GTPase Rac Regulates Conidiation, AFB1 Production and Stress Response in Pathogenic Fungus Aspergillus flavus


Abstract
:1. Introduction
2. Result
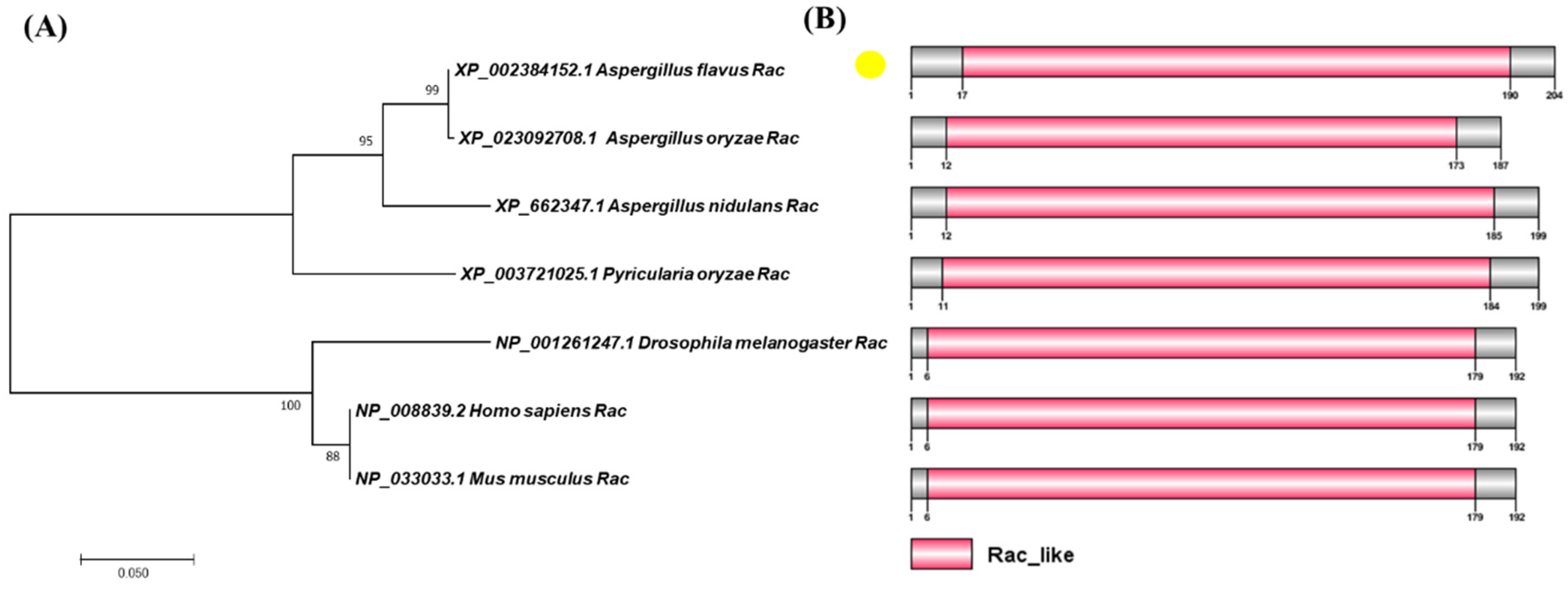
2.1. Biological Analysis of Rac
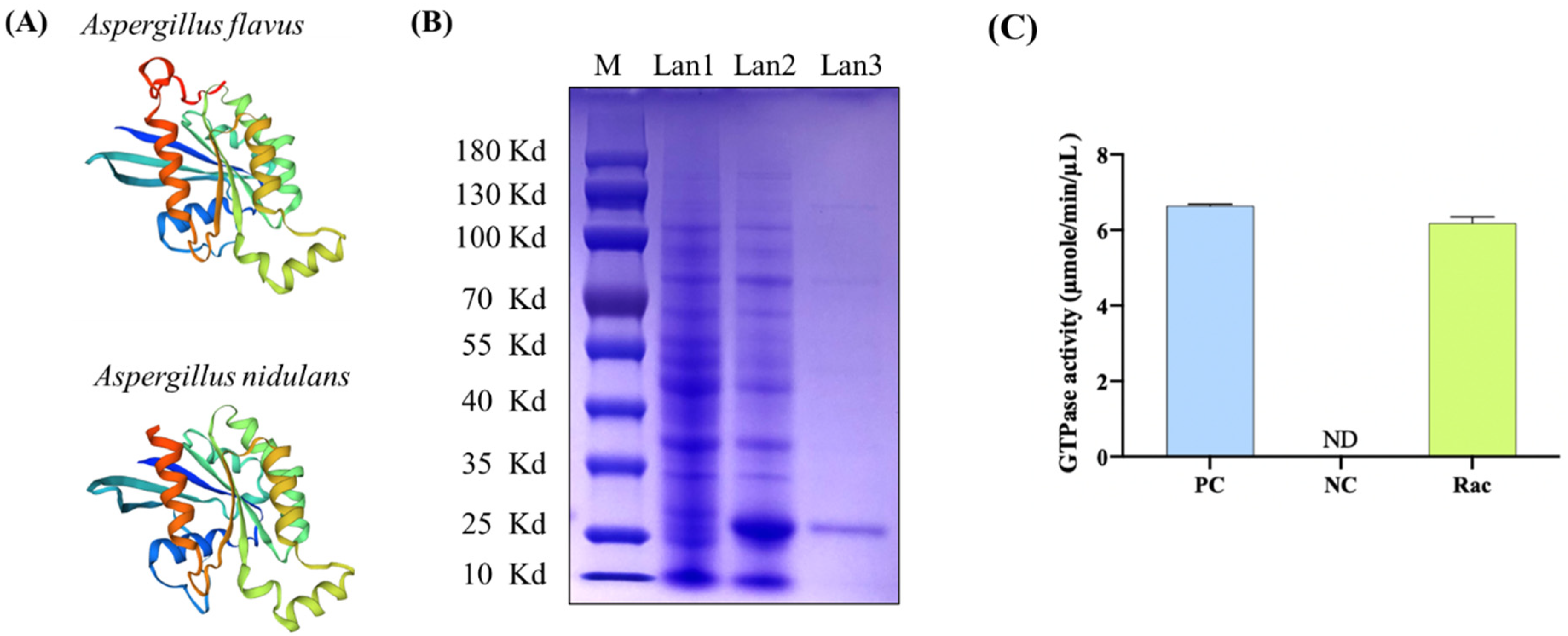
2.2. Rac from A. flavus Is a GTPase
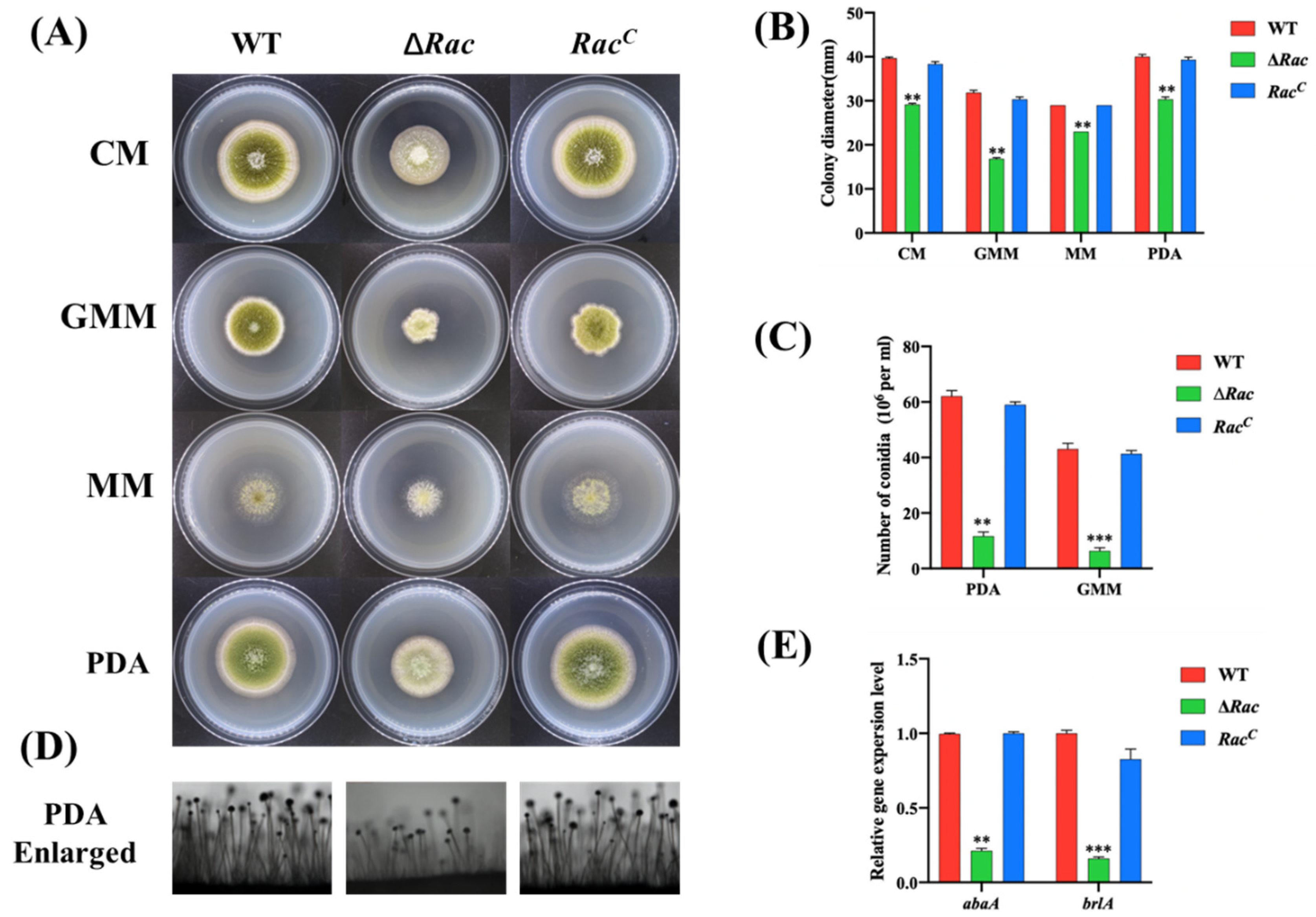
2.3. Rac Is Involved in Vegetative Growth and Conidiation
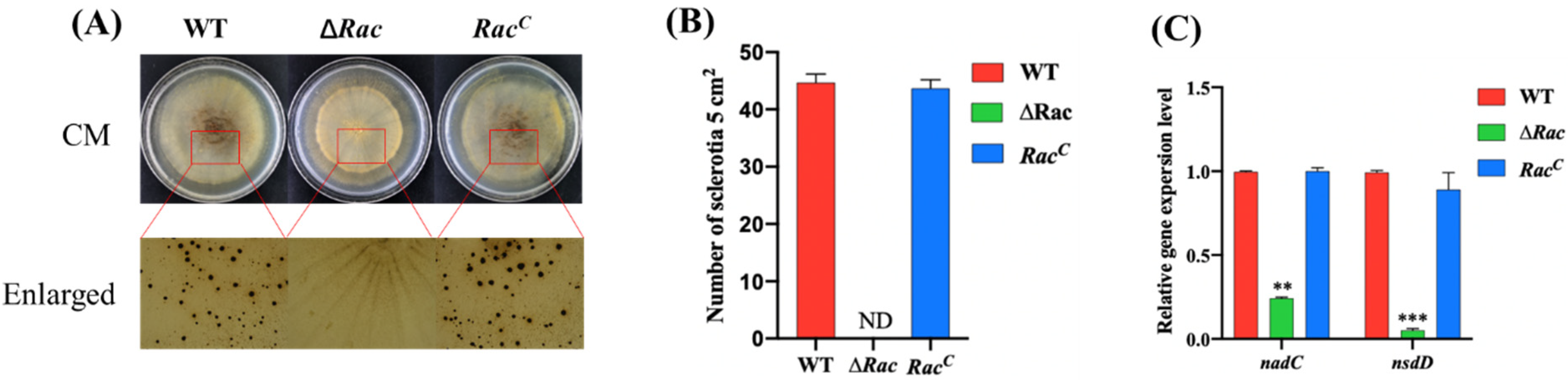
2.4. Rac Is Required for Sclerotial Formation in A. flavus
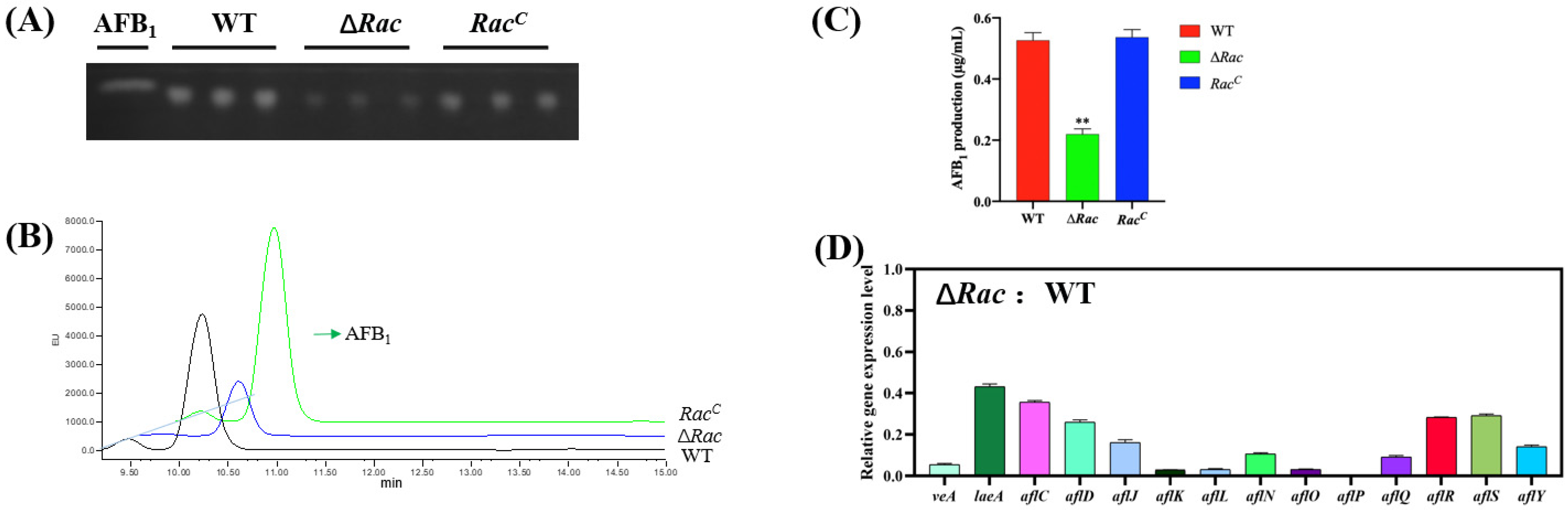
2.5. Effect of Rac on Aflatoxin Biosynthesis in A. flavus
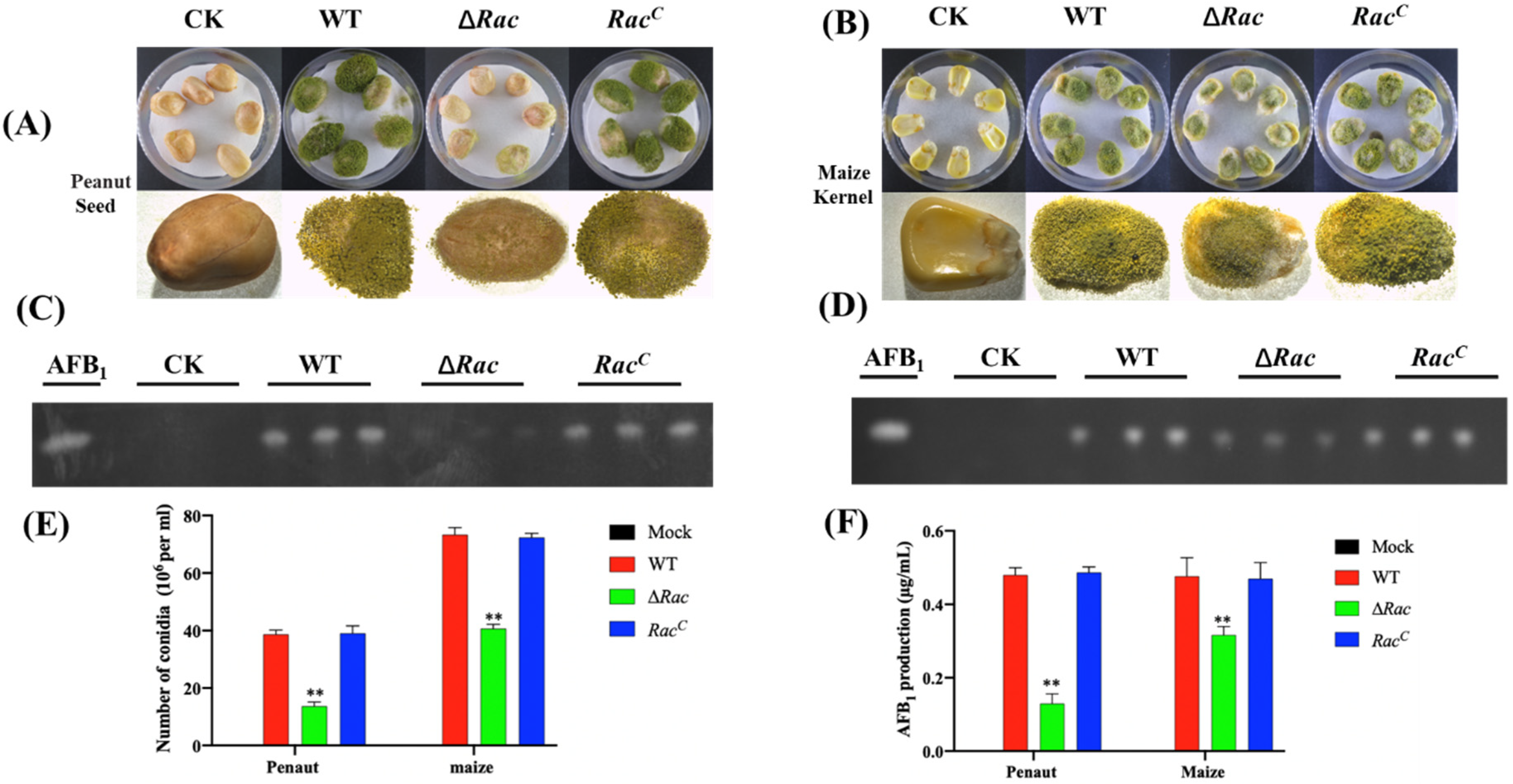
2.6. Role of Rac in Seed Infection
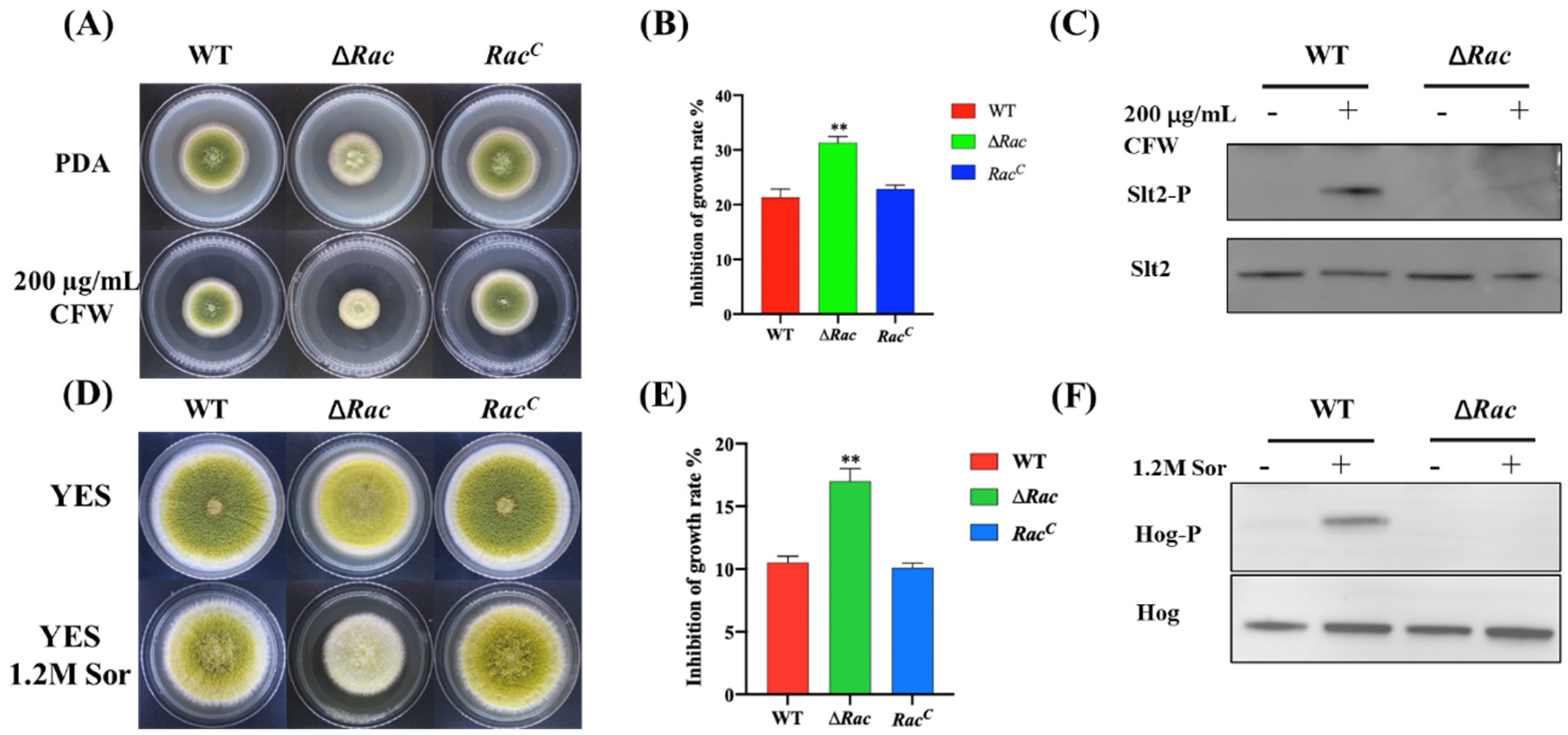
2.7. Rac Contributes to Cell Wall and Osmotic Stress Response
3. Discussion
4. Materials and Methods
4.1. Strains and Media
4.2. Domain and Phylogenetic Tree Analysis
4.3. Expression of Recombinant Protein and Detection of GTPase Activity
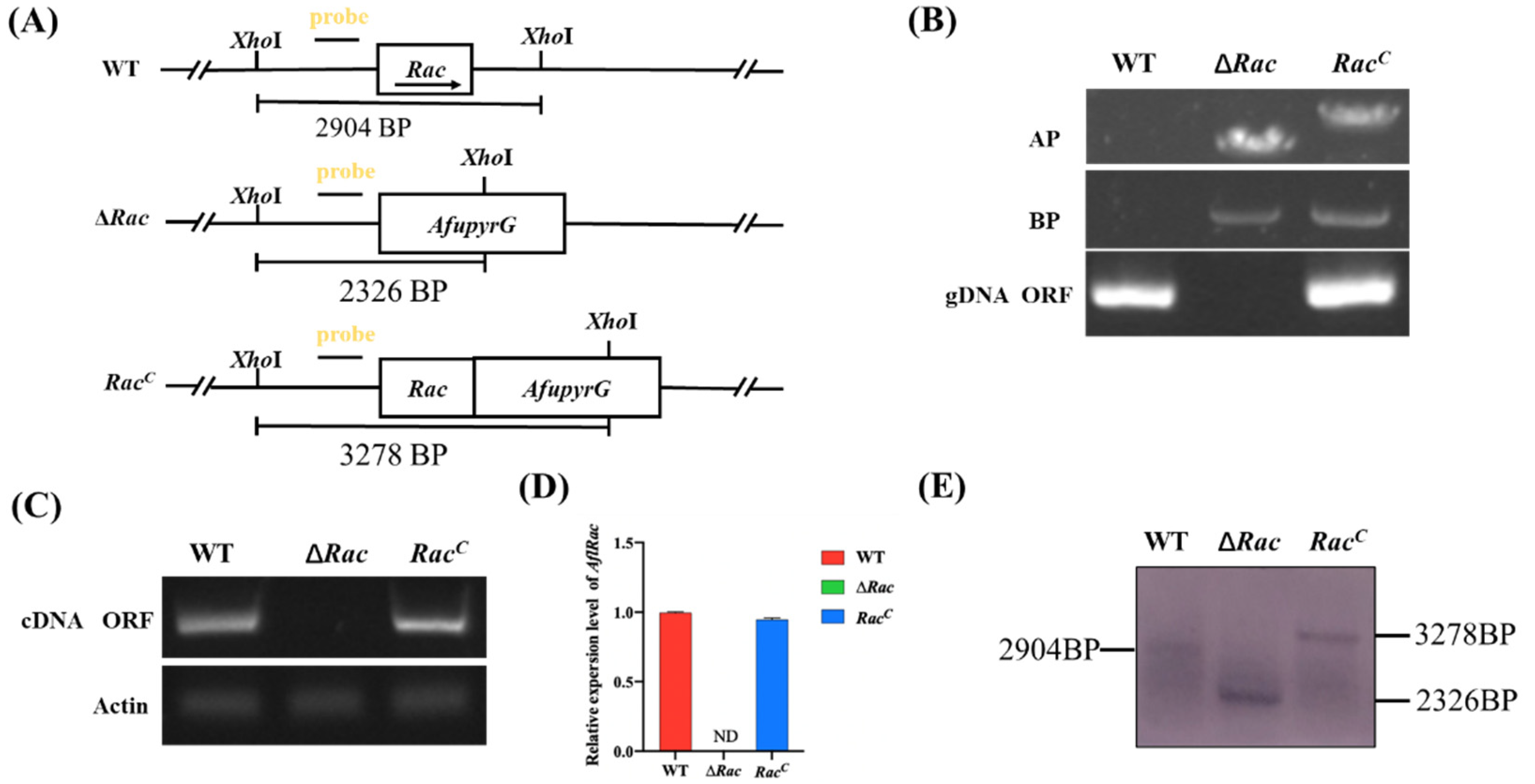
4.4. Construction of Mutant Strains
4.5. Mycelial Growth, Conidiation and Sclerotia Analysis
4.6. Aflatoxins Analysis
4.7. Seeds Infections
4.8. Cell Wall Stress and Osmotic Pressure Stress
4.9. Western Blot Analysis
