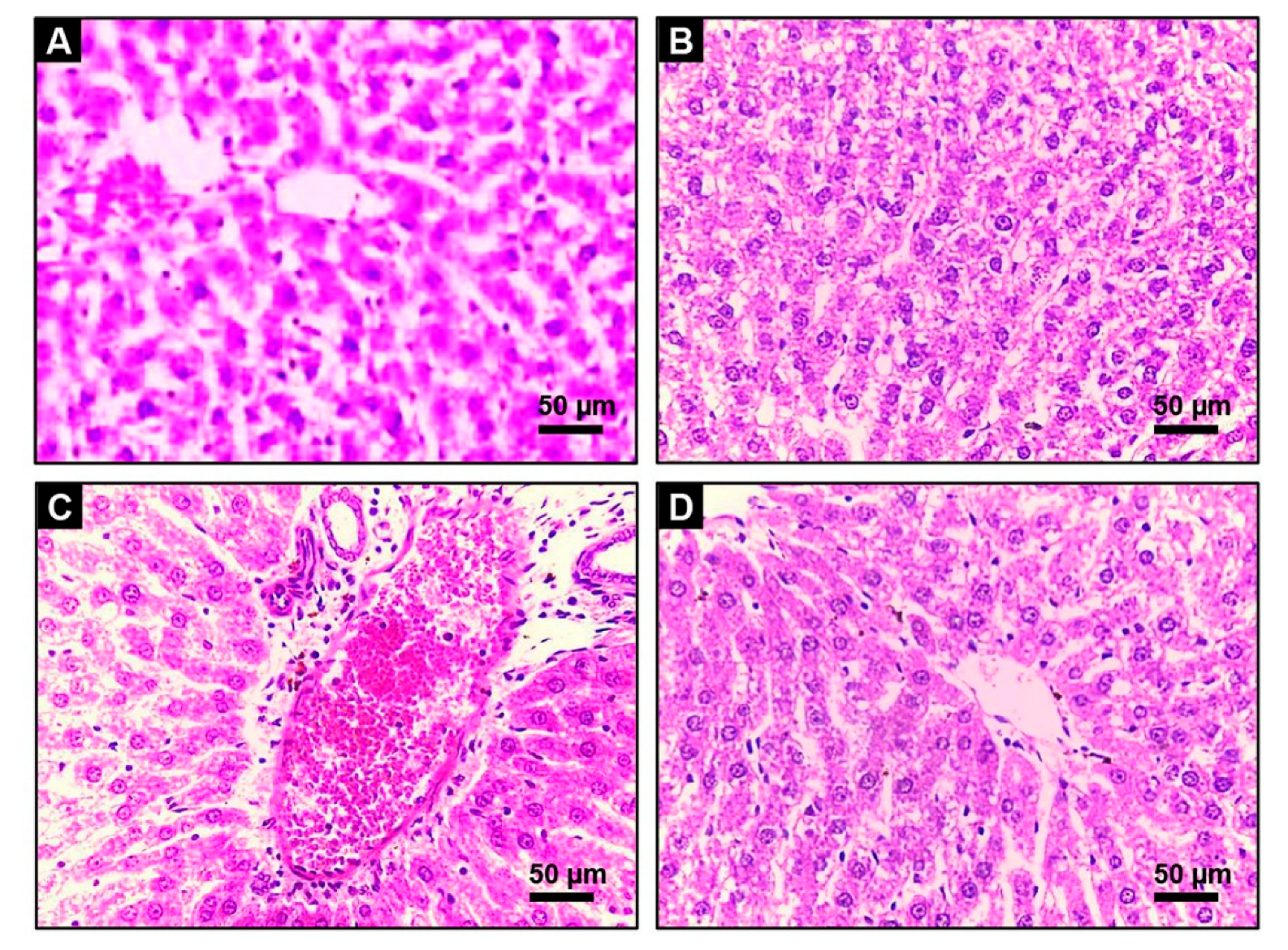
2.7. Hepatic Histoarchitecture Inspection
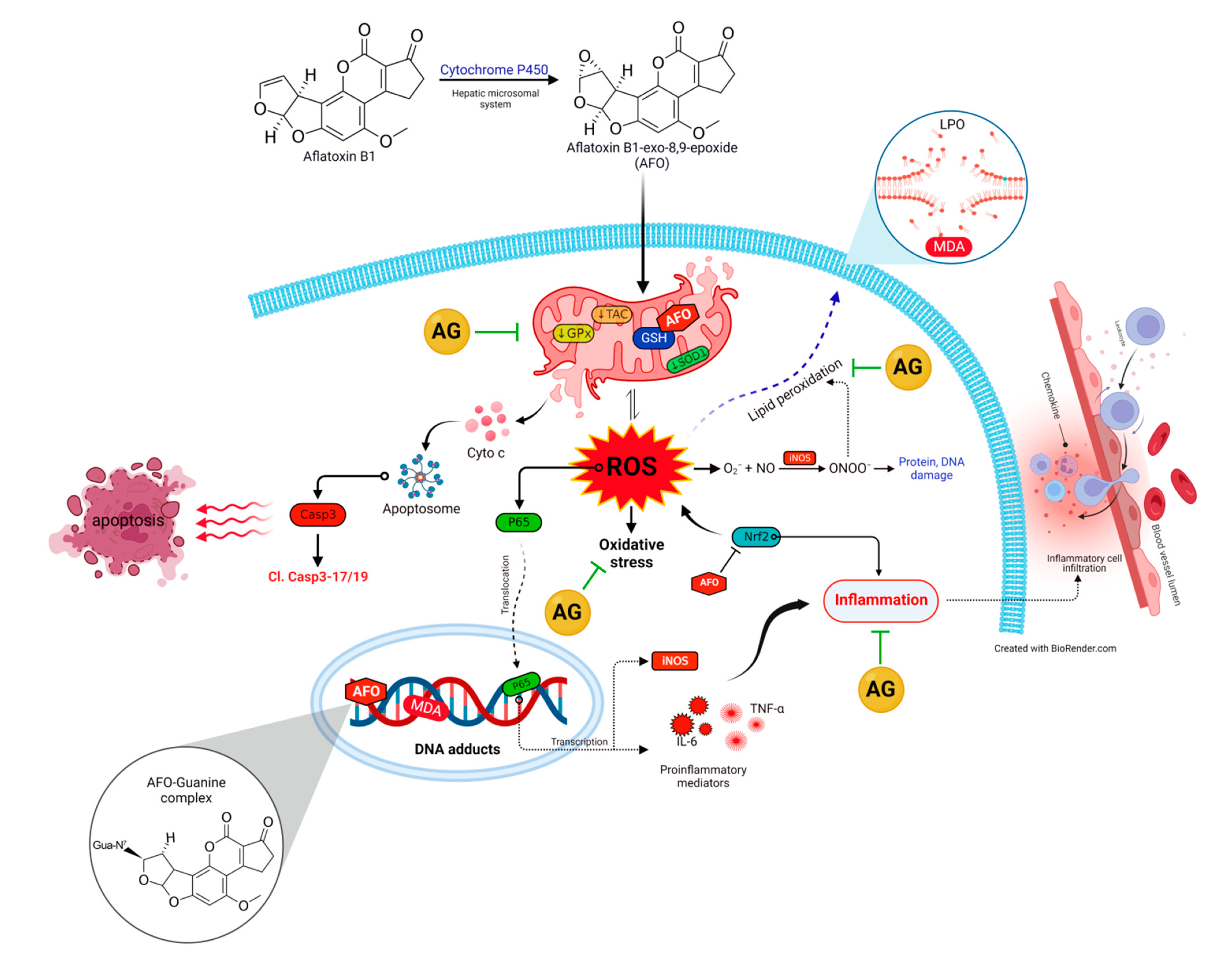
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Protocol of Trial
5.2. Specimens Collection and Processing
5.3. Biochemical Parameters Bioassay and Hematological Profile
5.4. Antioxidants and Peroxidation Biomarkers
5.5. Reverse Transcription-PCR
5.6. Western Blotting
5.7. Histoarchitectures Assessment
5.8. Statistical Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Zhu, F.; Zhao, X.; Li, J.; Guo, L.; Bai, L.; Qi, X. A new compound Trichomicin exerts antitumor activity through STAT3 signaling inhibition. Biomed. Pharmacother. 2020, 121, 109608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jiao, P.; Wang, X.; Sun, Y.; Liang, G.; Xie, X.; Zhang, Y. Evaluation of Growth Performance, Nitrogen Balance and Blood Metabolites of Mutton Sheep Fed an Ammonia-Treated Aflatoxin B1-Contaminated Diet. Toxins 2022, 14, 361. [Google Scholar] [CrossRef] [PubMed]
- COMMISSION REGULATION (EC) No 401/2006 Laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Comm. Regul. 2006, 70, 12.
