Altered Skeletal Muscle Fatty Acid Handling in Subjects with Impaired Glucose Tolerance as Compared to Impaired Fasting Glucose

Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Study Design
2.2.1. Hyperinsulinemic-Euglycemic Clamp
2.2.2. Postprandial Study
2.3. Biochemical Analyses
2.4. Skeletal Muscle Lipids
2.5. Skeletal Muscle mRNA Expression
2.6. Calculations
2.7. Statistical Analysis
3. Results
3.1. Subjects’ Characteristics
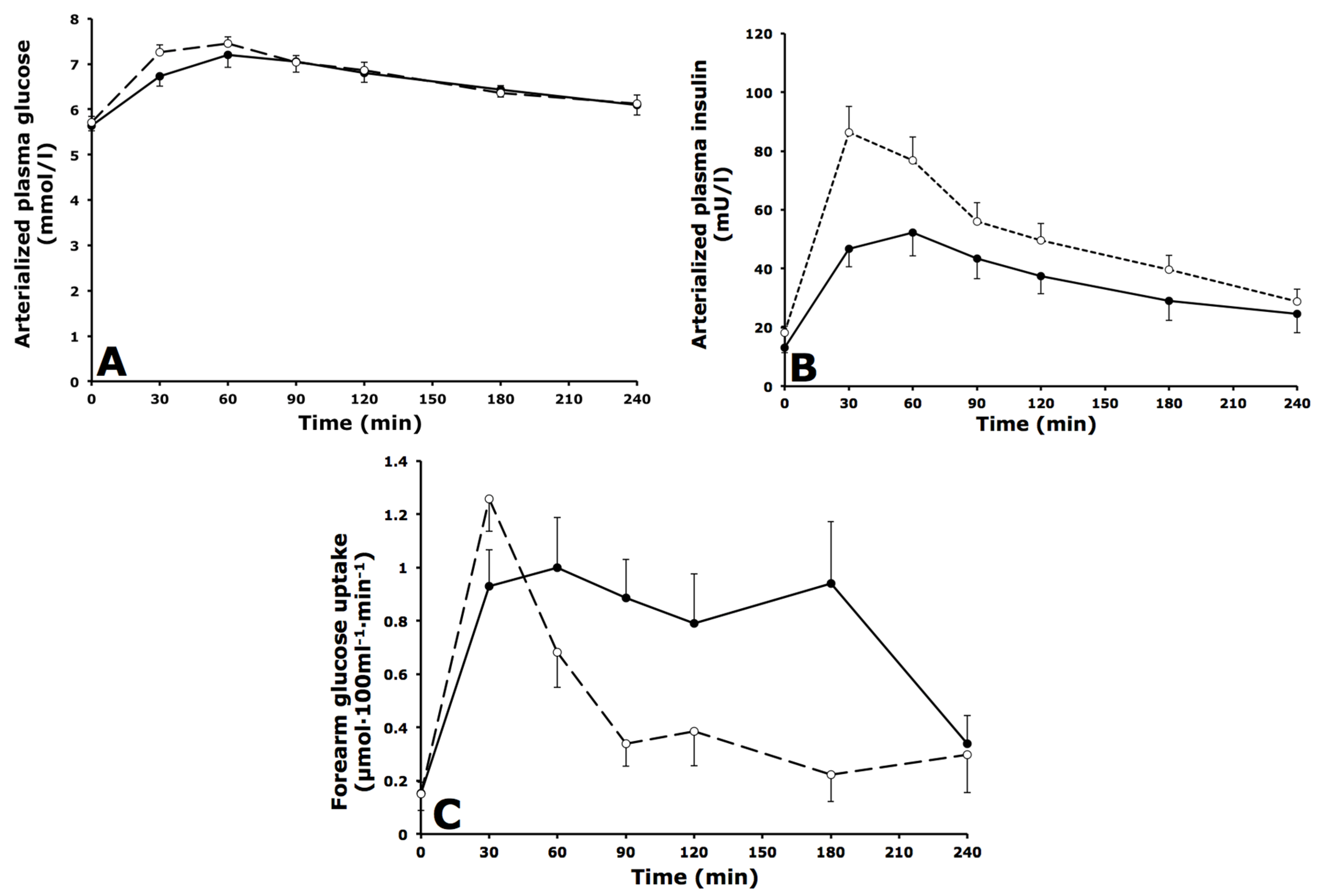
3.2. Arterialized Metabolites, Forearm Muscle Metabolism and Forearm Blood Flow
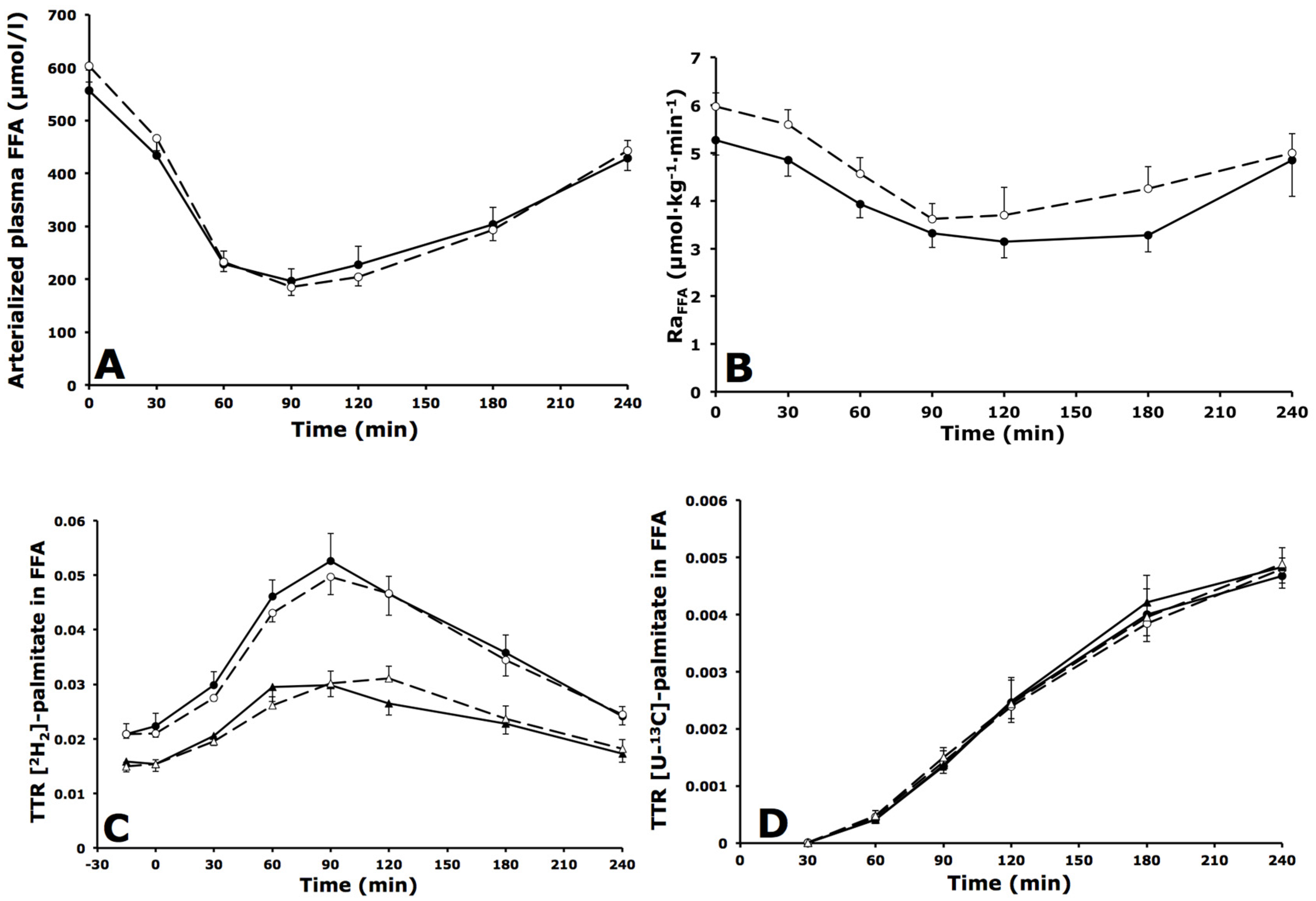
3.3. Whole-Body and Forearm Muscle FFA Metabolism
3.3.1. Whole-Body FFA Metabolism
3.3.2. Forearm Muscle FFA Metabolism
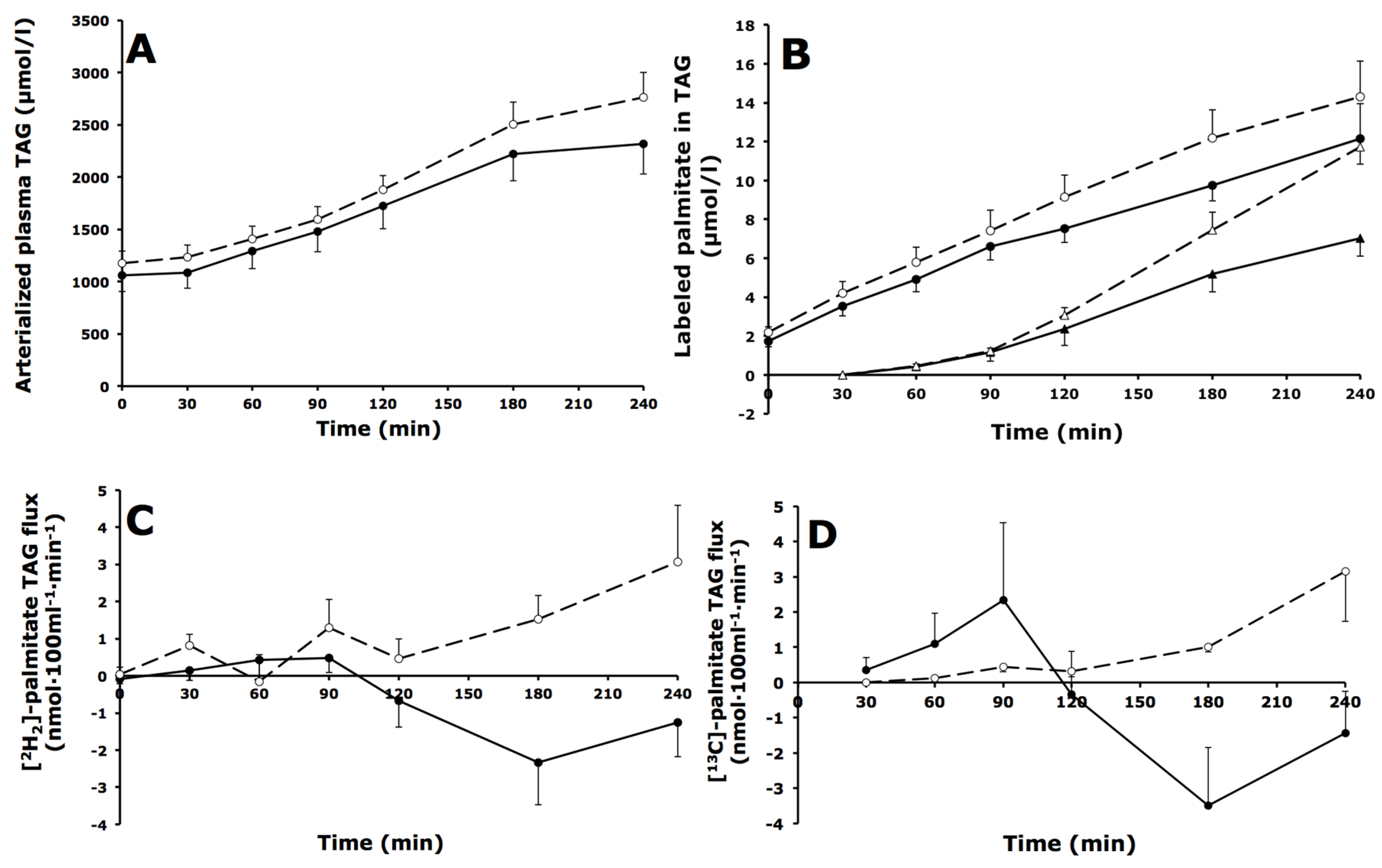
3.4. Whole-Body and Forearm Muscle TAG Metabolism
3.4.1. Whole-Body TAG Metabolism
3.4.2. Forearm Muscle TAG Metabolism
3.5. Intramuscular Lipid Metabolism
3.5.1. Lipid Content
3.5.2. Lipid Composition
3.6. Skeletal Muscle mRNA Expression
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DAG | Diacylglycerol |
| FA | fatty acid |
| FBF | forearm blood flow |
| FFA | free fatty acids |
| FSR | Fractional Synthetic Rate |
| HbA1c | Glycated hemoglobin |
| IGM | Impaired Glucose Metabolism |
| IFG | Impaired Fasting Glucose |
| IGT | Impaired Glucose Tolerance |
| LPL | lipoprotein lipase |
| MUFA | mono-unsaturated fatty acid |
| OGTT | Oral Glucose Tolerance Test |
| PL | phospholipid |
| PUFA | poly-unsaturated fatty acid |
| Ra | rate of appearance |
| SFA | saturated fatty acid |
| TAG | Triacylglycerol |
| T2DM | type 2 diabetes mellitus |
| TTR | Tracer/Tracee Ratio |
| VLDL | very-low density lipoprotein |
References
- Corpeleijn, E.; Mensink, M.; Kooi, M.E.; Roekaerts, P.M.; Saris, W.H.; Blaak, E.E. Impaired skeletal muscle substrate oxidation in glucose-intolerant men improves after weight loss. Obesity 2008, 16, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Mensink, M.; Blaak, E.E.; van Baak, M.A.; Wagenmakers, A.J.; Saris, W.H. Plasma free fatty acid uptake and oxidation are already diminished in subjects at high risk for developing type 2 diabetes. Diabetes 2001, 50, 2548–2554. