1. Introduction
Cereal grains, as critical crop products, play a crucial role in the human diet and livestock feed due to their valuable contents such as carbohydrates, proteins, fatty acids and vitamins [
1]. The feed supply chain is a crucial element for all livestock production systems. According to the FEFAC [
2], within the European Union-28 (EU-28), approximately 475 million tons of feedstuffs and forages are consumed by livestock each year. For example, France ranks the 1st among European countries in the production of durum wheat.
About one fourth of global agricultural products and half of cereal grain samples (or derived products) used for livestock feed are contaminated with mycotoxins in Europe and worldwide [
3,
4,
5]. Fumonisins, trichothecenes and zearalenone (ZEN) are the most commonly occurring fusariotoxins in cereal grains and animal feed [
6,
7]. Regarding finished feed, FBs (91%), deoxynivalenol (82%), ZEN (35%), aflatoxins (18%), T2 (4%) and ochratoxin A (3%) are the most frequently found mycotoxins under warm climate conditions [
8,
9]. Both ZEN and deoxynivalenol (DON) are produced in the field and upon postharvest storage (grain, silage), whereas T2 is produced only upon seed storage. All 3 are toxic to livestock, particularly monogastric animals which do not possess rumen with mycotoxin-degrading microorganisms [
10]. ZEN interacts with estrogen receptors, inducing reproduction troubles (abortion, impairment of fertility etc.) [
11,
12]. T2 inhibits protein and DNA synthesis, and causes emaciation, diarrhea, immunosuppression, skin necrosis and haemorrhage [
13,
14]. DON decreases weight gain and vaccination response, and causes vomiting and rejection of feed [
10,
15,
16].
Despite efforts to control fungal contamination, including the implementation of good agricultural and manufacturing practices, extensive mycotoxin contamination has been reported to occur in feed and food. At the field level, crop rotation, variety selection, irrigation, tillage, fungicide spray may allow a reduction of toxin-producing fungi or of mycotoxin prevalence. However, the presence of fungi or toxins within cereal products still remains a problem.
Upon storage of cereal products, addition of adsorbants such as clays (e.g., Mycofix
®, Biomin or Amadeite
®, Olmix, Brehan, France), active charcoal or seaweed-extracted biopolymers may reduce toxin bioavailability. Besides, a number of chemical or biological treatments have been proposed to transform or scavenge mycotoxins [
17]. They include ammoniation or gamma irradiation for seed decontamination but are rather inefficient against fusariotoxins or inapplicable by breeders [
18]. Moreover, the use of probiotics like micro-organisms and derived products as detoxifying agents [
19] has been proposed. However, these probiotics hardly affect some mycotoxins like DON [
20]. Moreover, most of the results are highly variable depending on the method and the mycotoxin used. Therefore, an efficient natural food solution protecting livestock against deleterious effects of mycotoxins is still needed.
Plants have long been a source of medicines and phytoproducts are widely used in the food industry, nutraceuticals, and medicine. The latest trend of returning to natural sources for health has generated a wide interest for bioactive compounds from plants. For instance, the antioxidant plants have gained a huge amount of attention during the last two decades and the supplementation of exogenous antioxidants appears as a promising way to improve the undesirable effects of oxidative stress. In that context, halophytes represent a renewable source of bioactive molecules for food, cosmetics, and pharmaceutical uses. These plants have been poorly studied until the last decade, and recent works showed that they exhibit numerous biological effects thanks to their high levels of secondary metabolites [
21,
22]. Among them, polyphenols display significant antioxidant activities and a wide spectrum of medicinal properties [
23,
24,
25].
At a cellular level, one of the main effects induced by mycotoxins on consumers is a significant oxidative stress and inflammatory process [
26]. Since halophytic plants have a constitutively strong antioxidant defense, we investigated the potent protective effect of edible halophytes against mycotoxin contamination in animal cells. Such property was studied along with the antioxidant and anti-inflammatory potential of these plants to assess the possible mechanisms of this cytoprotective action.
3. Discussion
Mycotoxins and their control generate a considerable economic impact not only to the feed producer but also to intermediaries such as elevators, buyers of grains and food processors. Previous studies have described the toxicological effects of
Fusarium toxins in farm animals [
27,
28]. In ruminants, the animal species least sensitive to DON, feed refusal syndrome was reported in cows after consumption of wheat contaminated with 6.4 mg DON/kg feed during 10 weeks [
29]. In pigs, the symptoms include vomiting, diarrhea, leukopenia, hemorrhage, shock and death. Toxic effects of T2 toxin are usually manifested in the form of Alimentary Toxic Aleukia (ATA). Ruminants are known to be relatively resistant to T2 toxin in comparison to monogastric animals. The toxic effects of ZEN on cows are associated with vulvar hypertrophy and ovarian atrophy. Ruminants are less susceptible than pigs to this toxin due to ruminal microbiota’s potential to transform ZEN to its hydroxyl metabolites, α-zearalenol and β-zearalenol. Some solutions have been described as addition of various inorganic ligands such as aluminosilicates, hydrated sodium calcium aluminosilicate and zeolite prevented the adverse effects of aflatoxins on swine performance. However, this anti-mycotoxin ability of these products could not prevent the absorption of
Fusarium toxins in the intestine of pigs [
30]. Than et al. [
31] determined the adverse effects of four commercial anti-mycotoxin additives in preventing the negative effects of DON. Anti-mycotoxin additives were ineffective at improving the growth performance at dietary concentrations used.