- Abdel-Daim, M.M.; Abdeen, A.; Jalouli, M.; Abdelkader, A.; Megahed, A.; Alkahtane, A.; Almeer, R.; Alhoshani, N.M.; Al-Johani, N.S.; Alkahtani, S.; et al. Fucoidan supplementation modulates hepato-renal oxidative stress and DNA damage induced by aflatoxin B1 intoxication in rats. Sci. Total Environ. 2021, 768, 144781. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, S.; Bag, H. Aflatoxin B1: Mechanism, oxidative stress, and effects on animal health. Insights Vet. Sci. 2022, 6, 17–24. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Xu, F.; Huang, W.; Ji, Q.; Han, Y.; Shao, B.; Li, Y. Protective effects of lycopene against AFB1-induced erythrocyte dysfunction and oxidative stress in mice. Res. Vet. Sci. 2020, 129, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef]
- Karamkhani, M.; Asilian-Mahabadi, H.; Daraei, B.; Seidkhani-Nahal, A.; Noori-Zadeh, A. Liver and kidney serum profile abnormalities in workers exposed to aflatoxin B1 in urban solid waste management centers. Environ. Monit. Assess. 2020, 192, 472. [Google Scholar] [CrossRef]
- Ma, J.; Liu, Y.; Guo, Y.; Ma, Q.; Ji, C.; Zhao, L. Transcriptional profiling of aflatoxin b1-induced oxidative stress and inflammatory response in macrophages. Toxins 2021, 13, 401. [Google Scholar] [CrossRef]
- Aleissa, M.S.; Alkahtani, S.; Abd Eldaim, M.A.; Ahmed, A.M.; Bungǎu, S.G.; Almutairi, B.; Bin-Jumah, M.; Alkahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M.; et al. Fucoidan Ameliorates Oxidative Stress, Inflammation, DNA Damage, and Hepatorenal Injuries in Diabetic Rats Intoxicated with Aflatoxin B 1. Oxid. Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef]
- Yilmaz, S.; Kaya, E.; Kisacam, M.A. The Effect on Oxidative Stress of Aflatoxin and Protective Effect of Lycopene on Aflatoxin Damage. Aflatoxin-Control Anal. Detect. Health Risks 2017, 30, 67–90. [Google Scholar] [CrossRef] [Green Version]
- Omar, M.; El-Din, H. Mycotoxins-Induced Oxidative Stress and Disease. Mycotoxin Food Saf. Dev. Ctries. 2013, 2013, 63–92. [Google Scholar] [CrossRef]
- Bbosa, G.S.; Kitya, D.; Odda, J.; Ogwal-Okeng, J. Aflatoxins metabolism, effects on epigenetic mechanisms and their role in carcinogenesis. Health 2013, 5, 14–34. [Google Scholar] [CrossRef]
- Abdel-Daim, M.; Dawood, M.A.O.; AlKahtane, A.A.; Abdeen, A.; Abdel-Latif, H.M.R.; Senousy, H.H.; Aleya, L.; Alkahtani, S. Spirulina platensis mediated the biochemical indices and antioxidative function of Nile tilapia (Oreochromis niloticus) intoxicated with aflatoxin B1. Toxicon 2020, 184, 152–157. [Google Scholar] [CrossRef]
- Li, S.; Liu, R.; Xia, S.; Wei, G.; Ishfaq, M.; Zhang, Y.; Zhang, X. Protective role of curcumin on aflatoxin B1-induced TLR4/RIPK pathway mediated-necroptosis and inflammation in chicken liver. Ecotoxicol. Environ. Saf. 2022, 233, 113319. [Google Scholar] [CrossRef]
- Owumi, S.; Najophe, E.S.; Farombi, E.O.; Oyelere, A.K. Gallic acid protects against Aflatoxin B1-induced oxidative and inflammatory. J. Food Biochem. 2020, 44, e13316. [Google Scholar] [CrossRef]