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N. Adipose tissue as a buffer for daily lipid flux. Diabetologia 2002, 45, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H. The role of adipose tissue dysfunction in the pathogenesis of obesity-related insulin resistance. Physiol. Behav. 2008, 94, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Coen, P.M.; Goodpaster, B.H. Role of intramyocelluar lipids in human health. Trends Endocrinol. Metab. 2012, 23, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Corpeleijn, E.; Saris, W.H.; Blaak, E.E. Metabolic flexibility in the development of insulin resistance and type 2 diabetes: Effects of lifestyle. Obes. Rev. 2009, 10, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Stinkens, R.; Goossens, G.H.; Jocken, J.W.; Blaak, E.E. Targeting fatty acid metabolism to improve glucose metabolism. Obes. Rev. 2015, 16, 715–757. [Google Scholar] [CrossRef] [PubMed]
- Blaak, E.E.; van Aggel-Leijssen, D.P.; Wagenmakers, A.J.; Saris, W.H.; van Baak, M.A. Impaired oxidation of plasma-derived fatty acids in type 2 diabetic subjects during moderate-intensity exercise. Diabetes 2000, 49, 2102–2107. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.E.; Goodpaster, B.; Wing, R.R.; Simoneau, J.A. Skeletal muscle fatty acid metabolism in association with insulin resistance, obesity, and weight loss. Am. J. Physiol. 1999, 277, E1130–E1141. [Google Scholar] [PubMed]
- Schrauwen, P.; Schrauwen-Hinderling, V.; Hoeks, J.; Hesselink, M.K. Mitochondrial dysfunction and lipotoxicity. Biochim. Biophys. Acta 2010, 1801, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Moro, C.; Bajpeyi, S.; Smith, S.R. Determinants of intramyocellular triglyceride turnover: Implications for insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E203–E213. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Hunerdosse, D.M.; Kerege, A.; Playdon, M.C.; Perreault, L. Localisation and composition of skeletal muscle diacylglycerol predicts insulin resistance in humans. Diabetologia 2012, 55, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Jocken, J.W.; Goossens, G.H.; Boon, H.; Mason, R.R.; Essers, Y.; Havekes, B.; Watt, M.J.; van Loon, L.J.; Blaak, E.E. Insulin-mediated suppression of lipolysis in adipose tissue and skeletal muscle of obese type 2 diabetic men and men with normal glucose tolerance. Diabetologia 2013, 56, 2255–2265. [Google Scholar] [CrossRef] [PubMed]