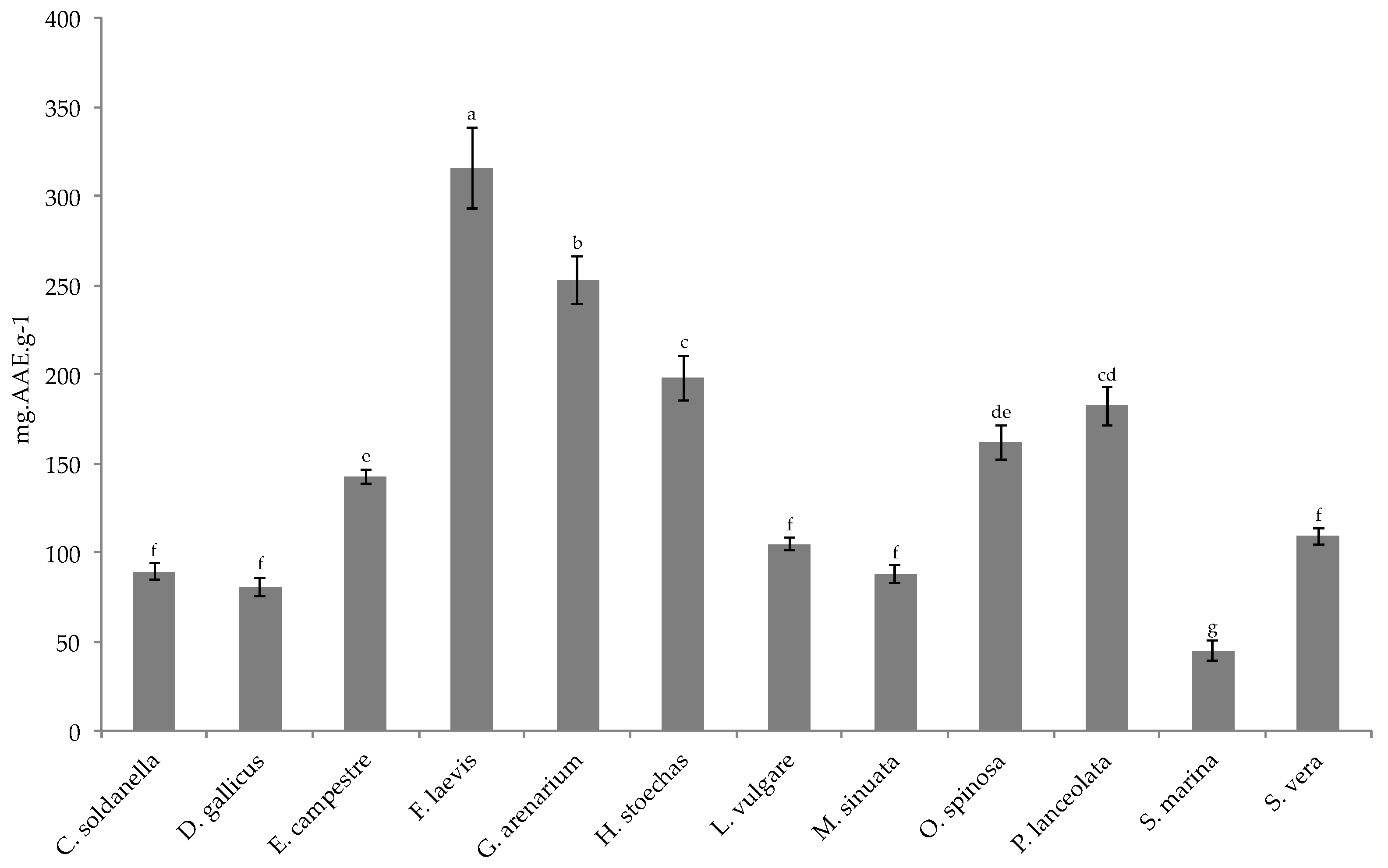
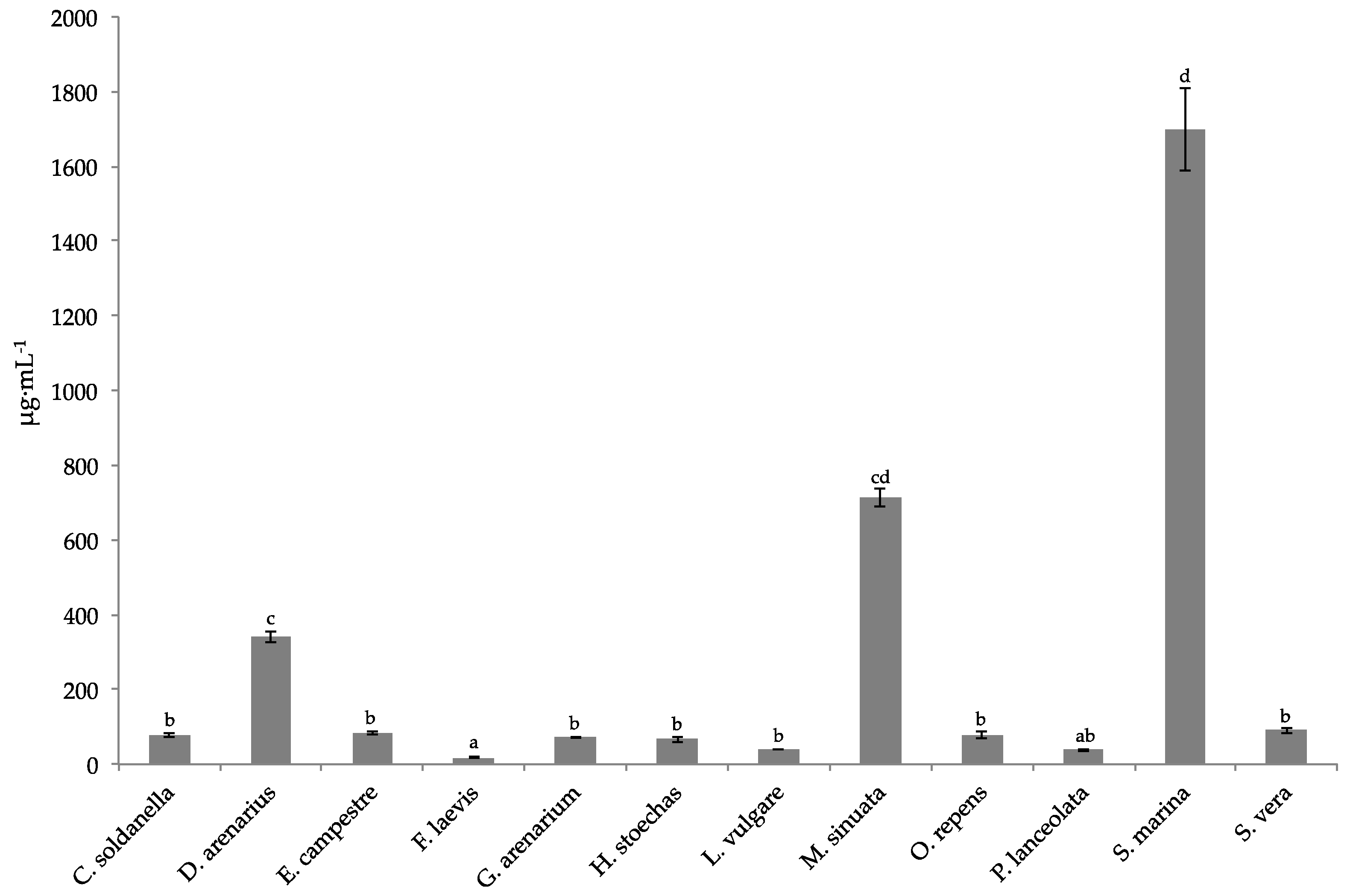
Over the past decades, the search for natural products in plants has led to the discovery of a number of biologically active substances, particularly secondary metabolites, which confer plants with a number of biological activities. Of the twelve species studied here, nine exhibited very strong antioxidant capacities (IC
50 < 100 μg·mL
−1 for DPPH radical and TAC higher than 100 mgAAEg-1DW), reinforcing the idea that halophytes are constitutively equipped with an efficient antioxidant system to cope with harmful coastal environments [
21,
32]. Moreover, it has already reported that antioxidant compounds may alleviate mycotoxin toxicity [
33]. Here, the studied halophytes showed a strong protective action on MDBK and IPEC-J2 cells against ZEN or T2 toxin contamination (restoring about 75% of cell viability at 10 μg·mL
−1). However, ranking of antioxidant activities of the studied species did not fully match with that of cytoprotective action upon mycotoxin exposure, suggesting that cell protection against intoxication would require additional mechanisms along with antioxidant action. In particular, every extract failed to protect animal cells against DON intoxication though they all showed antioxidant effects. The main toxic mechanism of DON is well-known. DON binds to ribosomes thereby destroying their structure. It is also reported to interfere with the peptidyl transferase active center existing on the 60S subunit of ribosomes thus inhibiting normal protein synthesis [
15]. At the molecular level, DON induces the global impact on the transcriptome and triggers prototypical signaling pathways linked to immunity and inflammation, including the p38 mitogen-activated protein kinases (p38-MAPK) and the NF-κB [
31]. Same effects were described on IPEC-J2, DON exposure alters the phosphorylation states and sites of multiple proteins found in differentiated intestinal epithelial cells [
34]. On MDBK, exposure to DON suggested decreased protein synthesis [
35]. Further investigations are necessary to analyze this intriguing mechanism of DON in the intestine and renal cell models.
