
The Inhibitory Effect of Early Pregnancy Factor on Red Meat Neu5Gc-Mediated Antibody Production in CMAH−/− Mice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Grouping
2.3. Validation of the In Vitro Activity of EPF Protein via Rosette Inhibition Assay
2.4. Detection of Serum Neu5Gc Antibody Levels Using the Indirect ELISA Method
2.5. Quantification of Inflammatory Cytokines Using an ELISA Kit
2.6. Determination of Myeloperoxidase (MPO) Activity
2.7. Determination of Tissue and Organ Index
2.8. HE Staining Analysis of Pathological Changes in Chronic Inflammation
2.9. Immunohistochemical Analysis of CD4 Cell Proliferation and Inhibition
2.10. Determination of CD Molecules Associated with B Cell Activation by RT-PCR
2.11. Statistical Analysis
3. Result
3.1. Cell Level Analysis of EPF Protein Activity
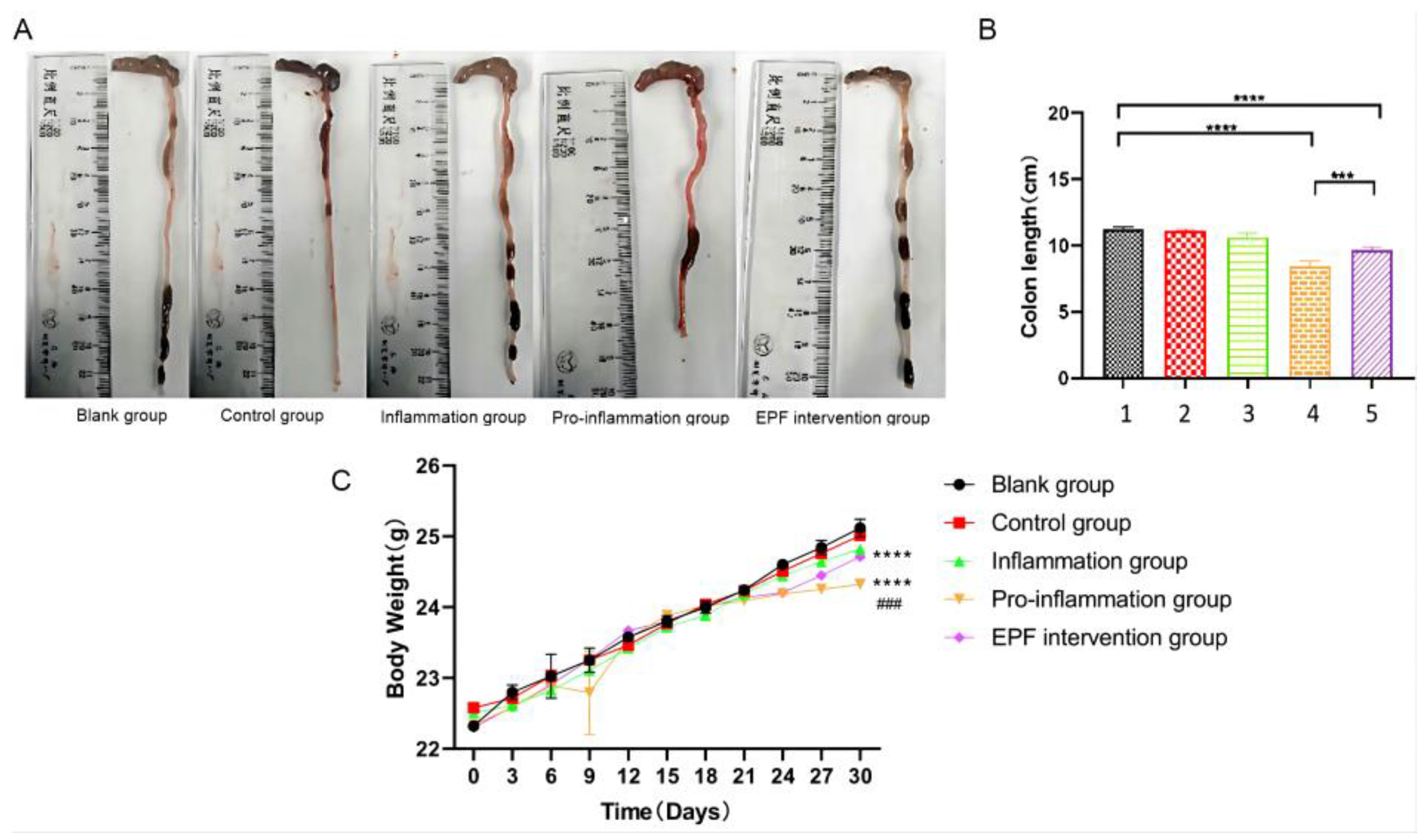
3.2. The Effect of EPF on the Colon Length and Body Weight in Mice with Red Meat Neu5Gc-Induced Inflammation
3.3. EPF Reduced the Levels of Anti-Neu5Gc Antibodies in Red Meat Neu5GC-Induced Inflammation Mice
3.4. EPF Attenuated the Content of Inflammatory Factors in the Serum of Inflammatory Mice
3.5. EPF Alleviates Histopathological Damage in Mice with Neu5Gc-Induced Inflammation
3.6. After Undergoing EPF Treatment, the CD4 Cell Count in Neu5Gc-Induced Inflammatory Mice Exhibited a Significant Decrease
3.7. EPF Downregulated the Expression of Key Molecules of B Cell Activation in Neu5GC-Induced Inflammatory Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, H.C.; Yang, X.; Zhang, C.; Zhu, C.; Tao, G.Z.; Zhao, L.J.; Tang, S.W.; Shu, Z.; Cai, J.; Dai, S.B.; et al. Red and Processed Meat Intake Is Associated with Higher Gastric Cancer Risk: A Meta-Analysis of Epidemiological Observational Studies. PLoS ONE 2013, 8, e70955. [Google Scholar] [CrossRef]
- Varki, A. N-glycolylneuraminic acid deficiency in humans. Biochimie 2001, 83, 615–622. [Google Scholar] [CrossRef]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef]
- Alisson-Silva, F.; Kawanishi, K.; Varki, A. Human risk of diseases associated with red meat intake: Analysis of current theories and proposed role for metabolic incorporation of a non-human sialic acid. Mol. Aspects Med. 2016, 51, 16–30. [Google Scholar] [CrossRef]
- Malykh, Y.N.; Schauer, R.; Shaw, L. N-glycolylneuraminic acid in human tumours. Biochimie 2001, 83, 623–634. [Google Scholar] [CrossRef]