- Van Hees, A.M.; Jans, A.; Hul, G.B.; Roche, H.M.; Saris, W.H.; Blaak, E.E. Skeletal muscle fatty acid handling in insulin resistant men. Obesity 2011, 19, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Bakke, S.S.; Moro, C.; Nikolic, N.; Hessvik, N.P.; Badin, P.M.; Lauvhaug, L.; Fredriksson, K.; Hesselink, M.K.; Boekschoten, M.V.; Kersten, S.; et al. Palmitic acid follows a different metabolic pathway than oleic acid in human skeletal muscle cells; lower lipolysis rate despite an increased level of adipose triglyceride lipase. Biochim. Biophys. Acta 2012, 1821, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Gaster, M.; Rustan, A.C.; Beck-Nielsen, H. Differential utilization of saturated palmitate and unsaturated oleate: Evidence from cultured myotubes. Diabetes 2005, 54, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Pinnamaneni, S.K.; Eo, S.J.; Cho, I.H.; Pyo, J.H.; Kim, C.K.; Sinclair, A.J.; Febbraio, M.A.; Watt, M.J. Saturated, but not n-6 polyunsaturated, fatty acids induce insulin resistance: Role of intramuscular accumulation of lipid metabolites. J. Appl. Physiol. 2006, 100, 1467–1474. [Google Scholar] [CrossRef] [PubMed]
- Montell, E.; Turini, M.; Marotta, M.; Roberts, M.; Noe, V.; Ciudad, C.J.; Mace, K.; Gomez-Foix, A.M. Dag accumulation from saturated fatty acids desensitizes insulin stimulation of glucose uptake in muscle cells. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E229–E237. [Google Scholar] [PubMed]
- Blaak, E.E.; Wagenmakers, A.J.; Glatz, J.F.; Wolffenbuttel, B.H.; Kemerink, G.J.; Langenberg, C.J.; Heidendal, G.A.; Saris, W.H. Plasma ffa utilization and fatty acid-binding protein content are diminished in type 2 diabetic muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E146–E154. [Google Scholar] [PubMed]
- Bickerton, A.S.; Roberts, R.; Fielding, B.A.; Tornqvist, H.; Blaak, E.E.; Wagenmakers, A.J.; Gilbert, M.; Humphreys, S.M.; Karpe, F.; Frayn, K.N. Adipose tissue fatty acid metabolism in insulin-resistant men. Diabetologia 2008, 51, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Jocken, J.W.; Goossens, G.H.; van Hees, A.M.; Frayn, K.N.; van Baak, M.; Stegen, J.; Pakbiers, M.T.; Saris, W.H.; Blaak, E.E. Effect of beta-adrenergic stimulation on whole-body and abdominal