4.10. Quantitative RT-PCR Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A Global Concern for Food Safety, Human Health and Their Management. Front. Microbiol. 2016, 7, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Roebuck, B.D.; Wogan, G.N.; Groopman, J.D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicol. Sci. 2011, 120 (Suppl. S1), S28–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roze, L.V.; Hong, S.Y.; Linz, J.E. Aflatoxin biosynthesis: Current frontiers. Annu. Rev. Food Sci. Technol. 2013, 4, 293–311. [Google Scholar] [CrossRef]
- Arenas-Huertero, F.; Zaragoza-Ojeda, M.; Sánchez-Alarcón, J.; Milić, M.; Šegvić Klarić, M.; Montiel-González, J.M.; Valencia-Quintana, R. Involvement of Ahr Pathway in Toxicity of Aflatoxins and Other Mycotoxins. Front. Microbiol. 2019, 10, 2347. [Google Scholar] [CrossRef] [Green Version]
- Strosnider, H.; Azziz-Baumgartner, E.; Banziger, M.; Bhat, R.V.; Breiman, R.; Brune, M.N.; DeCock, K.; Dilley, A.; Groopman, J.; Hell, K.; et al. Workgroup report: Public health strategies for reducing aflatoxin exposure in developing countries. Environ. Health Perspect. 2006, 114, 1898–1903. [Google Scholar] [CrossRef]
- Robens, J.; Cardwell, K. The Costs of Mycotoxin Management to the USA: Management of Aflatoxins in the United States. Toxin Rev. 2003, 22, 139–152. [Google Scholar] [CrossRef]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef] [Green Version]