This preliminary study allowed us to select the most efficient extract in terms of antioxidant capacity and cytoprotective effects, namely that of Galium arenarium, for further analyses addressed to identify the potent bioactive compounds. Moreover, such bioguided study was performed on MDBK cells, a relevant model to study bovin nephrotoxicity of mycotoxin and since it possessed a shorter generation time.
Several studies have investigated chemical composition and assessed biological properties of crude extracts from
Rubiaceae [
36,
37]. However, no studies so far have investigated thoroughly the molecules responsible for these activities. Here, enriched fractions of
G. arenarium extract have been evaluated for the first time for antioxidant, anti-inflammatory and cytoprotective activities, particularly in the context of mycotoxin exposure.
The crude extract of
G. arenarium exhibited a strong antioxidant activity, confirming previous works with other bioassays [
38]. The bio-guided fractionation of this extract allowed us to recover and concentrate the antioxidant activity into four fractions, eluted with 20 to 80% methanol. Accordingly, these fractions exhibited a DPPH IC
50 lower than 40 µg·mL
−1 and total antioxidant capacity higher than 300 mg AAE·g
−1 DW. Interestingly, these fractions all exhibited a strong protective action on bovine and porcine cells upon T2 or zearalenone exposure, suggesting that this effect is at least due to the relevant phenolic compounds present in these fractions, as it has been already reported [
39,
40,
41]. Accordingly, Montibus et al. [
42] showed that the phenolic composition of a maritime pine extract, and particularly methylated compounds, would be essential for its antifungal and anti-mycotoxin properties.
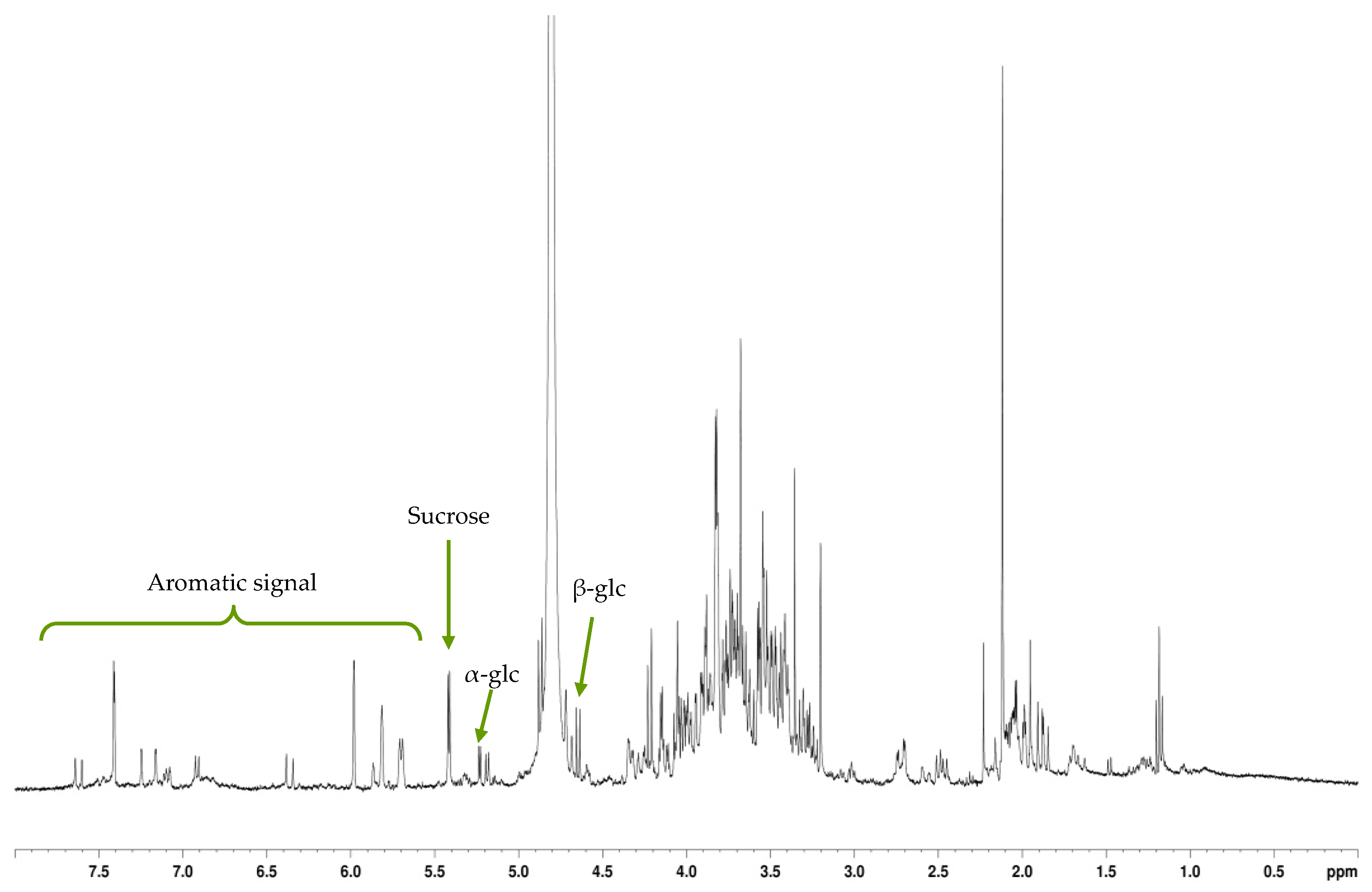
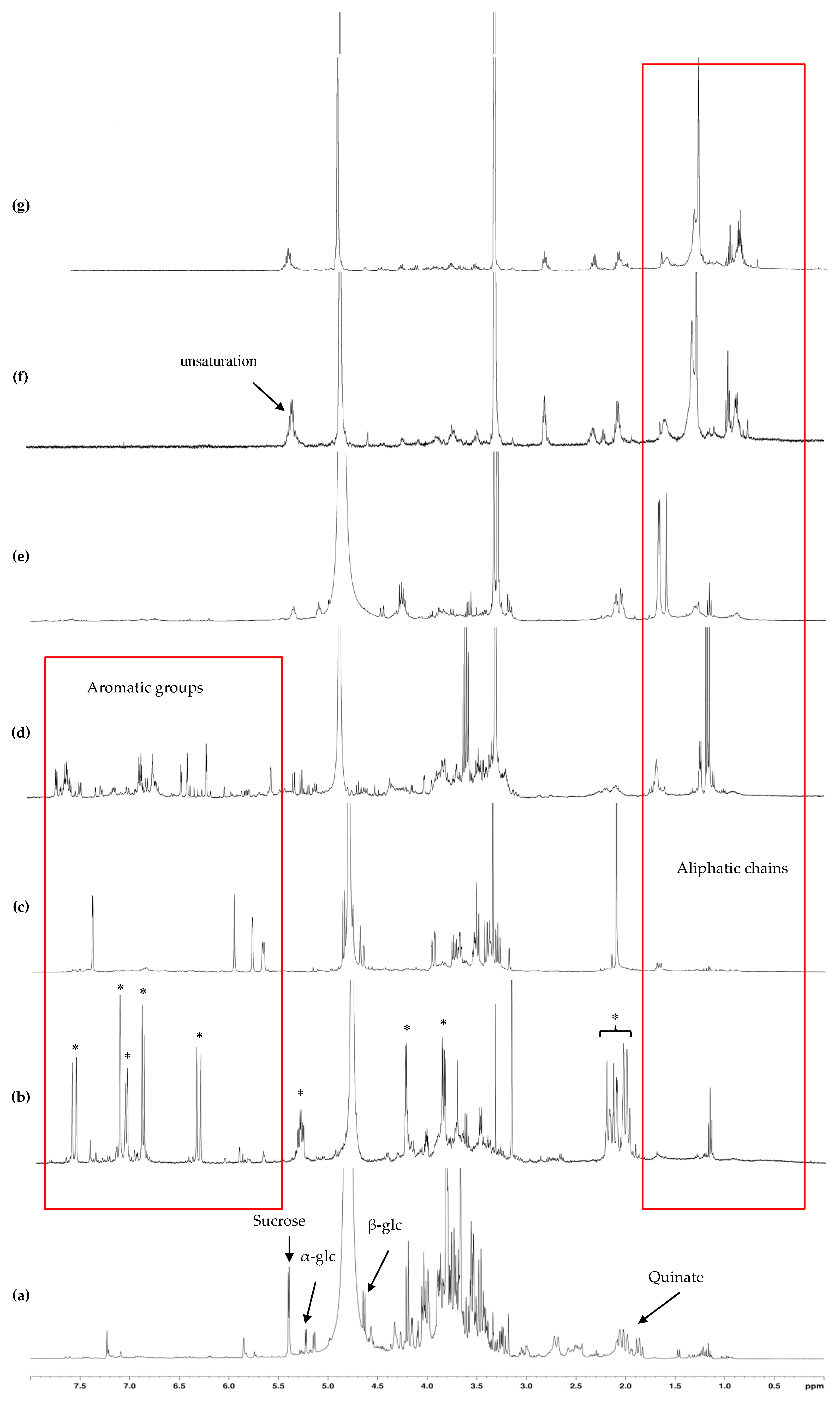
The fraction eluted with 20% MeOH exhibited a high phenolic content and, correlatively, a strong antioxidant activity. NMR analyses of this fraction showed that its major constituents were free quinic acid and its bound form chlorogenic acid, a well-known antioxidant compound already reported in
Galium genera [
43,
44].