- Xu, D.; Peng, S.; Guo, R.; Yao, L.; Mo, H.; Li, H.; Song, H.; Hu, L. Egcg alleviates oxidative stress and inhibits aflatoxin b1 biosynthesis via mapk signaling pathway. Toxins 2021, 13, 693. [Google Scholar] [CrossRef]
- Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Dietary curcumin alleviated acute ileum damage of ducks (Anas platyrhynchos) induced by afb1 through regulating nrf2-are and nf-κb signaling pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef]
- Hassanien, M.A. The protective and antioxidant effects of gum arabic: A review of recent evidence using the new PubMed system. Int. J. Community Med. Public Health 2019, 7, 356. [Google Scholar] [CrossRef]
- Elshama, S. The preventive role of Arabic gum in the treatment of Toxicity. Open Access Toxicol. Res. 2018, 1, 27–29. [Google Scholar]
- Ali, M.R.; EL Said, R.M. Assessment of the potential of Arabic gum as an antimicrobial and antioxidant agent in developing vegan “egg-free” mayonnaise. J. Food Saf. 2020, 40, e12771. [Google Scholar] [CrossRef]
- Mirghani, M.E.S.; Elnour, A.A.M.; Kabbashi, N.A.; Alam, M.Z.; Musa, K.H.; Abdullah, A. Determination of antioxidant activity of gum Arabic: An exudation from two different locations. Sci. Asia 2018, 44, 179–186. [Google Scholar] [CrossRef]
- Kamal, E.; Kaddam, L.A.; Dahawi, M.; Osman, M.; Salih, M.A.; Alagib, A.; Saeed, A. Gum Arabic fibers decreased inflammatory markers and disease severity score among rheumatoid arthritis patients, phase II Trial. Int. J. Rheumatol. 2018, 2018, 4197537. [Google Scholar] [CrossRef] [PubMed]
- Musa, H.H.; Ahmed, A.A.; Musa, T.H. Chemistry, Biological, and Pharmacological Properties of Gum Arabic. Ref. Ser. Phytochem. 2019, 2019, 797–814. [Google Scholar] [CrossRef]
- Avelino, A.L.N.; Silva, N.V.R.e.; Oliveira, G.B.D.; Silva, A.A.D.S.; Cavalcanti, B.C.; Jamacaru, F.V.F.; Dornelas, C.A. Antioxidant and Antigenotoxic Actions of Gum Arabic on the Intestinal Mucosa, Liver and Bone Marrow of Swiss Mice Submitted to Colorectal Carcinogenesis. Nutr. Cancer 2022, 74, 956–964. [Google Scholar] [CrossRef]
- Ali, N.E.; Kaddam, L.A.; Alkarib, S.Y.; Kaballo, B.G.; Khalid, S.A.; Higawee, A.; Abdelhabib, A.; Alaaaldeen, A.; Phillips, A.O.; Saeed, A.M. Gum Arabic (Acacia Senegal) Augmented Total Antioxidant Capacity and Reduced C-Reactive Protein among Haemodialysis Patients in Phase II Trial. Int. J. Nephrol. 2020, 2020, 7214673. [Google Scholar] [CrossRef]
- Eslick, S.; Thompson, C.; Berthon, B.; Wood, L. Short-chain fatty acids as anti-inflammatory agents in overweight and obesity: A systematic review and meta-analysis. Nutr. Rev. 2022, 80, 838–856. [Google Scholar] [CrossRef]
- Abu-Serie, M.M.; Hamouda, A.F.; Habashy, N.H. Acacia senegal gum attenuates systemic toxicity in CCl4-intoxicated rats via regulation of the ROS/NF-κB signaling pathway. Sci. Rep. 2021, 11, 20316. [Google Scholar] [CrossRef]
- Al-Kenanny, E.R.; Al-Hayaly, L.K.; Al-Badrany, A.G. Protective Effect of Arabic Gum on liver Injury Experimentally Induced by Gentamycin in Mice Kufa. J. Vet. Med. Sci. 2012, 3, 174–189. [Google Scholar] [CrossRef]
- Gamal El-din, A.M.; Mostafa, A.M.; Al-Shabanah, O.A.; Al-Bekairi, A.M.; Nagi, M.N. Protective effect of arabic gum against acetaminophen-induced hepatotoxicity in mice. Pharmacol. Res. 2003, 48, 631–635. [Google Scholar] [CrossRef]