subcutaneous adipose tissue lipolysis in lean and obese men. Diabetologia 2008, 51, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Riemens, S.C.; Sluiter, W.J.; Dullaart, R.P. Enhanced escape of non-esterified fatty acids from tissue uptake: Its role in impaired insulin-induced lowering of total rate of appearance in obesity and type ii diabetes mellitus. Diabetologia 2000, 43, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Bickerton, A.S.; McQuaid, S.E.; Roberts, R.; Karpe, F.; Frayn, K.N.; Fielding, B.A. The contribution of splanchnic fat to vldl triglyceride is greater in insulin-resistant than insulin-sensitive men and women: Studies in the postprandial state. Diabetes 2007, 56, 2433–2441. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Ghani, M.A.; Jenkinson, C.P.; Richardson, D.K.; Tripathy, D.; DeFronzo, R.A. Insulin secretion and action in subjects with impaired fasting glucose and impaired glucose tolerance: Results from the veterans administration genetic epidemiology study. Diabetes 2006, 55, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Bock, G.; Chittilapilly, E.; Basu, R.; Toffolo, G.; Cobelli, C.; Chandramouli, V.; Landau, B.R.; Rizza, R.A. Contribution of hepatic and extrahepatic insulin resistance to the pathogenesis of impaired fasting glucose: Role of increased rates of gluconeogenesis. Diabetes 2007, 56, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Pimenta, W.; Woerle, H.J.; Van Haeften, T.; Szoke, E.; Mitrakou, A.; Gerich, J. Different mechanisms for impaired fasting glucose and impaired postprandial glucose tolerance in humans. Diabetes Care 2006, 29, 1909–1914. [Google Scholar] [CrossRef] [PubMed]
- Bickerton, A.S.; Roberts, R.; Fielding, B.A.; Hodson, L.; Blaak, E.E.; Wagenmakers, A.J.; Gilbert, M.; Karpe, F.; Frayn, K.N. Preferential uptake of dietary fatty acids in adipose tissue and muscle in the postprandial period. Diabetes 2007, 56, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Van der Zijl, N.J.; Moors, C.C.; Goossens, G.H.; Hermans, M.M.; Blaak, E.E.; Diamant, M. Valsartan improves β-cell function and insulin sensitivity in subjects with impaired glucose metabolism: A randomized controlled trial. Diabetes Care 2011, 34, 845–851. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Tobin, J.D.; Andres, R. Glucose clamp technique: A method for quantifying insulin secretion and resistance. Am. J. Physiol. 1979, 237, E214–E223. [Google Scholar] [PubMed]