- Hedlund, M.; Padler-Karavani, V.; Varki, N.M.; Varki, A. Evidence for a human-specific mechanism for diet and antibody-mediated inflammation in carcinoma progression. Proc. Natl. Acad. Sci. USA 2008, 105, 18936–18941. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.P.; Valdés, Z.; Casacó, A.; Pimentel, G.; González, J.; Alvarez, I.; Osorio, M.; Velazco, M.; Figueroa, M.; Ortiz, R.; et al. Clinical evidences of GM3 (NeuGc) ganglioside expression in human breast cancer using the 14F7 monoclonal antibody labelled with 99mTc. Breast Cancer Res. Treat. 2006, 96, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Labrada, M.; Dorvignit, D.; Hevia, G.; Rodríguez-Zhurbenko, N.; Hernández, A.M.; Vázquez, A.M.; Fernández, L.E. GM3(Neu5Gc) ganglioside: An evolution fixed neoantigen for cancer immunotherapy. Semin. Oncol. 2018, 45, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Torbidoni, A.V.; Scursoni, A.; Camarero, S.; Segatori, V.; Gabri, M.; Alonso, D.; Chantada, G.; de Dávila, M.T.G. Immunoreactivity of the 14F7 Mab raised against N-Glycolyl GM3 Ganglioside in retinoblastoma tumours. Acta Ophthalmol. 2015, 93, E294–E300. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Wu, Y.; Li, C.; Tang, J.; Wang, X.; Ren, G.; Carr, A.; Pérez, R.; Guo, W. N-Glycolyl GM3 ganglioside immunoexpression in oral mucosal melanomas of Chinese. Oral Dis. 2012, 18, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Morton, H.H.V.; Clunie, G.J. Immunosuppression detected in pregnant mice by rosette inhibition test. Nature 1974, 249, 459–460. [Google Scholar] [CrossRef] [PubMed]
- Morton, H.; Rolfe, B.; Clunie, G.J. An early pregnancy factor detected in human serum by the rosette inhibition test. Lancet 1977, 1, 394–397. [Google Scholar] [CrossRef]
- Morton, H.; Mckay, D.; Murphy, R.M.; Somodevilla-Torres, M.J.; Swanson, C.E.; Cassady, A.I.; Summers, K.M.; Cavanagh, A.C. Production of a recombinant form of early pregnancy factor that can prolong allogeneic skin graft survival time in rats. Immunol. Cell Biol. 2000, 78, 603–607. [Google Scholar] [CrossRef]
- Cruz, Y.P.; Selwood, L.; Morton, H.; Cavanagh, A.C. Significance of serum early pregnancy factor concentrations during pregnancy and embryonic development in Sminthopsis macroura (Spencer) (Marsupialia: Dasyuridae). Reproduction 2001, 121, 933–939. [Google Scholar] [CrossRef]
- Taylor, R.E.; Gregg, C.J.; Padler-Karavani, V.; Ghaderi, D.; Yu, H.; Huang, S.S.; Sorensen, R.U.; Chen, X.; Inostroza, J.; Nizet, V.; et al. Novel mechanism for the generation of human xeno-autoantibodies against the nonhuman sialic acid N-glycolylneuraminic acid. J. Exp. Med. 2010, 207, 1637–1646. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.T.; Golden, B.; Okerblom, J.; Camboni, M.; Chandrasekharan, K.; Xu, R.; Varki, A.; Flanigan, K.M.; Kornegay, J.N. A Comparative Study of N-glycolylneuraminic Acid (Neu5Gc) and Cytotoxic T Cell (CT) Carbohydrate Expression in Normal and Dystrophin-Deficient Dog and Human Skeletal Muscle. PLoS ONE 2014, 9, e88226. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H. Immune cells contribute to systemic cross-talk between the embryo and mother during early pregnancy in cooperation with the endocrine system. Reprod. Med. Biol. 2006, 5, 19–29. [Google Scholar] [CrossRef]