The fraction eluted with 40% MeOH exhibited the strongest antioxidant capacity. 2D-NMR analyses of this fraction showed the predominance of the iridoid glycoside asperuloside. Such compound has already been reported in some
Galium species [
37,
45] but never in
G. arenarium. It has been related to a number of pharmacological properties as an anticancer [
46], anti-inflammatory and anti-obesity [
47], anti-viral, anti-malarial, anti-protozoal, anti-hypertensive, immunomodulatory, and antioxidant agent [
48]. Therefore, we suggest that the abundance of this iridoid is responsible for the very strong antioxidant and anti-inflammatory activities of the MeOH
40 fraction. Moreover, it could be involved in the cytoprotective action of
G. arenarium MeOH
40 fraction.
The fraction eluted with 60% MeOH exhibited the strongest DPPH-scavenging activity (
Table 7). The major compounds characterized in this fraction were glycosylated phenolic compounds. Considering the strong antioxidant power of phenolic glycosides [
49], these compounds are likely to be responsible for the strong antioxidant activity of this fraction. Moreover, the strong anti-inflammatory and cytoprotective actions of MeOH
60 fraction could be due to such phenolic glycosides, as it has been already reported [
34]. Besides, these activities could also be attributed to anthraquinones, as reported in several
Galium species [
50,
51].
The fraction eluted with 80% MeOH exhibited no signals of aromatic compounds but still higher antioxidant activities than those of the crude extract. Noteworthily, this fraction exhibited a remarkable cytoprotective activity. Along with the 1D- and 2D- NMR analyses, such activities could be due to terpenoids as reported by Grassman [
52].
The last two fractions showed low antioxidant activities but still a strong cytoprotective action against mycotoxin exposure (particularly T2 toxin and zearalenone), indicating that cytoprotective action against mycotoxin intoxication does not only proceed through antioxidant pathways. It is supposed here that another mechanism of cell protection like anti-inflammatory process would also provide cell protection effect. Moreover, their NMR spectra showed several strong signals in the aliphatic region, which could be attributed to phytosterols such as sitosterol or campesterol, as already reported [
53].
5. Materials and Methods
5.1. Chemicals, Culture Media and Supplements
Madin-Darby Bovine Kidney (MDBK) and intestinal porcine enterocyte (IPEC-J2) cell lines were purchased from DSMZ, (Braunschweig, Germany). Cell culture medium (DMEM, EMEM, Eagle’s Minimum Essential Medium), horse fœtal serum (HSF), Folin-Ciocalteu phenol reagent, DPPH, all standards and solvents used for chemical analyses were supplied by Sigma Aldrich (St. Louis, MO, USA). Concerning mycotoxins, deoxynivalenol (DON) (MW = 296.35 g/mol, purity ≥ 98%) is produced by Fusarium sp. T2 (MW = 466.5 g/mol, purity ≥ 98%) and zearalenone (ZEN) (MW = 320.38 g/mol, purity ≥ 98%) have a not specified origin. All mycotoxins were purchased from Sigma Aldrich.
5.2. Cell Culture
MDBK and IPEC-J2 cells were maintened at 37 °C under 5% CO2 and water saturated atmosphere. IPEC-J2 is a non-transformed, permanent intestinal cell line. The IPEC-J2 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) and supplemented with 1% HEPES, 10% fetal bovine serum (FBS), 1% penicillin/streptomycin. MDBK were cultured in medium (EMEM) containing 4.5 g/L glucose for cell growth was added with 10% horse fœtal serum (HSF) and 1% penicillin/streptomycin to support cell division.