- Moors, C.C.; van der Zijl, N.J.; Diamant, M.; Blaak, E.E.; Goossens, G.H. Impaired insulin sensitivity is accompanied by disturbances in skeletal muscle fatty acid handling in subjects with impaired glucose metabolism. Int. J. Obes. 2012, 36, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R. Radioactive and Stable Isotope Tracers in Biomedicine: Principles and Practice of Kinetic Analysis; Wiley Liss: New York, NY, USA, 1992. [Google Scholar]
- Van Hees, A.M.; Saris, W.H.; Hul, G.B.; Schaper, N.C.; Timmerman, B.E.; Lovegrove, J.A.; Roche, H.M.; Blaak, E.E. Effects of dietary fat modification on skeletal muscle fatty acid handling in the metabolic syndrome. Int. J. Obes. 2010, 34, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.B.; Karpe, F.; Milne, R.W.; Burdge, G.C.; Wootton, S.A.; Frayn, K.N. Selective partitioning of dietary fatty acids into the vldl tg pool in the early postprandial period. J. Lipid Res. 2003, 44, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Perreault, L.; Bergman, B.C.; Hunerdosse, D.M.; Playdon, M.C.; Eckel, R.H. Inflexibility in intramuscular triglyceride fractional synthesis distinguishes prediabetes from obesity in humans. Obesity 2010, 18, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Sparks, L.M.; Bosma, M.; Brouwers, B.; van de Weijer, T.; Bilet, L.; Schaart, G.; Moonen-Kornips, E.; Eichmann, T.O.; Lass, A.; Hesselink, M.K.; et al. Reduced incorporation of fatty acids into triacylglycerol in myotubes from obese individuals with type 2 diabetes. Diabetes 2014, 63, 1583–1593. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J. Storing up trouble: Does accumulation of intramyocellular triglyceride protect skeletal muscle from insulin resistance? Clin. Exp. Pharmacol. Physiol. 2009, 36, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Popeijus, H.E.; Saris, W.H.; Mensink, R.P. Role of stearoyl-coa desaturases in obesity and the metabolic syndrome. Int. J. Obes. 2008, 32, 1076–1082. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Description | Primer Sequence | Accession Number |
|---|---|---|---|