- Heasman, S.J.; Ridley, A.J. Mammalian Rho GTPases: New insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef]
- Mittnacht, S.; Paterson, H.; Olson, M.F.; Marshall, C.J. Ras signalling is required for inactivation of the tumour suppressor pRb cell-cycle control protein. Curr. Biol. 1997, 7, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Bokoch, G.M. Regulation of the phagocyte respiratory burst by small GTP-binding proteins. Trends Cell Biol. 1995, 5, 109–113. [Google Scholar] [CrossRef]
- De Curtis, I. Functions of Rac GTPases during neuronal development. Dev. Neurosci. 2008, 30, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Liao, Y.J.; Jan, L.Y.; Jan, Y.N. Distinct morphogenetic functions of similar small GTPases: Drosophila Drac1 is involved in axonal outgrowth and myoblast fusion. Genes Dev. 1994, 8, 1787–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, J.; Luo, L. Rho GTPases regulate axon growth through convergent and divergent signaling pathways. Neuron 2004, 44, 779–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virag, A.; Lee, M.P.; Si, H.; Harris, S.D. Regulation of hyphal morphogenesis by cdc42 and rac1 homologues in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 1579–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Barker, B.M.; Grahl, N.; Puttikamonkul, S.; Bell, J.D.; Craven, K.D.; Cramer, R.A., Jr. The small GTPase RacA mediates intracellular reactive oxygen species production, polarized growth, and virulence in the human fungal pathogen Aspergillus fumigatus. Eukaryot. Cell 2011, 10, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Mahlert, M.; Leveleki, L.; Hlubek, A.; Sandrock, B.; Bölker, M. Rac1 and Cdc42 regulate hyphal growth and cytokinesis in the dimorphic fungus Ustilago maydis. Mol. Microbiol. 2006, 59, 567–578. [Google Scholar] [CrossRef]
- Boyce, K.J.; Hynes, M.J.; Andrianopoulos, A. Control of morphogenesis and actin localization by the Penicillium marneffei RAC homolog. J. Cell Sci. 2003, 116, 1249–1260. [Google Scholar] [CrossRef] [Green Version]
- Kost, B.; Lemichez, E.; Spielhofer, P.; Hong, Y.; Tolias, K.; Carpenter, C.; Chua, N.H. Rac homologues and compartmentalized phosphatidylinositol 4, 5-bisphosphate act in a common pathway to regulate polar pollen tube growth. J. Cell Biol. 1999, 145, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, C.A.; Beckerich, J.M.; Gaillardin, C.; Rachubinski, R.A. A rac homolog is required for induction of hyphal growth in the dimorphic yeast Yarrowia lipolytica. J. Bacteriol. 2000, 182, 2376–2386. [Google Scholar] [CrossRef] [Green Version]
