First Report on Microcystin-LR Occurrence in Water Reservoirs of Eastern Cuba, and Environmental Trigger Factors

 ,
,
Abstract
:1. Introduction
2. Results
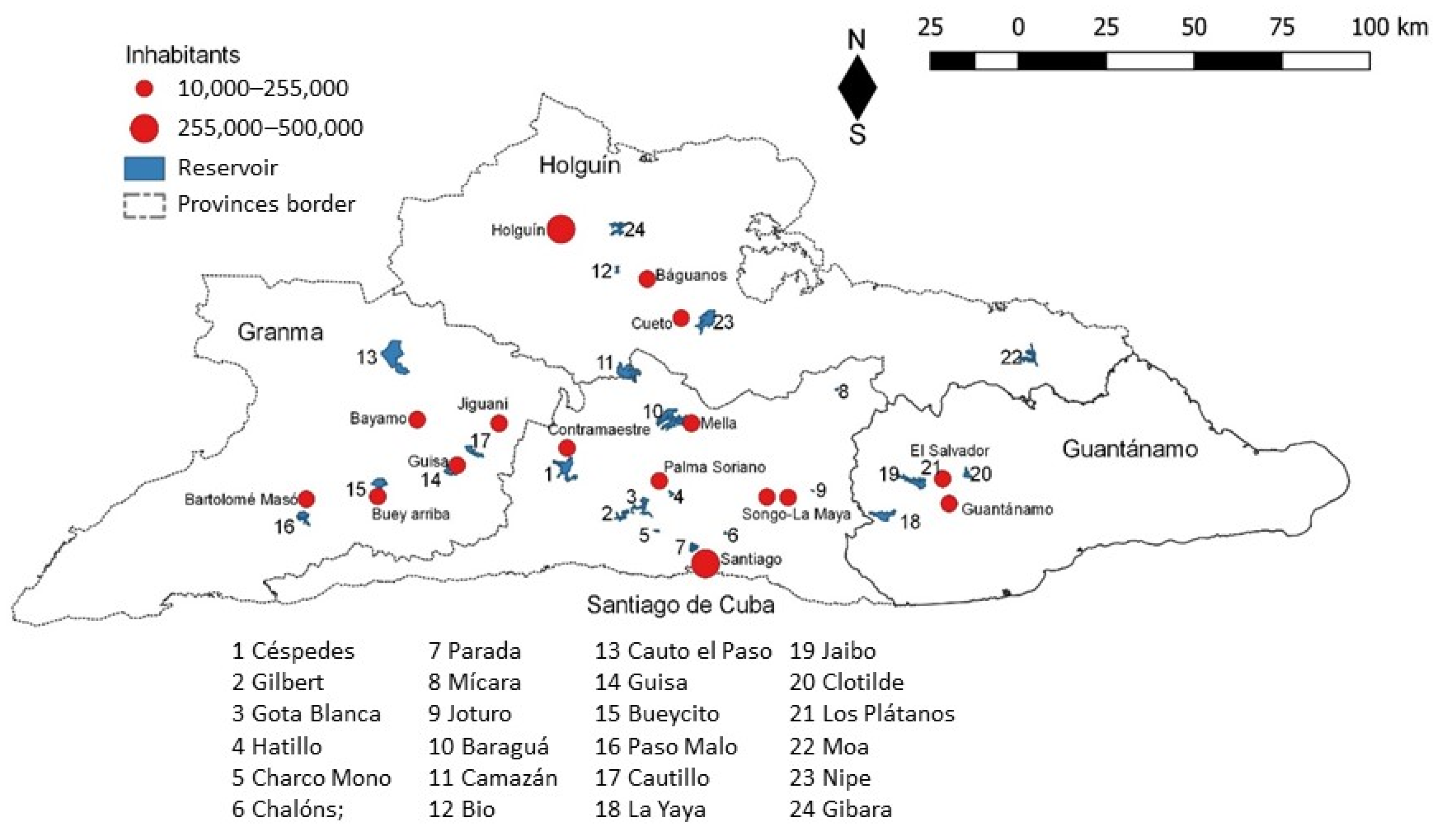
2.1. Catchment Description
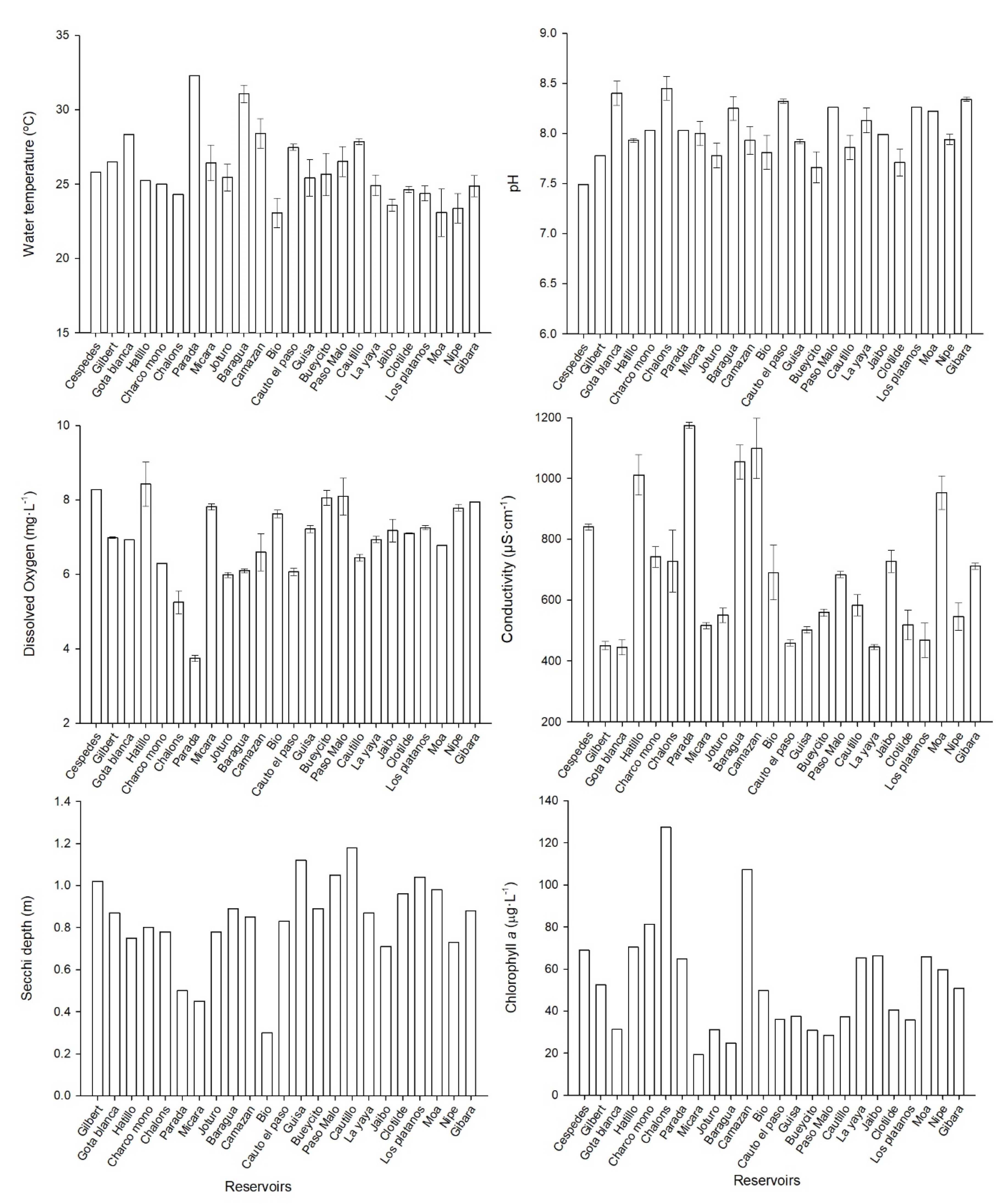
2.2. Physicochemical Parameters
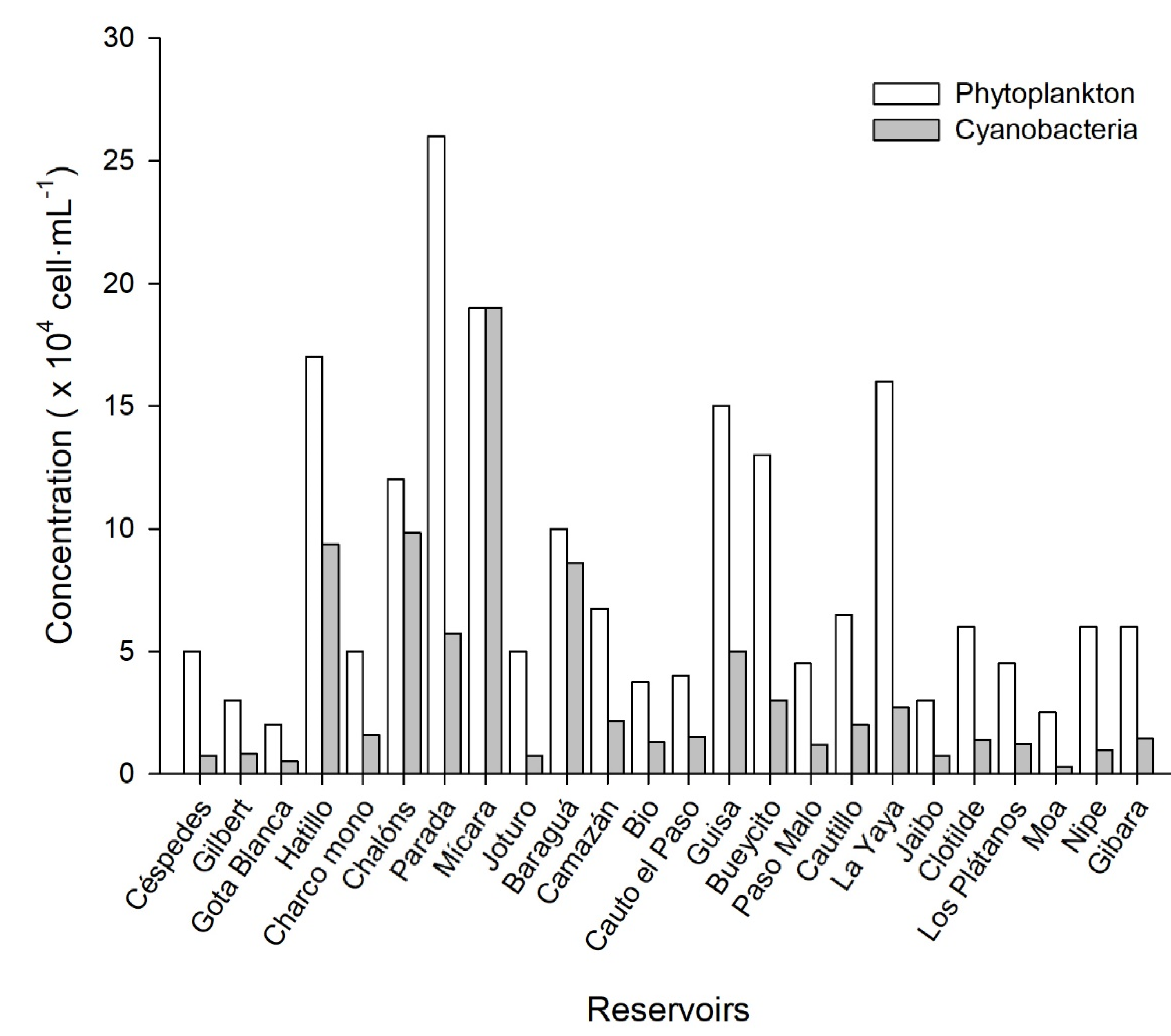
2.3. Phytoplankton/Cyanobacteria
2.4. Microcystin-LR Concentration
2.5. Correlation Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Area
5.2. Sampling
5.3. Catchment Analysis and Land Uses
5.4. Physico-Chemical Analysis
5.5. Phytoplankton Analysis and Taxonomic Identification
5.6. Microcystin-LR (MC-LR) Analysis
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Páll, E.; Niculae, M.; Kiss, T.; Şandru, C.D.; Spînu, M. Human impact on the microbiological water quality of the rivers. J. Med. Microbiol. 2013, 62, 1635–1640. [Google Scholar] [CrossRef]
- Teta, R.; Romano, V.; Della Sala, G.; Picchio, S.; De Sterlich, C.; Mangoni, A.; Di Tullio, G.; Costantino, V.; Lega, M. Cyanobacteria as indicators of water quality in Campania coasts, Italy: A monitoring strategy combining remote/proximal sensing and in situ data. Environ. Res. Lett. 2017, 12, 024001. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Raffoul, M.; Enanga, E.; Senar, O.; Creed, I.; Trick, C. Assessing the potential health risk of cyanobacteria and cyanotoxins in Lake Naivasha, Kenya. Hydrobiologia 2020, 847, 1041–1056. [Google Scholar] [CrossRef]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Dalu, T.; Wasserman, R.J. Cyanobacteria dynamics in a small tropical reservoir: Understanding spatio-temporal variability and influence of environmental variables. Sci. Total Environ. 2018, 643, 835–841. [Google Scholar] [CrossRef]
- Weirich, C.A.; Miller, T.R. Freshwater Harmful Algal Blooms: Toxins and Children’s Health. Curr. Probl. Pediatr. Adolesc. Health Care 2014, 44, 2–24. [Google Scholar] [CrossRef]
- Nazari-Sharabian, M.; Ahmad, S.; Karakouzian, M. Climate Change and Eutrophication: A Short Review. Eng. Technol. Appl. Sci. Res. 2018, 8, 3668–3672. [Google Scholar] [CrossRef]
- O’neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Niamien-Ebrottie, J.E.; Bhattacharyya, S.; Deep, P.R.; Nayak, B. Cyanobacteria and cyanotoxins in the World. Ijar 2015, 1, 563–569. [Google Scholar]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [Green Version]
- Arteaga, D.N. El Desarrollo Humano y la Generación de Capacidades. Diálogos 2010, 5, 47–58. [Google Scholar]