- Cao, Q.Q.; Lin, L.X.; Xu, T.T.; Lu, Y.; Zhang, C.D.; Yue, K.; Huang, S.C.; Dong, H.J.; Jian, F.C. Aflatoxin B1 alters meat quality associated with oxidative stress, inflammation, and gut-microbiota in sheep. Ecotoxicol. Environ. Saf. 2021, 225, 112754. [Google Scholar] [CrossRef] [PubMed]
- Rotimi, O.A.; Rotimi, S.O.; Goodrich, J.M.; Adelani, I.B.; Agbonihale, E.; Talabi, G. Time-course effects of acute aflatoxin B1 exposure on hepatic mitochondrial lipids and oxidative stress in rats. Front. Pharmacol. 2019, 10, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Yan, W.R.; Tang, J.K.; Jin, X.; Xue, H.H.; Wang, T.; Sun, Q.Y.; Liang, Z.X.; Zhang, L.W. Dietary phillygenin supplementation ameliorates aflatoxin B1-induced oxidative stress, inflammation, and apoptosis in chicken liver. Ecotoxicol. Environ. Saf. 2022, 236, 113481. [Google Scholar] [CrossRef]
- Hussain, S.P.; Amstad, P.; He, P.; Robles, A.; Lupold, S.; Kaneko, I.; Ichimiya, M.; Sengupta, S.; Mechanic, L.; Okamura, S.; et al. p53-Induced Up-Regulation of MnSOD and GPx but not Catalase Increases Oxidative Stress and Apoptosis. Cancer Res. 2004, 64, 2350–2356. [Google Scholar] [CrossRef]
- Gao, X.; Xu, J.; Jiang, L.; Liu, W.; Hong, H.; Qian, Y.; Li, S.; Huang, W.; Zhao, H.; Yang, Z.; et al. Morin alleviates aflatoxin B1-induced liver and kidney injury by inhibiting heterophil extracellular traps release, oxidative stress and inflammatory responses in chicks. Poult. Sci. 2021, 100, 101513. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Hua, H.; Sheng, J.; Cui, Y.; Zhang, W.; Hu, B.; Cheng, Y.; Guo, Y.; Qian, H. The intervention and mechanism of action for aloin against subchronic aflatoxin b1 induced hepatic injury in rats. Int. J. Mol. Sci. 2021, 22, 11620. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Tian, L.; Liu, F.; Jin, S.; Liang, G.; Yang, H. Toxic effects and residue of a fl atoxin B1 in tilapia (Oreochromis niloticus × O. aureus) during long-term dietary exposure. Aquaculture 2010, 307, 233–240. [Google Scholar] [CrossRef]
- Dónmez, N.; Dónmez, H.H.; Keskin, E.; Kisadere, I. Effects of aflatoxin on some haematological parameters and protective effectiveness of esterified glucomannan in merino rams. Sci. World J. 2012, 2012, 342468. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Nada, S.A.; Khalil, F.A. Physiological and toxicological responses in rats fed aflatoxin-contaminated diet with or without sorbent materials. Anim. Feed Sci. Technol. 2002, 97, 209–219. [Google Scholar] [CrossRef]
- Marin, D.E.; Bulgaru, C.V.; Anghel, C.A.; Pistol, G.C.; Dore, M.I.; Palade, M.L.; Taranu, I. Grape Seed Waste Counteracts Aflatoxin B1 Toxicity in Piglet Mesenteric Lymph Nodes. Toxins 2020, 12, 800. [Google Scholar] [CrossRef] [PubMed]