- Kwon, M.J.; Nitsche, B.M.; Arentshorst, M.; Jørgensen, T.R.; Ram, A.F.; Meyer, V. The transcriptomic signature of RacA activation and inactivation provides new insights into the morphogenetic network of Aspergillus niger. PLoS ONE 2013, 8, e68946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, S.P.; Milde, L.; Trapp, M.K.; Frisvad, J.C.; Keller, N.P.; Bok, J.W. Requirement of LaeA for secondary metabolism and sclerotial production in Aspergillus flavus. Fungal Genet. Biol. 2008, 45, 1422–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef]
- Du, W.; Obrian, G.R.; Payne, G.A. Function and regulation of aflJ in the accumulation of aflatoxin early pathway intermediate in Aspergillus flavus. Food Addit. Contam. 2007, 24, 1043–1050. [Google Scholar] [CrossRef]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. Production of cyclopiazonic acid, aflatrem, and aflatoxin by Aspergillus flavus is regulated by veA, a gene necessary for sclerotial formation. Appl. Microbiol. Biotechnol. 2007, 73, 1158–1168. [Google Scholar] [CrossRef]
- Rolke, Y.; Tudzynski, P. The small GTPase Rac and the p21-activated kinase Cla4 in Claviceps purpurea: Interaction and impact on polarity, development and pathogenicity. Mol. Microbiol. 2008, 68, 405–423. [Google Scholar] [CrossRef]
- Di Ciano-Oliveira, C.; Thirone, A.C.; Szászi, K.; Kapus, A. Osmotic stress and the cytoskeleton: The R(h)ole of Rho GTPases. Acta Physiol. 2006, 187, 257–272. [Google Scholar] [CrossRef]
- Qin, L.; Li, D.; Zhao, J.; Yang, G.; Wang, Y.; Yang, K.; Tumukunde, E.; Wang, S.; Yuan, J. The membrane mucin Msb2 regulates aflatoxin biosynthesis and pathogenicity in fungus Aspergillus flavus. Microb. Biotechnol. 2021, 14, 628–642. [Google Scholar] [CrossRef]
- Lan, H.; Wu, L.; Fan, K.; Sun, R.; Yang, G.; Zhang, F.; Yang, K.; Lin, X.; Chen, Y.; Tian, J.; et al. Set3 Is Required for Asexual Development, Aflatoxin Biosynthesis, and Fungal Virulence in Aspergillus flavus. Front. Microbiol. 2019, 10, 530. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Nie, X.; Yang, K.; Xu, P.; Wang, X.; Liu, M.; Yang, Y.; Chen, Z.; Wang, S. Molecular and structural basis of nucleoside diphosphate kinase-mediated regulation of spore and sclerotia development in the fungus Aspergillus flavus. J. Biol. Chem. 2019, 294, 12415–12431. [Google Scholar] [CrossRef]
- Available online: http://smart.embl-heidelberg.de/ (accessed on 20 August 2022).