- Piedra, J.I.G. El Manejo de Cuencas en Cuba. H. Cotler (Comp.), El Manejo Integral de Cuencas en México: Estudios y Reflexiones para Orientar la Política Ambiental; Instituto Nacional de Ecología: Tlalpan, Mexico, 2004; pp. 21–40. [Google Scholar]
- CITMA. Enfrentamiento al Cambio Climático en la República de Cuba Tarea Vida; CITMATEL: La Habana, Cuba, 2017. [Google Scholar]
- Rodríguez Tito, J.C.; Gómez, L.L.M. Estado trófico de 24 embalses de agua en el oriente de Cuba. Rev. Cuba. Quím. 2020, 32, 136–153. [Google Scholar]
- Gomez Luna, L.; Álvarez Hubert, I.; Rodríguez, T.J.C.; Álamo Diaz, B. Cianobacterias: Un Riesgo Emergente en Embalses de Agua de Santiago de Cuba; Universidad de Oriente: Santiago de Cuba, Cuba, 2011; p. 76. [Google Scholar]
- Gomez, L.; Álvarez, I.; Rodríguez, J.C. CUBA: Toxic cyanobacteria risk assessment, research and management. In Current Approaches to Cyanotoxin Risk Assessment, Risk Management and Regulations in Different Countries; Chorus, I., Ed.; Federal Environment Agency (Umweltbundesamt): Dessau-Roßlau, Germany, 2012; pp. 40–48. [Google Scholar]
- Rodríguez Tito, J.C.; Pérez Silva, R.M.; Gomez Luna, L.; Álvarez Hubert, I. Evaluación químico analítica y microbiológica de los embalses Chalons y Parada de Santiago de Cuba. Rev. Cuba. Quím. 2017, 29, 302–307. [Google Scholar]
- Jiang, Y.; Ji, B.; Wong, R.N.S.; Wong, M.H. Statistical study on the effects of environmental factors on the growth and microcystins production of bloom-forming cyanobacterium—Microcystis aeruginosa. Harmful Algae 2008, 7, 127–136. [Google Scholar] [CrossRef]
- Gerard, C.; Poullain, V.; Lance, E.; Acou, A.; Brient, L.; Carpentier, A. Influence of toxic cyanobacteria on community structure and microcystin accumulation of freshwater molluscs. Environ. Pollut. 2009, 157, 609–617. [Google Scholar] [CrossRef]
- Munawar, S.; Aisha, K. Occurrence of bloom of blue green algae from the coast of Buleji, Karachi, Pakistan. Int. J. Biol. Res. 2017, 5, 31–34. [Google Scholar]
- Bouma-Gregson, K.; Power, M.E.; Bormans, M. Rise and fall of toxic benthic freshwater cyanobacteria (Anabaena spp.) in the Eel river: Buoyancy and dispersal. Harmful Algae 2017, 66, 79–87. [Google Scholar] [CrossRef]
- Gomez, L.M.; Álamo, B.; Rodríguez Tito, J.C. Riesgo de contaminación con cianobacterias en tres embalses de agua de Santiago de Cuba. Medisan 2010, 14, 175–183. [Google Scholar]