- Arbab, M.; Tahir, S.; Niazi, M.K.; Ishaq, M.; Hussain, A.; Siddique, P.M.; Saeed, S.; Khan, W.A.; Qamar, R.; Butt, A.M.; et al. TNF-α genetic predisposition and higher expression of inflammatory pathway components in keratoconus. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3481–3487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Dai, Y.; Zhu, L.; He, X.; Huang, K.; Xu, W. Single-cell sequencing reveals novel mechanisms of Aflatoxin B1-induced hepatotoxicity in S phase-arrested L02 cells. Cell Biol. Toxicol. 2020, 36, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.K.; Chang, S.N.; Kang, S.C. The inflammation response and risk associated with aflatoxin B1 contamination was minimized by insect peptide CopA3 treatment and act towards the beneficial health outcomes. Environ. Pollut. 2021, 268, 115713. [Google Scholar] [CrossRef] [PubMed]
- Cruceriu, D.; Baldasici, O.; Balacescu, O.; Berindan-Neagoe, I. The dual role of tumor necrosis factor-alpha (TNF-α) in breast cancer: Molecular insights and therapeutic approaches. Cell. Oncol. 2020, 43, 1–18. [Google Scholar] [CrossRef]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The role of tumor necrosis factor alpha (Tnf-α) in autoimmune disease and current tnf-α inhibitors in therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef]
- Benkerroum, N. Chronic and acute toxicities of aflatoxins: Mechanisms of action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Abdeen, A.; Samir, A.; Elkomy, A.; Aboubaker, M.; Habotta, O.A.; Gaber, A.; Alsanie, W.F.; Abdullah, O.; Elnoury, H.A.; Baioumy, B.; et al. The potential antioxidant bioactivity of date palm fruit against gentamicin-mediated hepato-renal injury in male albino rats. Biomed. Pharmacother. 2021, 143, 112154. [Google Scholar] [CrossRef]
- Owumi, S.E.; Kazeem, A.I.; Wu, B.; Ishokare, L.O.; Arunsi, U.O.; Oyelere, A.K. Apigeninidin-rich Sorghum bicolor (L. Moench) extracts suppress A549 cells proliferation and ameliorate toxicity of aflatoxin B1-mediated liver and kidney derangement in rats. Sci. Rep. 2022, 12, 7438 . [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.N.; Korge, P.; Honda, H.M.; Ping, P. Role of the mitochondrial permeability transition in myocardial disease. Circ. Res. 2003, 93, 292–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Li, Y.; Cao, Z.; Zhang, J.; Huang, W. AFB1-induced mice liver injury involves mitochondrial dysfunction mediated by mitochondrial biogenesis inhibition. Ecotoxicol. Environ. Saf. 2021, 216, 112213. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, Y.; Matsuda, E.; Nishii, T.; Ishida, Y.; Kawaichi, M. Down-regulation of CIBZ, a novel substrate of caspase-3, induces apoptosis. J. Biol. Chem. 2008, 283, 14242–14247. [Google Scholar] [CrossRef]
- Meki, A.-R.M.A.; Abdel-Ghaffar, S.K.; El-Gibaly, I. Afl atoxin B1 Induces Apoptosis in Rat Liver: Protective Effect of Melatonin Article. Neuroendocrinol. Lett. 2001, 22, 417–426. [Google Scholar]
- Omer, S.A.; Badi, R.M.; Garelnabi, M.E.; Gafar, R.A.; Saeed, A.M. Gum Arabic-induced oral photoprotection: Shifting the balance against circulating immune suppressive cytokines. Sci. Afr. 2021, 14, e00973. [Google Scholar] [CrossRef]
- Mahmoud, M.F.; Diaai, A.A.; Ahmed, F. Evaluation of the efficacy of Ginger, Arabic gum, and Boswellia in acute and chronic renal failure. Ren. Fail. 2012, 34, 73–82. [Google Scholar] [CrossRef]