| DGAT1 | Diacylglycerol O-Acyltransferase 1 | fw-TATTGCGGCCAATGTCTTTGC re-CACTGGAGTGATAGACTCAACCA | NM_012079.5 |
| DGAT2 | Diacylglycerol O-Acyltransferase 2 | fw-GAATGGGAGTGGCAATGCTAT re-CCTCGAAGATCACCTGCTTGT | NM_001253891.1 NM_032564.4 |
| GPAT GPAM | Mitochondrial glycerol-3-phosphate acyltransferase | fw-GATGTAAGCACACAAGTGAGGA re-TCCGACTCATTAGGCTTTCTTTC | NM_001244949.1/ NM_020918.5 |
| SPTLC1 | Serine palmitoyltransferase, long chain base subunit 1 | fw-GGTGGAGATGGTACAGGCG re-TGGTTGCCACTCTTCAATCAG | NM_001281303.1 NM_178324.2 NM_006415.3 |
| SPTLC2 | Serine palmitoyltransferase, long chain base subunit 2 | fw-TGCTCACGTATGTGGGGTATG re-GATTGGCCGATTCCAGTTGTC | NM_004863.3 |
| ASAH1 | Acid ceramidase I | fw-ATTGGCCCCAGCCTACTTTAT re-CCCTGCTTAGCATCGAGTTCAT | NM_001127505.1 NM_177924.3 NM_004315.4 |
| ASAH2 | Non-lysosomal ceramidase 2 | fw-GTGCCTTTAACCCCAGAGTCT re-GTGCAGTCAGCTCGTCCAA | NM_001143974.1 NM_019893.2 |
| CERK | Ceramide kinase | fw-TGTGCCTGTATCTGAGATCATCG re-TGCCGTCGTGCTCTCTTTAC | NM_022766.5 |
| CPT1B | Carnitine palmitoyltransferase 1B (muscle) | fw-CGGGACAGGGGTAAGTTCTG re-TCTCGCAGGTCTGCTTTTGTG | NM_152246.2 NM_004377.3 NM_001145137.1 NM_001145135.1 NM_001145134.1 NM_152245.2 |
| PPARGC1A | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha | fw-TCTGAGTCTGTATGGAGTGACAT re-CCAAGTCGTTCACATCTAGTTCA | NM_013261.3 |
| PPARA | Peroxisome proliferator-activated receptor alpha | fw-AAAAGCCTAAGGAAACCGTTCTG re- TATCGTCCGGGTGGTTGCT | NM_001001928.2 NM_005036.4 |
| PPARD | Peroxisome proliferator-activated receptor delta | fw-AGAAGAACCGCAACAAGTGC re-CTCCCCTCGTTTGCAGTCAG | NM_001171818.1 NM_006238.4 NM_177435.2 NM_001171819.1 |
| SDHB | Succinate dehydrogenase complex, subunit B | fw-CCACAGCTCCCCGTATCAAG re-TCGGAAGGTCAAAGTAGAGTCAA | NM_003000.2 |
| NDFU5B | NADH dehydrogenase (ubiquinone) 1 beta subcomplex, 5 | fw-GCTGCTCCTGTTCGACACA re-CTGCTAGTTCAGCTTGACCAAT | NM_001199958.1 NM_002492.3 |
| IFG | IGT | p-Value | |
|---|---|---|---|
| n | 12 | 14 | |
| Gender (male/female) | 7/5 | 7/7 | |
| Family history type 2 diabetes | 10 | 6 | |
| Age (years) | 59.3 ± 1.7 | 58.6 ± 1.9 | 0.863 |
| Weight (kg) | 83.7 ± 4.1 | 90.3 ± 3.4 | 0.112 |
| Height (cm) | 170.6 ± 2.0 | 170.2 ± 2.4 | 0.816 |
| BMI (kg/m2) | 28.7 ± 1.1 | 31.2 ± 1.1 | 0.126 |
| Waist/Hip ratio | 0.98 ± 0.02 | 1.00 ± 0.02 | 0.329 |