- Salmaso, N.; Bernard, C.; Humbert, J.F.; Akçaalan, R.; Albay, M.; Ballot, A.; Catherine, A.; Fastner, J.; Häggqvist, K.; Horecká, M.; et al. Basic Guide to Detection and Monitoring of Potentially Toxic Cyanobacteria. In Handbook of Cyanobacterial Monitoring Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 48–69. [Google Scholar]
- Bittencourt-Oliveira, M.; Piccin Santos, V.; Moura, A.N.; Aragao Tavares, N.K.; Cordeiro Araujo, M.K. Cyanobacteria, microcystins and cylindrospermopsin in public drinking supply reservoirs of Brazil. An. Acad. Bras. Ciênc. 2014, 86, 297–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccin-Santos, V.; Bittencourt-Oliveira, M.C. Toxic cyanobacteria in four Brazilian water supply reservoirs. J. Environ. Prot. 2012, 3, 16872. [Google Scholar] [CrossRef] [Green Version]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.F. A review of current knowledge on toxic benthic freshwater cyanobacteria-Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Sant’Anna, C.L.; Azevedo, M.D.P.; Werner, V.R.; Dogo, C.R.; Rios, F.R.; de Carvalho, L.R. Review of toxic species of Cyanobacteria in Brazil. Algol. Stud. 2008, 126, 251–265. [Google Scholar] [CrossRef]
- Lins, R.P.M.; Barbosa, L.G.; Minillo, A.; De Ceballos, B.S.O. Cyanobacteria in a eutrophicated reservoir in a semi-arid region in Brazil: Dominance and microcystin events of blooms. Braz. J. Bot. 2016, 39, 583–591. [Google Scholar] [CrossRef]
- Lyon-Colbert, A.; Su, S.; Cude, C. A systematic literature review for evidence of Aphanizomenon flos-aquae toxigenicity in recreational waters and toxicity of dietary supplements: 2000–2017. Toxins 2018, 10, 254. [Google Scholar] [CrossRef] [Green Version]
- Manning, S.R.; Nobles, D.R. Impact of global warming on water toxicity: Cyanotoxins. Curr. Opin. Food Sci. 2017, 18, 14–20. [Google Scholar] [CrossRef]
- Svirčev, Z.; Obradović, V.; Codd, G.A.; Marjanović, P.; Spoof, L.; Drobac, D.; Tokodi, N.; Petković, A.; Nenin, T.; Simeunović, J. Massive fish mortality and Cylindrospermopsis raciborskii bloom in Aleksandrovac Lake. Ecotoxicology 2016, 25, 1353–1363. [Google Scholar] [CrossRef]
- Heddam, S.; Kisi, O. Extreme learning machines: A new approach for modeling dissolved oxygen (DO) concentration with and without water quality variables as predictors. Environ. Sci. Pollut. Res. 2017, 24, 16702–16724. [Google Scholar] [CrossRef]
- Carlson, R.E.; Simpson, J. Trophic State Equations. In A Coordinator’s Guide to Volunteer Lake Monitoring Methods; North American Lake Management Society: Madison, WI, USA, 1996; p. 96. [Google Scholar]