- Sanford, J.A.; O’Neill, A.M.; Zouboulis, C.C.; Gallo, R.L. Short-Chain Fatty Acids from Cutibacterium acnes Activate Both a Canonical and Epigenetic Inflammatory Response in Human Sebocytes. J. Immunol. 2019, 202, 1767–1776. [Google Scholar] [CrossRef]
- Hamid, M.; Abdulrahim, Y.; Abdelnasir, A.; Mohammedsalih, K.M.; Omer, N.A. Protective Effect of Gum Arabic on Liver Oxidative Stress, Inflammation and Apoptosis Induced by CCl 4 in vivo. EAS J. Nurs. Midwifery 2021, 3, 27–34. [Google Scholar] [CrossRef]
- Al-Majed, A.A.; Abd-Allah, A.R.A.; Al-Rikabi, A.C.; Al-Shabanah, O.A.; Mostafa, A.M. Effect of oral administration of arabic gum on cisplatin-induced Nephrotoxicity in Rats. J. Biochem. Mol. Toxicol. 2003, 17, 146–153. [Google Scholar] [CrossRef]
- Tang, L.; Guan, H.; Ding, X.; Wang, J.S. Modulation of aflatoxin toxicity and biomarkers by lycopene in F344 rats. Toxicol. Appl. Pharmacol. 2007, 219, 10–17. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward (from 5′ to 3′) | Reverse (from 3′ to 5′) | Accession No. |
|---|---|---|---|
| NF-κB/p65 | GACGAGGCTCGGAGAGCCCA | CTGGGGCGGCTGACCGAATG | NM_001029913.1 |
| iNOS | CACCACCCTCCTTGTTCAAC | CAATCCACAACTCGCTCCAA | NM_012611.3 |
| IL-6 | TCCTACCCCAACTTCCAATGCTC | TTGGATGGTCTTGGTCCTTAGCC | NM_012589.2 |
| TNF-α | AAATGGGCTCCCTCTCATCAGTTC | TCTGCTTGGTGGTTTGCTACGAC | X66539.1 |
| β-actin | AGGCACCAGGGTGTGAT | ATGTCACGCACGATTTCC | NM_031144.3 |
| Antibody | Manufacturer | Catalog No. | Clone No. | Dilution Factor |
|---|---|---|---|---|
| Nrf2 | SANTA CRUZ | Sc-518036 | H-10 | 1/1000 |
| SOD1 | SANTA CRUZ | Sc-101523 | 24 | 1/1000 |
| Cytochrome c | Cell signaling | 12963S | 6H2.B4 | 1/1000 |
| IL-6 | Cell signaling | 12912 | D5W4V | 1/1000 |
| Caspase3 | Cell signaling | #9662 | 1/1000 | |
| Cleaved-Caspase3 (Asp175) | Cell signaling | #9664 | 5A1E | 1/1000 |
| β-actin | Sigma-Aldrich | A5441 | AC-15 | 1/1000 |
| Goat anti-rat IgG/HPR | Abcam | ab205720 | 1/1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, N.; El-Rayes, S.M.; Khalil, W.F.; Abdeen, A.; Abdelkader, A.; Youssef, M.; Maher, Z.M.; Ibrahim, A.N.; Abdelrahman, S.M.; Ibrahim, S.F.; et al. Arabic Gum Could Alleviate the Aflatoxin B1-provoked Hepatic Injury in Rat: The Involvement of Oxidative Stress, Inflammatory, and Apoptotic Pathways. Toxins 2022, 14, 605. https://doi.org/10.3390/toxins14090605
Ahmed N, El-Rayes SM, Khalil WF, Abdeen A, Abdelkader A, Youssef M, Maher ZM, Ibrahim AN, Abdelrahman SM, Ibrahim SF, et al. Arabic Gum Could Alleviate the Aflatoxin B1-provoked Hepatic Injury in Rat: The Involvement of Oxidative Stress, Inflammatory, and Apoptotic Pathways. Toxins. 2022; 14(9):605. https://doi.org/10.3390/toxins14090605
Chicago/Turabian StyleAhmed, Noha, Samir M. El-Rayes, Waleed F. Khalil, Ahmed Abdeen, Afaf Abdelkader, Mohammed Youssef, Zainab M. Maher, Amany N. Ibrahim, Shaymaa M. Abdelrahman, Samah F. Ibrahim, and et al. 2022. "Arabic Gum Could Alleviate the Aflatoxin B1-provoked Hepatic Injury in Rat: The Involvement of Oxidative Stress, Inflammatory, and Apoptotic Pathways" Toxins 14, no. 9: 605. https://doi.org/10.3390/toxins14090605