| Systolic BP (mmHg) | 129 ± 3 | 135.1 ± 3 | 0.447 |
| Diastolic BP (mmHg) | 80 ± 2 | 83 ± 2 | 0.179 |
| Fasting glucose (mmol/L) | 5.9 ± 0.3 | 5.9 ± 0.2 | 0.786 |
| 2 h glucose (mmol/L) | 5.3 ± 0.4 | 8.8 ± 0.8 | <0.001 |
| HbA1C | 6.1 ± 0.1 | 6.0 ± 0.2 | 0.713 |
| M-value (mg·kg−1·min−1) | 4.1 ± 0.5 | 2.9 ± 0.3 | 0.064 |
| Fasting insulin (mU/L) | 12.2 ± 1.9 | 16.9 ± 2.1 | 0.093 |
| Total body fat (%) | 29.6 ± 2.0 | 33.6 ± 2.0 | 0.122 |
| HDL cholesterol (mmol/L) | 1.1 ± 0.1 | 1.2 ± 0.1 | 0.433 |
| LDL cholesterol (mmol/L) | 3.7 ± 0.2 | 3.6 ± 0.2 | 0.667 |
| Triacylglycerol (mmol/L) | 1.0 ± 0.1 | 1.4 ± 0.1 | 0.048 |
| IFG (n = 12) | IGT (n = 14) | p-Value | |
|---|---|---|---|
| Forearm blood flow (mL·100mL−1·min−1) | |||
| Fasting | 2.35 ± 0.19 | 2.59 ± 0.18 | 0.257 |
| Postprandial (AUC0–4 h) | 2.58 ± 0.22 | 2.85 ± 0.26 | 0.298 |
| Net fluxes across forearm muscle | |||
| Glycerol (nmol·100 mL−1·min−1) | |||
| Fasting | −18.7 ± 7.8 | −22.1 ± 19.0 | 0.978 |
| Postprandial (AUC0–4 h) | −17.1 ± 7.3 | −26.5 ± 15.4 | 0.753 |
| FFA (nmol·100 mL−1·min−1) | |||
| Fasting | 50.3 ± 19.8 | 40.8 ± 37.6 | 0.851 |
| Postprandial (AUC0–4 h) | 37.1 ± 15.2 | 11.3 ± 19.8 | 0.379 |
| [2H2]-palmitate FFA (nmol·100 mL−1·min−1) | |||
| Fasting | 1.59 ± 0.18 | 1.61 ± 0.19 | 0.917 |
| Postprandial (AUC0–4 h) | 2.45 ± 0.12 | 2.26 ± 0.14 | 0.335 |
| TAG (nmol·100 mL−1·min−1) | |||
| Fasting | 30.3 ± 6.8 | 42.6 ± 18.9 | 0.656 |
| Postprandial (AUC0–4 h) | 104.7 ± 18.9 | 82.5 ± 15.9 | 0.204 |
| [2H2]-palmitate TAG (nmol·100 mL−1·min−1) | |||
| Fasting | −0.20 ± 0.17 | 0.01 ± 0.20 | 0.374 |
| Postprandial (AUC0–4 h) | −0.57 ± 0.47 | 0.70 ± 0.24 | 0.043 |
| [U-13C]-palmitate TAG (nmol·100 mL−1·min−1) | |||
| Fasting | N/A | N/A | - |
| Postprandial (AUC0.5–4 h) | 0.01 ± 0.32 | 0.77 ± 0.23 | 0.109 |
| IFG (n = 12) | IGT (n = 14) | p-Value | |
|---|---|---|---|
| TAG | |||
| Total (mol/g dry weight) | 178.1 ± 26.5 | 384.5 ± 91.8 | 0.025 |
| Saturation (%) | 35.6 ± 1.2 | 31.3 ± 1.1 | 0.017 |
| MUFA (%) | 47.0 ± 1.6 | 49.5 ± 1.7 | 0.190 |
| PUFA (%) | 17.4 ± 1.5 | 19.1 ± 1.9 | 0.562 |
| FSR (%/h) | 0.26 ± 0.07 | 0.13 ± 0.03 | 0.073 |
| DAG | |||
| Total (mol/g dry weight) | 6.0 ± 1.0 | 3.0 ± 0.3 | 0.031 |
| Saturation (%) | 42.3 ± 1.9 | 36.4 ± 0.9 | 0.059 |