- Mazumder, A.; Havens, K.E. Nutrient-chlorophyll-Secchi relationships under contrasting grazer communities of temperate versus subtropical lakes. Can. J. Fish. Aquat. Sci. 1998, 55, 1652–1662. [Google Scholar] [CrossRef]
- Pham, T.L.; Shimizu, K.; Dao, T.S.; Motoo, U. First report on free and covalently bound microcystins in fish and bivalves from Vietnam: Assessment of risks to humans. Environ. Toxicol. Chem. 2017, 36, 2953–2957. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, X.Q.; Liu, L.M.; Zhang, W.J.; Guo, P.Y. Algae community and trophic state of subtropical reservoirs in southeast Fujian, China. Environ. Sci. Pollut. Res. 2012, 19, 1432–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, W.; Zhang, J.; Ding, T.; Stevenson, R.J.; Zhu, Y. Environmental factors regulating cyanobacteria dominance and microcystin production in a subtropical lake within the Taihu watershed, China. J. Zhejiang Univ. Sci. A 2012, 13, 311–322. [Google Scholar] [CrossRef]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N and P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [CrossRef]
- Umaña, V. One year weekly dynamics of limnological conditions and phytoplankton in Lake Bonilla, Costa Rica. Rev. Biol. Trop. 2016, 64, 1771–1781. [Google Scholar]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [Green Version]
- Barros, M.U.G.; Wilson, A.E.; Leitão, J.I.R.; Pereira, S.P.; Buley, R.P.; Fernandez-Figueroa, E.G.; Capelo-Neto, J. Environmental factors associated with toxic cyanobacterial blooms across 20 drinking water reservoirs in a semi-arid region of Brazil. Harmful Algae 2019, 86, 128–137. [Google Scholar] [CrossRef]
- Beaver, J.R.; Tausz, C.E.; Scotese, K.C.; Pollard, A.I.; Mitchell, R.M. Environmental factors influencing the quantitative distribution of microcystin and common potentially toxigenic cyanobacteria in U.S. lakes and reservoirs. Harmful Algae 2018, 78, 118–128. [Google Scholar] [CrossRef]
- Geada, P.; Pereira, R.N.; Vasconcelos, V.; Vicente, A.A.; Fernandes, B.D. Assessment of synergistic interactions between environmental factors on Microcystis aeruginosa growth and microcystin production. Algal Res. 2017, 27, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Mowe, M.A.D.; Mitrovic, S.M.; Lim, R.P.; Furey, A.; Yeo, D.C.J. Tropical cyanobacterial blooms: A review of prevalence, problem taxa, toxins and influencing environmental factors. J. Limnol. 2015, 74, 205–224. [Google Scholar] [CrossRef] [Green Version]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Beklioğlu, M.; Bucak, T.; Levi, E.E.; Erdoğan, Ş.; Özen, A.; Filiz, N.; Bezirci, G.; Çakıroğlu, A.