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1. 0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Lan, H.; Wu, L.; Sun, R.; Keller, N.P.; Yang, K.; Ye, L.; He, S.; Zhang, F.; Wang, S. The HosA Histone Deacetylase Regulates Aflatoxin Biosynthesis Through Direct Regulation of Aflatoxin Cluster Genes. Mol. Plant Microbe Interact. 2019, 32, 1210–1228. [Google Scholar] [CrossRef]
- Lan, H.; Sun, R.; Fan, K.; Yang, K.; Zhang, F.; Nie, X.Y.; Wang, X.; Zhuang, Z.; Wang, S. The Aspergillus flavus Histone Acetyltransferase AflGcnE Regulates Morphogenesis, Aflatoxin Biosynthesis, and Pathogenicity. Front. Microbiol. 2016, 7, 1324. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Liu, Y.; Wang, S.; Wu, L.; Xie, R.; Lan, H.; Fasoyin, O.E.; Wang, Y.; Wang, S. Cyclase-Associated Protein Cap with Multiple Domains Contributes to Mycotoxin Biosynthesis and Fungal Virulence in Aspergillus flavus. J. Agric. Food Chem. 2019, 67, 4200–4213. [Google Scholar] [CrossRef]
- Jia, K.; Yan, L.; Jia, Y.; Xu, S.; Yan, Z.; Wang, S. aflN Is Involved in the Biosynthesis of Aflatoxin and Conidiation in Aspergillus flavus. Toxins 2021, 13, 831. [Google Scholar] [CrossRef]
- Tan, C.; Deng, J.L.; Zhang, F.; Zhu, Z.; Yan, L.J.; Zhang, M.J.; Yuan, J.; Wang, S.H. CWI pathway participated in vegetative growth and pathogenicity through a downstream effector AflRlm1 in Aspergillus flavus. iScience 2021, 24, 103159. [Google Scholar] [CrossRef]
- Yang, M.; Zhu, Z.; Bai, Y.; Zhuang, Z.; Ge, F.; Li, M.; Wang, S. A novel phosphoinositide kinase Fab1 regulates biosynthesis of pathogenic aflatoxin in Aspergillus flavus. Virulence 2021, 12, 96–113. [Google Scholar] [CrossRef]
- Yang, K.; Liang, L.; Ran, F.; Liu, Y.; Li, Z.; Lan, H.; Gao, P.; Zhuang, Z.; Zhang, F.; Nie, X.; et al. The DmtA methyltransferase contributes to Aspergillus flavus conidiation, sclerotial production, aflatoxin biosynthesis and virulence. Sci. Rep. 2016, 6, 23259. [Google Scholar] [CrossRef] [Green Version]
- Mengjuan, Z.; Guanglan, L.; Xiaohua, P.; Weitao, S.; Can, T.; Xuan, C.; Yanling, Y.; Zhenhong, Z. The PHD transcription factor Cti6 is involved in the fungal colonization and aflatoxin B1 biological synthesis of Aspergillus flavus. IMA Fungus 2021, 12, 12. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, Z.; Guo, Z.; Li, D.; Zhang, F.; Shen, J.; Zhang, Y.; Wang, S.; Zhuang, Z. PbsB Regulates Morphogenesis, Aflatoxin B1 Biosynthesis, and Pathogenicity of Aspergillus flavus. Front. Cell Infect. Microbiol. 2018, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Yang, M.; Bai, Y.; Ge, F.; Wang, S. Antioxidant-related catalase CTA1 regulates development, aflatoxin biosynthesis, and virulence in pathogenic fungus Aspergillus flavus. Environ. Microbiol. 2020, 22, 2792–2810. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Available online: https://www.graphpad.com (accessed on 20 August 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Genotype Descripion | Source |
|---|---|---|
| A. flavus CA14 | Δku70, ΔpyrG | Purchased from FGSC (Manhattan, NY, USA) |
| Wild-type (WT) | Δku70, ΔpyrG::AfpyrG | This study |
| ΔAflRac | Δku70, ΔpyrG::AfpyrG, ΔAflRac | This study |
| ΔAflRacC | Δku70, ΔAflmsb2:: Aflmsb2, pyrG | This study |
| E. coil DH5α | Takara | |
| E. coil Rosetta | Takara |
| Primer | Sequence (5′-3′) | Characteristics |
|---|---|---|
| Rac-p1 | GGTTTCCTCAACGGTGTT | For amplifying |
| Rac-p3 | GGGTGAAGAGCATTGTTTGAGGC TCTTTCAGAATCTGCGATAT | 5′UTR of ΔRac |
| Rac-p6 | GCATCAGTGCCTCCTCTCAGA CAATTTTCTCCCGACTATAA | For amplifying 3′ UTR of ΔRac |
| Rac-p8 | CATCATTCCTAATGTGCTT | |
| Rac-p2 | GTTGGGAAAGAGGTGTCG | For fusion PCR |
| Rac-p7 | GTCTCAGTGCGTGTTGCT | of ΔRac |
| pyrG-F | GCCTCAAACAATGCTCTTCACCC | For amplifying |
| pyrG-R | GTCTGAGAGGAGGCACTGATGC | A. fumigatus pyrG |