5.3. Plant Sampling
Aerial parts of twelve halophytic species were collected along the Brittany shoreline of Finistère (France):
Convolvulus soldanella L. (Convolvulaceae),
Dianthus gallicus Pers. (Caryophyllaceae),
Eryngium campestre L. (Apiaceae),
Frankenia laevis L. (Frankeniaceae),
Galium arenarium Loisel. (Rubiaceae),
Helichrysum stoechas (L.) Moench (Asteraceae),
Limonium vulgare Mill. (Plumbaginaceae),
Matthiola sinuata (L.) R.Br. (Brassicaceae),
Ononis spinosa subsp.
procurrens Wallr. (Rosaceae),
Plantago lanceolata (L.) (Plantaginaceae),
Spergularia marina (L.) (Caryophyllaceae), and
Suaeda vera Forssk. ex J.F.Gmel. (Amaranthaceae). These species belong to eleven different families (
Table 8) and grow in three different habitats (cliffs, sand dune, salt marsh). One sample of each species was deposited at the herbarium of University of Brest. The aerial parts of every sample were rinsed and the leaves were frozen and subsequently freeze-dried. Each sample was then ground to a fine powder in an AE 200 blender (Mettler, Viroflay, France), before extraction and analyses.
5.4. Extraction of Metabolites from Halophyte Leaves
About 500 mg of dry powder were homogenized with 5 mL water/ethanol (1:2) under magnetic stirring at 4 °C for 20 min. After centrifugation of the mixture (15 min at 4 °C, 4000× g), the resulting pellet was extracted twice following the same protocol. The supernatants were collected, pooled and filtered over glass wool. The obtained extract was concentrated by rotary evaporation at 40 °C and resuspended in either DMSO (for cell treatments) or 50% ethanol (for chemical analyses).
5.5. Evaluation of Plant Extract Effects on Cell Lines
5.5.1. Cell Treatments
Cells (MDBK and IPEC-J2) were seeded at 5.105 cells/mL in 96 well plates. After one day of culture, plant extracts were added (0.1–10 μg·mL−1). Twenty-four hours later, cells were incubated with cytotoxic concentration of mycotoxins (DON, ZEN or T2) during 48 h before assay analysis.
5.5.2. Measurement of the Epithelial Barrier Function
Measurement of the trans-epithelial electrical resistance (TEER) was performed to evaluate monolayer integrity and possible damage of the cellular monolayer during the experiments. TEER was measured under sterile conditions using the Millicell ERS system (Millipore Co., Bedford, MA, USA) according to the manufacturer’s instructions. TEER values were recorded every 24 h and expressed as Ω × cm2 on the basis of the following equation: TEER = (R − Rb) × A, where R is the resistance of filter insert with cells, Rb is the resistance of the filter alone and A is the growth area of the filter in cm2.
5.5.3. Evaluation of Cell Viability
The cytotoxic effect of polar and apolar extracts was studied on MDBK and IPEC-J2 cells. Thus, cytotoxicity was evaluated using the CellTiter 96AQueous One cell proliferation assay (Promega, Madison, WI, USA), as described by Hymery et al. [
54] (2014). This colorimetric method determines cell viability based on the reduction of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) to formazan by mitochondrial dehydrogenases in viable cells. After time of incubation in the presence of samples, cells were washed with PBS, resuspended in 100 μL of the same buffer, and seeded in 96-well plates at 37 °C, under 5% CO
2 atmosphere and 100% humidity. Then, 20 μL of CellTiter 96AQueous One solution were added to each well and the cells were further incubated for 3 h at 37 °C, under 5% CO
2 atmosphere and 100% humidity. The absorbance was measured at 490 nm. Cytotoxicity was expressed as the concentration of samples inhibiting cell growth compared with the control (cells treated with 1% DMSO). All tests and analyses were run in triplicate and averaged.
5.5.4. Measurements of Cytokine and Interleukin Productions
MDBK and IPEC-J2 cell lines were incubated for 24 h in the presence of plant extracts. To evaluate a possible antiinflammatory effect of plant extract, IL-8 and TNF-α were monitored. The amount of Tumor Necrosis Factor alpha (TNF-α) and IL-8 in the supernatants of plant extract treated cell cultures was then quantified using ELISA kits (Promocell, Heidelberg, Germany). Three plant extracts were chosen from cytotoxicity results.
5.6. Measurement of Antioxidant Activities in Plant Extracts
5.6.1. Total antioxidant Capacity (TAC)
Total antioxidant capacity of ethanolic extracts was evaluated through the assay of a green phosphate/Mo5+ complex according to the method described by Prieto et al. [
55]. An aliquot (0.1 mL) of diluted samples was combined with 1 mL of reagent solution (0.3 N sulfuric acid, 28 mM sodium phosphate and 4 mM ammonium molybdate). Methanol was used instead of the sample for the blank. The tubes were incubated in a boiling water bath for 90 min. Then, the samples were cooled to room temperature and the absorbance was measured at 695 nm against blank in UV-Visible spectrophotometer (Anthelie Advanced 2, Secomam, Champigny sur Marne, France). Antioxidant capacity was expressed as mg ascorbic acid equivalent per gram dry weight (mg AAE g
−1 DW). All samples were analyzed in triplicate.