İ.; Tavşanoğlu, Ü.N.; Gökçe, D.; et al. Influences of climate and nutrient enrichment on the multiple trophic levels of Turkish shallow lakes. Inland Waters 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Paerl, H.W. Impacts of Climate Change on Cyanobacteria in Aquatic Environments. In Climate Change and Microbial Ecology: Current Research and Future Trends; Caister Academic Press: Poole, UK, 2016; pp. 5–22. [Google Scholar]
- García Miranda, F.G.; Miranda Rosales, V. Eutrofización, una Amenaza para el Recurso Hídrico. In Impacto Socio-Ambiental, Territorios Sostenibles y Desarrollo Regional Desde el Turismo; Asociación Mexicana de Ciencias para el Desarrollo Regional A.C, Ed.; Universidad Nacional Autónoma de México: Mexico City, México, 2018; Volume 2. [Google Scholar]
- Haakonsson, S.; Rodríguez Gallego, L.; Somma, A.; Bonilla, S. Temperature and precipitation shape the distribution of harmful cyanobacteria in subtropical lotic and lentic ecosystems. Sci. Total Environ. 2017, 609, 1132–1139. [Google Scholar] [CrossRef]
- Thomas, M.K.; Litchman, E. Effects of temperature and nitrogen availability on the growth of invasive and native cyanobacteria. Hydrobiologia 2016, 763, 357–369. [Google Scholar] [CrossRef]
- Walls, J.T.; Wyatt, K.H.; Doll, J.C.; Rubenstein, E.M.; Rober, A.R. Hot and toxic: Temperature regulates microcystin release from cyanobacteria. Sci. Total Environ. 2018, 610, 786–795. [Google Scholar] [CrossRef]
- Havens, K.E. Cyanobacteria blooms: Effects on aquatic ecosystems. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: Berlin/Heidelberg, Germany, 2008; pp. 733–747. [Google Scholar]
- WHO. Guidelines for Drinking-Water Quality, 4th ed.; WHO: Geneva, Switzerland, 2017; 631p. [Google Scholar]
- Oficina Nacional de Normalización. Norma Cubana Higiene comunal—Fuentes de abastecimiento de agua—Calidad y protección sanitaria. In NC:1021; Oficina Nacional de Normalización: La Habana, Cuba, 2014. [Google Scholar]
- Amorim, C.A.; Dantas, Ê.W.; do Nascimento Moura, A. Modeling cyanobacterial blooms in tropical reservoirs: The role of physicochemical variables and trophic interactions. Sci. Total Environ. 2020, 744, 140659. [Google Scholar] [CrossRef]
- Brito, M.T.S.; Duarte-Neto, P.J.; Molica, R.J.R. Cylindrospermopsis raciborskii and Microcystis aeruginosa competing under different conditions of pH and inorganic carbon. Hydrobiologia 2018, 815, 253–266. [Google Scholar] [CrossRef]