| Rac-p9 | CGGCTAATAGACGACCAAT | For validating ORF |
| Rac-p10 | AGACGCTCTTCAGATTACG | |
| Rac-C-p1 | GATTGTTCCCTTATCATTG | For amplifying |
| Rac-C-p2 | CGAACAAGGTGTATAGTCT | ORF of RacC |
| Rac-C-p3 | GATTGTTCCCTTATCATTG | |
| Rac-C-p4 | GGGTGAAGAGCATTGTTTGAG GCCTACAGAATCAGACATTTGCTCTTC | For amplifying 5′UTR of RacC |
| Rac-C-p5 | GCATCAGTGCCTCCTCTCAGA CATCGATTCTTATAATTTTCTCCCGA | For amplifying 3′UTR of RacC |
| Rac-C-p6 | CGAACAAGGTGTATAGTCT | |
| Rac-C-O1 | CCTGCCTTGTGGTATTTC | For fusion PCR |
| Rac-C-O2 | ATGCTTTGCTGACGCTAT | of RacC |
| Rac-S-O1 | ACCAGCCATTCAGTGTTC | For Southern blot |
| Rac-S-O2 | AATTGCAGTGACAAGAGATG | |
| pyrG-907-F | ATGACGGCGATGTAGGGA | For screening ΔRac mutant |
| pyrG-919-R | CGACATCCTCACCGATTTCA | |
| Rac-G-O1 | AATGGGTCGCGGATCCCTGGAAGTTC TGTTCCAGGGGCCCATGGCGACCGGT | For amplifying |
| Rac-G-O2 | GGTGGTGGTGGTGGTGCTCGAGCTACAGA ATCAGACATTTGCTC | complete Rac gene |
| Primer | Sequence (5′-3′) | Characteristics |
|---|---|---|
| abaA-qRT-F | TCTTCGGTTGATGGATGATTTC | For amplifying abaA |
| abaA-qRT-R | CCGTTGGGAGGCTGGGT | |
| brlA-qRT-F | GCCTCCAGCGTCAACCTTC | For amplifying brlA |
| brlA-qRT-R | TCTCTTCAAATGCTCTTGCCTC | |
| nsdC-qRT-F | GCCAGACTTGCCAATCAC | For amplifying nsdC |
| nsdC-qRT-R | CATCCACCTTGCCCTTTA | |
| nsdD-qRT-F | GGACTTGCGGGTCGTGCTA | For amplifying nsdD |
| nsdD -qRT-R | AGAACGCTGGGTCTGGTGC | |
| veA -qRT-F | TATCATTCCGTGGCTCAAT | For amplifying veA |
| veA -qRT-R | GAGAGGTACTGCTGGATG | |
| laeA -qRT-F | TTGTTGGGGTTGACCTTGCT | For amplifying laeA |
| laeA -qRT-R | GCCATCCCATCACACTTCCA | |
| aflC-qRT-F | TTACGCTGCGATCAGTTCCTC | For amplifying aflC |
| aflC-qRT-R | CGACTCGCATTACAGCATCTAAC | |
| aflD-qRT-F | GTGGTGGTTGCCAATGCG | For amplifying aflD |
| aflD-qRT-R | CTGAAACAGTAGGACGGGAGC | |
| aflJ-qRT-F | CGGCGTATGAGGAGAATG | For amplifying aflJ |
| aflJ-qRT-R | CTTCATCAACCTGGCATCA | |
| aflK-qRT-F | GAGCGACAGGAGTAACCGTAAG | For amplifying aflK |
| aflK-qRT-R | CCGATTCCAGACACCATTAGCA | |
| aflL-qRT-F | GGCTGCGGAACTGTATTG | For amplifying aflL |
| aflL-qRT-R | TGTGGAGTGCTGGAAGAG | |
| aflN-qRT-F | TTCATTCCTGAGCGATGG | For amplifying aflN |
| aflN-qRT-R | CGTATGCTGGCGTAATATC | |
| aflO-qRT-F | GATTGGGATGTGGTCATGCGATT | For amplifying aflO |
| aflO-qRT-R | GCCTGGGTCCGAAGAATGC | |
| aflP-qRT-F | ACGAAGCCACTGGTAGAGGAGATG | For amplifying aflP |
| aflP-qRT-R | GTGAATGACGGCAGGCAGGT | |
| aflQ-qRT-F | GTCGCATATGCCCCGGTCGG | For amplifying aflQ |
| aflQ-qRT-R | GGCAACCAGTCGGGTTCCGG | |
| aflR-qRT-F | AAAGCACCCTGTCTTCCCTAAC | For amplifying aflR |
| aflR-qRT-R | GAAGAGGTGGGTCAGTGTTTGTAG | |
| aflS-qRT-F | AAGCTAAGGCCGAGTCTGG | For amplifying aflS |
| aflS-qRT-R | CAGGTTGTGTTGCTGTTGATAG | |
| aflY-qRT-F | AGGCAGACTTTCTAACACT | For amplifying aflY |
| aflY-qRT-R | CCTTCAGTTCCACACCAA | |
| Rac -qRT-p9 | GGTGACGGTGCTGTTGGA | |
| Rac -qRT-p10 | CGGGTCGTGGATTGAGAA | |
| actin-qRT-F | ACGGTGTCGTCACAAACTGG | For amplifying actin |
| actin -qRT-R | CGGTTGGACTTAGGGTTGATAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, L.; Yang, L.; Zhao, J.; Zeng, W.; Su, M.; Wang, S.; Yuan, J. GTPase Rac Regulates Conidiation, AFB1 Production and Stress Response in Pathogenic Fungus Aspergillus flavus. Toxins 2022, 14, 581. https://doi.org/10.3390/toxins14090581
Qin L, Yang L, Zhao J, Zeng W, Su M, Wang S, Yuan J. GTPase Rac Regulates Conidiation, AFB1 Production and Stress Response in Pathogenic Fungus Aspergillus flavus. Toxins. 2022; 14(9):581. https://doi.org/10.3390/toxins14090581
Chicago/Turabian StyleQin, Ling, Lan Yang, Jiaru Zhao, Wanlin Zeng, Minxuan Su, Shihua Wang, and Jun Yuan. 2022. "GTPase Rac Regulates Conidiation, AFB1 Production and Stress Response in Pathogenic Fungus Aspergillus flavus" Toxins 14, no. 9: 581. https://doi.org/10.3390/toxins14090581