5.6.2. DPPH Scavenging Activity
The scavenging activity of the stable 1,1-diphenyl-2-picrylhydrazyl (DPPH) free radical was determined by the method of Marwah et al. [
56]. Briefly, the reaction medium contained 100 μL of 100 μM DPPH violet solution in ethanol and 100 μL of plant extracts at different concentrations (or water for the control). The reaction mixture was incubated in the dark for 15 min and the absorbance was recorded at 517 nm on a microtiter reader (Multiskan EAR 400, Labsystems, Thermo Scientific, Madison, WI, USA). The assay was carried out in triplicate and butylated hydroxytoluene (BHT) was used as a positive control. The decrease in absorbance upon addition of test samples was used to calculate the inhibition percentage (%IP) of DPPH radical, following the equation:
where A
c and A
s are the absorbances of the control and the test sample, respectively. From a plot of concentration against %IP, a linear regression analysis was performed to determine the antiradical activity, as expressed by the IC
50 (extract concentration resulting in a 50% inhibition) value for each sample.
5.7. Fractionation of Plant Extracts
Fractionation of Galium arenarium raw extract was performed by solid-liquid partition chromatography on C18-bound silica gel. The elution of polar compounds was made with increasing methanol concentrations (successively 0, 20, 40, 60, 80, 100%) and finally by ethanol and then by ethyl acetate. The fractions were then concentrated by rotary evaporation at 40 °C and resuspended in the corresponding solvent.
5.8. Solute Purification
When necessary, purification from one fraction was performed by HPLC using a Shimadzu UFLC XR device equipped with a PDA detector (SPD-M20A, Shimadzu, Kyoto, Japan). A Spherisorb ODS2 column (5 μm, 250 × 4.6 mm, Waters, Milford, MA) was used for solute separation, and the mobile phase consisted of a mixture of acetonitrile 100% (A) and ultrapure water (B). The following linear gradient was applied: t = 0 min 100% B; t = 10 min 100% A; t = 12 min 100% A; t = 15 min 100% B. Compounds were detected at 254 nm and collected. When needed, some of them were submitted to acid hydrolysis treatment (1 N HCl, 110 °C for 1 h) before structural elucidation.
5.9. NMR Analyses
For bioactive compound characterization, an aliquot of G. arenarium crude extract and each fraction was concentrated by rotary evaporation at 35 °C, and the dry residue was solubilized in deuterated-water (D2O) or methanol (MeOD) for NMR analyses. 1H- NMR spectra were obtained using a Brüker Avance DRX-400 spectrometer (400 MHz), equipped with a 5 mm TBI probe (1H, X, 31P) with z gradient (Brüker, Rheinstetten, Germany). A typical 1D 1H-NMR spectrum consisted of 32 scans. The determination of major solutes present in sea fennel extract or fractions was made on NMR spectra in comparison with external standards. All 13C (J-mod) and 2D Homo- and heteronuclear NMR analyses (COSY, HMBC, HMQC experiments) were performed on a Brüker Avance III HD500 spectrometer equipped with a 5 mm TCI cryoprobe (1H, 13C, 15N) with z gradient. Data analysis was performed using TopSpin® software, 4.0 (Brüker).
5.10. Mass Spectrometry Analyses
Mass spectrometry analysis was performed using an Autoflex III smartbean spectrometer (Bruker Daltonics, Billerica, MA, USA). Datas were acquired with Flexcontrol software and analyzed with FlexAnalysis software. The experiments were carried out in positive reflectron mode over a mass range of 0 to 1500 m/z. The mixture (1 μL of product with 1 μL of matrix) was deposited on a metal plate. The matrix used is of the 4-hydroxy-α-cinnamic Acid (HCCA) type at 10 mg·mL−1 in 60% acetonitrile and 40% 0.1% TFA.
5.11. Statistical Analyses
All extractions and assays were conducted in triplicate. Results were expressed as mean ± standard deviation (SD), and the means were compared by using one-way analysis of variance (ANOVA) followed by Duncan’s multiple range tests performed by the ‘Statistica v. 5.1′ software (Statsoft, Tulsa, OK, USA). Differences between individual means were deemed to be significant at p < 0.05. For antioxidant bioassays, the IC50 values were calculated by the sigmoidal fitting of the data using the GraphPad Prism v. 5.0 program (GraphPad Software, San Diego, CA, USA).

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}