- Jorgensen, S.E.; Fath, B.D. Encyclopedia of Ecology; Newnes: Oxford, UK, 2014. [Google Scholar]
- Chi-Yong, A.; Seung-Hyun, J.; Sook-Kyoung, Y.; Hee-Mock, O. Alternative Alert System for Cyanobacterial Bloom, Using Phycocyanin as a Level Determinant. J. Microbiol. 2007, 45, 98–104. [Google Scholar]
- Biagini, M. Valutazioni dellinquinamento delle acque correnti per mezzo del fitoplancton. Biol. Ital. 1980, 6, 29–31. [Google Scholar]
- WHO. Guidelines for Drinking-Water Quality, 4th ed.; WHO: Geneva, Switzerland, 2011; 541p. [Google Scholar]
- Scott, J.T.; McCarthy, M.J.; Otten, T.G.; Steffen, M.M.; Baker, B.C.; Grantz, E.M.; Wilhelm, S.W.; Paerl, H.W. Comment: An alternative interpretation of the relationship between TN: TP and microcystins in Canadian lakes. Can. J. Fish. Aquat. Sci. 2013, 70, 1265–1268. [Google Scholar] [CrossRef]
- Ahn, C.-Y.; Chung, A.-S.; Oh, H.M. Rainfall, phycocyanin, and N:P ratios related to cyanobacterial blooms in a Korean large reservoir. Hydrobiologia 2002, 474, 117–124. [Google Scholar] [CrossRef]
- Ouahid, Y.; del Campo, F.F. Typing of toxinogenic Microcystis from environmental samples by multiplex PCR. Appl. Microbiol. Biotechnol. 2009, 85, 405–412. [Google Scholar] [CrossRef]
- Ouahid, Y.; Pérez-Silva, G.; Campo, F.F.D. Identification of potentially toxic environmental microcystis by individual and multiple PCR amplification of specific microcystin synthetase gene regions. Environ. Toxicol. Int. J. 2005, 20, 235–242. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, R.; Hu, M.; Luo, J.; Li, J.; Liang, Q. Combining citizen science and land use data to identify drivers of eutrophication in the Huangpu River system. Sci. Total Environ. 2017, 584–585, 651–664. [Google Scholar] [CrossRef]
- Álvarez, X.; Valero, E.; Santos, R.M.; Varandas, S.; Fernandes, L.S.; Pacheco, F.A.L. Anthropogenic nutrients and eutrophication in multiple land use watersheds: Best management practices and policies for the protection of water resources. Land Use Policy 2017, 69, 1–11. [Google Scholar] [CrossRef]
- Beaver, J.R.; Manis, E.E.; Loftin, K.A.; Graham, J.L.; Pollard, A.I.; Mitchell, R.M. Land use patterns, ecoregion, and microcystin relationships in U.S. lakes and reservoirs: A preliminary evaluation. Harmful Algae 2014, 36, 57–62. [Google Scholar] [CrossRef]
- Jarvis, A.; Reuter, H.I.; Nelson, A.; Guevara, E. Hole-Filled SRTM for the Globe Version 4. 2008. CGIAR-CSI SRTM 90 m Database. Available online: https://srtm.csi.cgiar.org (accessed on 10 January 2021).
