Insights from the Structure of an Active Form of Bacillus thuringiensis Cry5B


Abstract
:1. Introduction
2. Results
2.1. Structure of Cry5B(27–698) with Its Functionally Required N-Terminal Extension
2.2. Cry5B (27–698) Is an Active Toxin
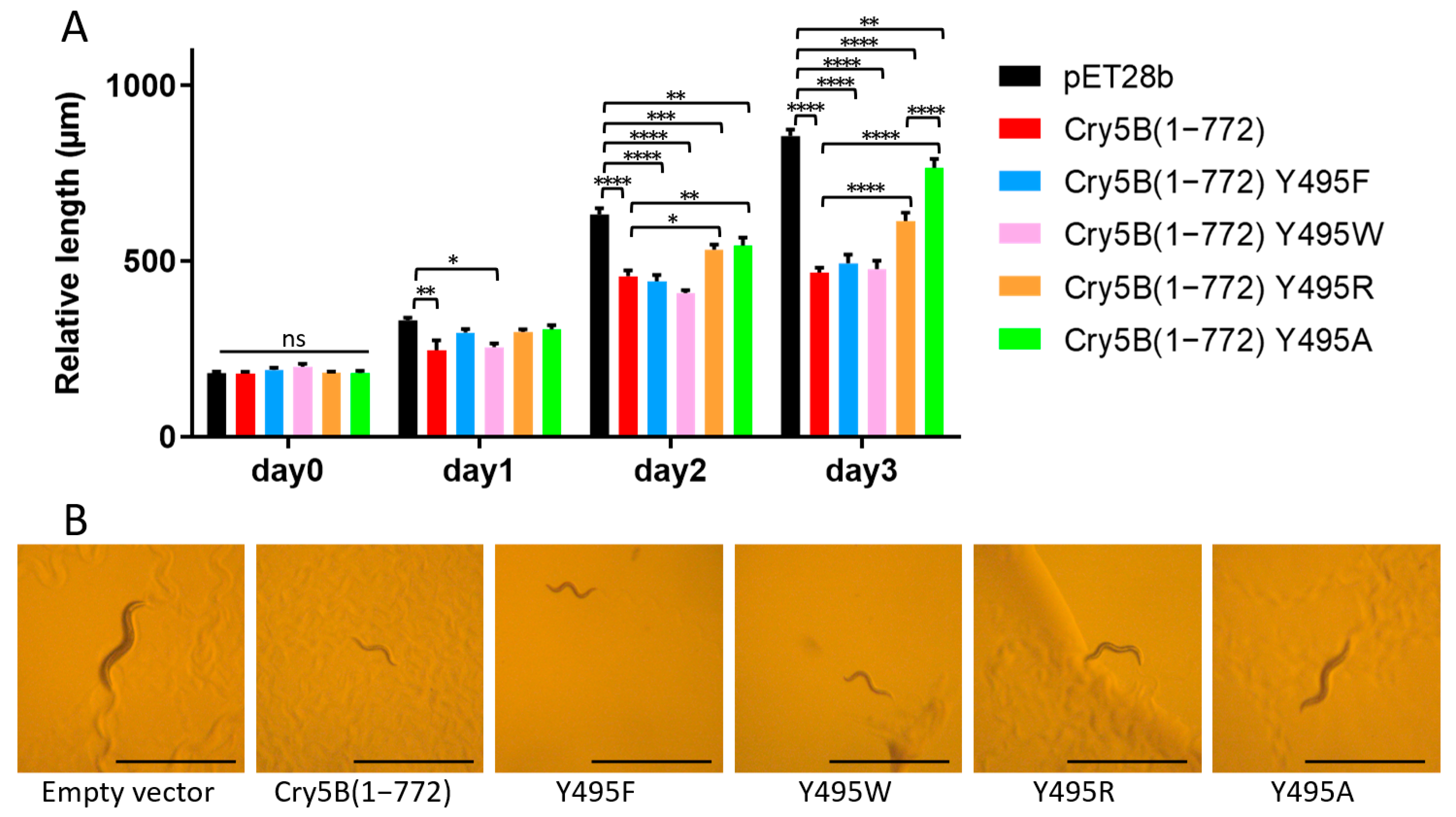
2.3. Identification of Tyr495 as a Residue with a Key Role in the Nematicidal Activity of Cry5B
3. Discussion
3.1. Computational Model for the N-Terminal Extension in Cry5B Tetramer
3.2. Cry5B N-Terminal Region (Residues 27–111) Contributes to Binding with Galactose
3.3. Comparison of the Cry5B(27–698) with Other Three-Domain Cry Proteins
4. Conclusions
5. Materials and Methods
5.1. Cloning of Cry5B Fragments and Mutants
5.2. Expression and Purification of Cry5B(1–772)
5.3. Crystallization and Data Collection
5.4. Phase Determination and Refinement
5.5. Cry5B Toxicity Study on C. elegans
5.5.1. General Preparation
5.5.2. Toxicity Test
5.5.3. Microscale Thermophoresis (MST)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tabashnik, B.E. Communal benefits of transgenic corn. Science 2010, 330, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Zaman, M.; Shaheen, T.; Irem, S.; Zafar, Y. Safe use of Cry genes in genetically modified crops. Environ. Chem. Lett. 2015, 13, 239–249. [Google Scholar] [CrossRef]
- Briefs, I. Global status of commercialized biotech/GM crops in 2017: Biotech crop adoption surges as economic benefits accumulate in 22 years. ISAAA Brief 2017, 53, 25–26. [Google Scholar]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, M.; Bungiro, R.D.; Harrison, L.M.; Bischof, L.J.; Griffitts, J.S.; Barrows, B.D.; Aroian, R.V. A purified Bacillus thuringiensis crystal protein with therapeutic activity against the hookworm parasite Ancylostoma ceylanicum. Proc. Natl. Acad. Sci. USA 2006, 103, 15154–15159. [Google Scholar] [CrossRef] [Green Version]
- Urban Jr, J.F.; Hu, Y.; Miller, M.M.; Scheib, U.; Yiu, Y.Y.; Aroian, R.V. Bacillus thuringiensis-derived Cry5B has potent anthelmintic activity against Ascaris suum. PLoS Negl. Trop. Dis. 2013, 7, e2263. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Abraham, A.; Gazzola, D.; Hu, Y.; Beamer, G.; Flanagan, K.; Soto, E.; Rus, F.; Mirza, Z.; Draper, A. Recombinant paraprobiotics as a new paradigm for treating gastrointestinal nematode parasites of humans. Antimicrob. Agents Chemother. 2021, 65, e01469-20. [Google Scholar] [CrossRef]