| MUFA (%) | 40.8 ± 1.4 | 48.7 ± 1.2 | ≤0.001 |
| PUFA (%) | 16.9 ± 1.2 | 14.9 ± 0.6 | 0.124 |
| FSR (%/h) | 0.39 ± 0.12 | 0.28 ± 0.06 | 0.439 |
| PL | |||
| Total (mol/g dry weight) | 71.4 ± 3.4 | 59.2 ± 3.7 | 0.029 |
| Saturation (%) | 40.6 ± 1.3 | 39.7 ± 0.7 | 0.610 |
| MUFA (%) | 10.4 ± 0.4 | 11.4 ± 1.0 | 0.333 |
| PUFA (%) | 49.0 ± 1.3 | 48.9 ± 0.9 | 0.911 |
| FFA | |||
| Total (mol/g dry weight) | 3.2 ± 1.0 | 2.2 ± 0.6 | 0.311 |
| Saturation (%) | 50.1 ± 3.5 | 68.0 ± 4.0 | 0.004 |
| MUFA (%) | 34.0 ± 2.9 | 19.7 ± 2.9 | 0.003 |
| PUFA (%) | 16.0 ± 2.6 | 12.3 ± 2.0 | 0.352 |
| Gene | IFG | IGT | p-Value |
|---|---|---|---|
| TAG synthesis | |||
| DGAT1 | 1.79 ± 0.56 | 1.99 ± 1.03 | 0.203 |
| DGAT2 | 0.25 ± 0.16 | 0.30 ± 0.25 | 0.195 |
| GPAT1 | 1.79 ± 0.56 | 2.00 ± 1.03 | 0.477 |
| Ceramide synthesis | |||
| SPTLC1 | 1.36 ± 0.43 | 1.30 ± 0.47 | 0.181 |
| SPTLC2 | 1.48 ± 0.38 | 1.42 ± 0.51 | 0.117 |
| ASAH2 | 0.31 ± 0.18 | 0.50 ± 0.69 | 0.848 |
| CERK | 1.71 ± 0.88 | 1.31 ± 0.77 | 0.072 |
| Oxidative metabolism | |||
| mCPT1 | 2.01 ± 0.18 | 1.81 ± 0.18 | 0.434 |
| PGC1α | 0.63 ± 0.07 | 0.45 ± 0.04 | 0.027 |
| PPARα | 2.44 ± 0.22 | 1.70 ± 0.15 | 0.009 |
| PPARδ | 0.48 ± 0.03 | 0.43 ± 0.04 | 0.309 |
| SDHB | 2.00 ± 0.17 | 1.57 ± 0.13 | 0.063 |
| NDFU5B | 1.83 ± 0.14 | 1.66 ± 0.12 | 0.384 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goossens, G.H.; Moors, C.C.M.; Jocken, J.W.E.; Van der Zijl, N.J.; Jans, A.; Konings, E.; Diamant, M.; Blaak, E.E. Altered Skeletal Muscle Fatty Acid Handling in Subjects with Impaired Glucose Tolerance as Compared to Impaired Fasting Glucose. Nutrients 2016, 8, 164. https://doi.org/10.3390/nu8030164
Goossens GH, Moors CCM, Jocken JWE, Van der Zijl NJ, Jans A, Konings E, Diamant M, Blaak EE. Altered Skeletal Muscle Fatty Acid Handling in Subjects with Impaired Glucose Tolerance as Compared to Impaired Fasting Glucose. Nutrients. 2016; 8(3):164. https://doi.org/10.3390/nu8030164
Chicago/Turabian StyleGoossens, Gijs H., Chantalle C. M. Moors, Johan W. E. Jocken, Nynke J. Van der Zijl, Anneke Jans, Ellen Konings, Michaela Diamant, and Ellen E. Blaak. 2016. "Altered Skeletal Muscle Fatty Acid Handling in Subjects with Impaired Glucose Tolerance as Compared to Impaired Fasting Glucose" Nutrients 8, no. 3: 164. https://doi.org/10.3390/nu8030164