- Parmar, S. Sediment Yield Estimation Using SAGA GIS: A Case Study of Watershed–63 of Narmada River. J. Appl. Sci. Comput. 2018, 5, 888–895. [Google Scholar]
- García Fernández, J.M.; Gutiérrez Díaz, J.B. La Gestión de Cuencas Hidrográficas en Cuba; INRH; Consejo Nacional de Cuencas Hidrográficas: La Habana, Cuba, 2015. [Google Scholar]
- Oficina Nacional de Normalización. Norma Cubana. Evaluación de los objetos hídricos de uso pesquero. In NC:25; Oficina Nacional de Normalización: La Habana, Cuba, 1999. [Google Scholar]
- D’Elia, C.F.; Steudler, P.A.; Corwin, N. Determination of total nitrogen in aqueous samples using persulfate digestion. Limnol. Oceanogr. 1977, 22, 760–764. [Google Scholar] [CrossRef] [Green Version]
- Menzel, D.W.; Corwin, N. The measurement of total phosphorus in seawater based on the liberation of organically bound fractions by persulfate oxidation. Limnol. Oceanogr. 1965, 10, 280–282. [Google Scholar] [CrossRef]
- Ovsyanyi, E.; Zaburdaev, V.; Romanov, A.J.P.O. On the application of the units of measurement of the amount and composition of matter in oceanology. Phys. Oceanogr. 2004, 14, 243–254. [Google Scholar] [CrossRef]
- Catherine, A.; Maloufi, S.; Congestri, R.; Viaggiu, E.; Pilkaityte, R. Cyanobacterial samples: Preservation, enumeration, and biovolume measurements. In Handbook of Cyanobacterial Monitoring Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 313–330. [Google Scholar]
- Wher, J.; Sheath, R. (Eds.) Freshwater Algae of North America; Academic Press: San Diego, CA, USA, 2003; p. 916. [Google Scholar]
- Annadotter, H. Mass Developing Cyanobacteria in Temperate and Tropical Freshwater Bodies. Ecology, Toxins and Effects on Human Health. Ph.D. Thesis, University of Copenhagen, Copenhagen, Denmark, 2006. [Google Scholar]
- Cronberg, G.; Annadotter, H. (Eds.) Manual on Aquatic Cyanobacteria. A Photo Guide and a Synopsis of Their Toxicology; IOC-UNESCO, ISSHA: Copenhagen, Denmark, 2006; p. 106. [Google Scholar]
- Comas, A. Cianoprocariotas (Cianobacterias, Cianofíceas) y Microalgas Como Indicadores de la Calidad del Agua en el Embalse, Paso Bonito, Cienfuegos; Instituto de Información Científica y Tecnológica: Cienfuegos, Cuba, 2007. [Google Scholar]
- Comas, A. Catálogo de las Algas y Cianoprocariotas Dulciacuícolas de Cuba; Universidad “Carlos R. Rodríguez”: Cienfuegos, Cuba, 2009; p. 147. [Google Scholar]
- Guiry, M.; Guiry, G. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2014. [Google Scholar]
- Cerasino, L.; Meriluoto, J.; Bláha, L.; Carmeli, S.; Kaloudis, T.; Mazur-Marzec, H. Extraction of cyanotoxins from cyanobacterial biomass. In Handbook of Cyanobacterial Monitoring Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 350–353. [Google Scholar]
- Triantis, T.M.; Kaloudis, T.; Zervou, S.K.; Hiskia, A. Solid-Phase Extraction of Microcystins and Nodularin from Drinking Water. In Handbook of Cyanobacterial Monitoring Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 354–357. [Google Scholar]
- Spoof, L.; Vesterkvist, P.; Lindholm, T.; Meriluoto, J. Screening for cyanobacterial hepatotoxins, microcystins and nodularin in environmental water samples by reversed-phase liquid chromatography–electrospray ionisation mass spectrometry. J. Chromatogr. A 2003, 1020, 105–119. [Google Scholar] [CrossRef]
- Chen, Y.M.; Lee, T.H.; Lee, S.J.; Lin, J.Z.; Huang, R.; Chou, H.N. Potential of a simple solid-phase extraction method coupled to analytical and bioanalytical methods for an improved determination of microcystins in algal samples. J. Chromatogr. B 2006, 844, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Draper, W.M.; Xu, D.; Behniwal, P.; McKinney, M.J.; Jayalath, P.; Dhoot, J.S.; Wijekoon, D. Optimizing LC-MS-MS determination of microcystin toxins in natural water and drinking water supplies. Anal. Methods 2013, 5, 6796–6806. [Google Scholar] [CrossRef]