- Urban, J.F., Jr.; Nielsen, M.K.; Gazzola, D.; Xie, Y.; Beshah, E.; Hu, Y.; Li, H.; Rus, F.; Flanagan, K.; Draper, A. An inactivated bacterium (paraprobiotic) expressing Bacillus thuringiensis Cry5B as a therapeutic for Ascaris and Parascaris spp. infections in large animals. One Health 2021, 12, 100241. [Google Scholar] [CrossRef]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Griffitts, J.S.; Haslam, S.M.; Yang, T.; Garczynski, S.F.; Mulloy, B.; Morris, H.; Cremer, P.S.; Dell, A.; Adang, M.J.; Aroian, R.V. Glycolipids as receptors for Bacillus thuringiensis crystal toxin. Science 2005, 307, 922–925. [Google Scholar] [CrossRef] [Green Version]
- Hui, F.; Scheib, U.; Hu, Y.; Sommer, R.J.; Aroian, R.V.; Ghosh, P. Structure and glycolipid binding properties of the nematicidal protein Cry5B. Biochemistry 2012, 51, 9911–9921. [Google Scholar] [CrossRef] [PubMed]
- Meagher, J.L.; Winter, H.C.; Ezell, P.; Goldstein, I.J.; Stuckey, J.A. Crystal structure of banana lectin reveals a novel second sugar binding site. Glycobiology 2005, 15, 1033–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-Q.; Tan, A.; Voegtline, M.; Bekele, S.; Chen, C.-S.; Aroian, R.V. Expression of Cry5B protein from Bacillus thuringiensis in plant roots confers resistance to root-knot nematode. Biol. Control. 2008, 47, 97–102. [Google Scholar] [CrossRef]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Buzdin, A.; Revina, L.; Kostina, L.; Zalunin, I.; Chestukhina, G. Interaction of 65-and 62-kD proteins from the apical membranes of the Aedes aegypti larvae midgut epithelium with Cry4B and Cry11A endotoxins of Bacillus thuringiensis. Biochemistry (Moscow) 2002, 67, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Boonserm, P.; Davis, P.; Ellar, D.J.; Li, J. Crystal structure of the mosquito-larvicidal toxin Cry4Ba and its biological implications. J. Mol. Biol. 2005, 348, 363–382. [Google Scholar] [CrossRef] [PubMed]
- Thamwiriyasati, N.; Sakdee, S.; Chuankhayan, P.; Katzenmeier, G.; Chen, C.-J.; Angsuthanasombat, C. Crystallization and preliminary X-ray crystallographic analysis of a full-length active form of the Cry4Ba toxin from Bacillus thuringiensis. Acta Crystallogr. F 2010, 66, 721–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.; Wan, D.; Cheng, C.; Ye, X.; Sun, M. Nematode-specific cadherin CDH-8 acts as a receptor for Cry5B toxin in Caenorhabditis elegans. Appl. Microbiol. Biotechnol. 2018, 102, 3663–3673. [Google Scholar] [CrossRef] [PubMed]
- Rajamohan, F.; Cotrill, J.A.; Gould, F.; Dean, D.H. Role of domain II, loop 2 residues of Bacillus thuringiensis CryIAb δ-endotoxin in reversible and irreversible binding to Manduca sexta and Heliothis virescens. J. Biol. Chem. 1996, 271, 2390–2396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, M.S.; Liu, X.S.; Dean, D.H. Membrane insertion of the Bacillus thuringiensis Cry1Ab toxin: Single mutation in domain II block partitioning of the toxin into the brush border membrane. Biochemistry 2008, 47, 5814–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.