- Triantis, T.M.; Kaloudis, T.; Zervou, S.K.; Hiskia, A. Determination of Microcystins and Nodularin in Filtered and Drinking Water by LC-MS/MS. In Handbook of Cyanobacterial Monitoring Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 372–378. [Google Scholar]
- Kruve, A.; Rebane, R.; Kipper, K.; Oldekop, M.-L.; Evard, H.; Herodes, K.; Ravio, P.; Leito, I. Tutorial review on validation of liquid chromatography-mass spectrometry methods: Part I. Anal. Chim. Acta 2015, 870, 29–44. [Google Scholar] [CrossRef]
- Kruve, A.; Rebane, R.; Kipper, K.; Oldekop, M.-L.; Evard, H.; Herodes, K.; Ravio, P.; Leito, I. Tutorial review on validation of liquid chromatography–mass spectrometry methods: Part II. Anal. Chim. Acta 2015, 870, 8–28. [Google Scholar] [CrossRef]
- Evard, H.; Kruve, A.; Leito, I. Tutorial on estimating the limit of detection using LC–MS analysis, part I: Theoretical review. Anal. Chim. Acta. 2016, 942, 23–39. [Google Scholar] [CrossRef]
- Vuković, G.; Ðukić, M.; Bursić, V.; Popović, A.; Petrović, A.; Gvozdenac, S.; Grabić, J.; Zemunac, R. Method validation of microcystin-LR in water. In Proceedings of the 7th International Scientific and Professional Conference, “Water For All”, Osijek, Croatia, 9–10 March 2017; pp. 349–355. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reservoirs | MC-LR (Extracellular) | MC-LR (Intracellular) |
|---|---|---|
| Céspedes | 0.04 | ND |
| Gilbert | 0.10 | ND |
| Gota Blanca | 0.19 | ND |
| Hatillo | 2.38 | 36.50 |
| Charco Mono | ND | 0.015 |
| Chalóns | 2.56 | 10.69 |
| Parada | 2.21 | 1.93 |
| Mícara | 5.05 | 55.78 |
| Joturo | 0.35 | ND |
| Baraguá | 1.63 | 17.09 |
| Camazán | 0.14 | ND |
| Bio | 0.20 | 0.06 |
| Cauto el Paso | ND | ND |
| Guisa | 0.95 | 1.35 |
| Bueycito | 0.53 | 0.21 |
| Paso Malo | ND | ND |
| Cautillo | 14.49 | 65.76 |
| La Yaya | 14.58 | 60.52 |
| Jaibo | 0.83 | 1.41 |
| Clotilde | ND | ND |
| Los Plátanos | ND | ND |
| Moa | ND | ND |
| Nipe | ND | ND |
| Gibara | ND | ND |
| Air Temp | Cond | DO | pH | Chl a | Secchi | Phyto | Cyano | Phyco | TN | TP | MC-LR Water | MC-LR Intrac | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Water temp | 0.77 *** | −0.21 | −0.51 * | 0.12 | −0.26 | −0.41 * | 0.37 * | 0.09 | 0.24 | 0.11 | 0.38 * | 0.08 | 0.30 |
| Air temp | - | 0.08 | −0.37 * | 0.01 | −0.20 | −0.25 | 0.47 * | 0.06 | 0.22 | 0.26 | 0.50 * | −0.11 | −0.28 |
| Cond | - | 0.19 | 0.08 | −0.45 * | 0.36 * | −0.29 | −0.46 * | −0.31 | −0.10 | −0.31 | 0.14 | −0.03 | |
| DO | - | −0.29 | −0.23 | 0.41 * | −0.26 | 0.09 | −0.36 * | 0.03 | −0.12 | 0.01 | −0.26 | ||
| pH | - | 0.04 | 0.01 | 0.11 | 0.26 | −0.06 | 0.06 | 0.09 | −0.60 * | −0.38 | |||
| Chl a | - | 0.41 * | 0.12 | 0.01 | 0.36 * | −0.17 | 0.06 | 0.26 | 0.38 | ||||
| Secchi | - | −0.27 | −0.15 | −0.12 | 0.01 | −0.39 | −0.13 | 0.01 | |||||
| Phyto | - | 0.53 ** | 0.39 * | 0.16 | 0.42 * | −0.43 * | −0.40 | ||||||
| Cyano | - | 0.16 | 0.39 * | 0.28 | −0.62 ** | −0.55 * | |||||||
| Phyto | - | −0.19 | 0.37 * | 0.25 | 0.14 | ||||||||
| TN | - | 0.39 * | −0.67 ** | −0.37 | |||||||||
| TP | - | −0.18 | −0.26 | ||||||||||
| MC-LR water | - | 0.91 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tito, J.C.R.; Luna, L.M.G.; Noppe, W.N.; Hubert, I.A. First Report on Microcystin-LR Occurrence in Water Reservoirs of Eastern Cuba, and Environmental Trigger Factors. Toxins 2022, 14, 209. https://doi.org/10.3390/toxins14030209
Tito JCR, Luna LMG, Noppe WN, Hubert IA. First Report on Microcystin-LR Occurrence in Water Reservoirs of Eastern Cuba, and Environmental Trigger Factors. Toxins. 2022; 14(3):209. https://doi.org/10.3390/toxins14030209
Chicago/Turabian StyleTito, José Carlos Rodríguez, Liliana Maria Gomez Luna, Wim Noppe Noppe, and Inaudis Alvarez Hubert. 2022. "First Report on Microcystin-LR Occurrence in Water Reservoirs of Eastern Cuba, and Environmental Trigger Factors" Toxins 14, no. 3: 209. https://doi.org/10.3390/toxins14030209