-S.; Lin, H.-Y.; Ying, L.; Shu, C.-L.; Song, F.-P.; Zhang, J. The minimal active fragment of the Cry1Ai toxin is located between 36I and 605I. J. Integr. Agric. 2014, 13, 1036–1042. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, Z.; Song, F.; Shu, C.; Huang, D.; Zhang, J. Identification of the minimal active fragment of the Cry1Ah toxin. Biotechnol. Lett. 2011, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Battye, T.G.G.; Kontogiannis, L.; Johnson, O.; Powell, H.R.; Leslie, A.G. iMOSFLM: A new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr. D 2011, 67, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D 2006, 62, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Project, C.C. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D 1994, 50, 760–763. [Google Scholar]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkóczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.-W.; Jain, S.; McCoy, A.J. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. D 2019, 75, 861–877. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans (February 11, 2006), The C. elegans Research Community. In WormBook; Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 30 October 2022). [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cry5B(27–698) | |
| Data collection | |
| Wavelength (Å) | 0.99984 |
| Space group | P 4 21 2 |
| Cell dimensions: a, b, c (Å) | 114.4 114.4 263.4 |
| α, β, γ (°) | 90.0 90.0 90.0 |
| Resolution (Å) | 20 (4.5) |
| Rmerge | 0.274 (0.559) |
| I/σI | 2.2 (1.3) |
| Completeness (%) | 99.7 (100) |
| Redundancy | 5.3 (5.4) |
| Refinement | |
| No. reflections | 10,836 |
| Rwork/Rfree | 23.76/28.85 |
| No. atoms: Protein | 10,254 |
| Ligand/ion | 0 |
| Water | 0 |
| B-factors(Å2): Protein | 107.08 |
| R.m.s. deviations: Bond lengths (Å) | 0.002 |
| Bond angles (°) | 0.54 |
| Range of residues | 27–83, 88–107, 113–165, 173–214, 224–698 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wang, L.; Kotaka, M.; Lee, M.M.; Chan, M.K. Insights from the Structure of an Active Form of Bacillus thuringiensis Cry5B. Toxins 2022, 14, 823. https://doi.org/10.3390/toxins14120823
Li J, Wang L, Kotaka M, Lee MM, Chan MK. Insights from the Structure of an Active Form of Bacillus thuringiensis Cry5B. Toxins. 2022; 14(12):823. https://doi.org/10.3390/toxins14120823
Chicago/Turabian StyleLi, Jiaxin, Lin Wang, Masayo Kotaka, Marianne M. Lee, and Michael K. Chan. 2022. "Insights from the Structure of an Active Form of Bacillus thuringiensis Cry5B" Toxins 14, no. 12: 823. https://doi.org/10.3390/toxins14120823