- Samraj, A.N.; Pearce, O.M.T.; Läubli, H.; Crittenden, A.N.; Bergfeld, A.K.; Banda, K.; Gregg, C.J.; Bingman, A.E.; Secrest, P.; Diaz, S.L.; et al. A red meat-derived glycan promotes inflammation and cancer progression. Proc. Natl. Acad. Sci. USA 2015, 112, 542–547. [Google Scholar] [CrossRef]
- Tahara, H.; Ide, K.; Basnet, N.B.; Tanaka, Y.; Matsuda, H.; Takematsu, H.; Kozutsumi, Y.; Ohdan, H. Immunological Property of Antibodies against N-Glycolylneuraminic Acid Epitopes in Cytidine Monophospho–N-Acetylneuraminic Acid Hydroxylase-Deficient Mice. J. Immunol. 2010, 184, 3269–3275. [Google Scholar] [CrossRef]
- Zheng, Z.Q.; Qin, Z.H.; Ma, A.Y.; Qiao, C.X.; Wang, H. Detection of early pregnancy factor-like activity in human amniotic fluid. Am. J. Reprod. Immunol. 1990, 22, 9–11. [Google Scholar]
- Athanasas-Platsis, S.; Morton, H.; Dunglison, G.F.; Kaye, P.L. Antibodies to early pregnancy factor retard embryonic development in mice in vivo. J. Reprod. Fertil. 1991, 92, 443–451. [Google Scholar] [CrossRef]
- Cheng, S.J.; Zheng, Z.Q. Early pregnancy factor in cervical mucus of pregnant women. Am. J. Reprod. Immunol. 2004, 51, 102–105. [Google Scholar] [CrossRef]
- Lai, J.L.; Liu, Y.H.; Peng, Y.C.; Ge, P.; He, C.F.; Liu, C.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Treatment of Lipopolysaccharide-Induced Mastitis in a Mouse Model and Activity in Mouse Mammary Epithelial Cells. Mediat. Inflamm. 2017, 2017, 3082805. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.H.; Hayakawa, T.; Diaz, S.; Krings, M.; Indriati, E.; Leakey, M.; Paabo, S.; Satta, Y.; Takahata, N.; Varki, A. Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 11736–11741. [Google Scholar] [CrossRef] [PubMed]
- Samraj, A.N.; Läubli, H.; Varki, N.; Varki, A. Involvement of a non-human sialic Acid in human cancer. Front. Oncol. 2014, 4, 33. [Google Scholar] [CrossRef] [PubMed]
- Bergfeld, A.K.; Pearce, O.M.T.; Diaz, S.L.; Pham, T.; Varki, A. Metabolism of Vertebrate Amino Sugars with N-Glycolyl Groups: Elucidating the intracellular fate of the non-human sialic acid N-glycolylneuraminic acid. J. Biol. Chem. 2012, 287, 28865–28881. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xie, B.Y.; Wang, B.; Troy, F.A. LC-MS/MS glycomic analyses of free and conjugated forms of the sialic acids, Neu5Ac, Neu5Gc and KDN in human throat cancers. Glycobiology 2015, 25, 1362–1374. [Google Scholar] [CrossRef]
- Lang, T.J.; Nguyen, P.; Peach, R.; Gause, W.C.; Via, C.S. In vivo CD86 blockade inhibits CD4+ T cell activation, whereas CD80 blockade potentiates CD8+ T cell activation and CTL effector function. J. Immunol. 2002, 168, 3786–3792. [Google Scholar] [CrossRef] [PubMed]
- Vasu, C.; Wang, A.; Gorla, S.R.; Kaithamana, S.; Prabhakar, B.S.; Holterman, M.J. CD80 and CD86 C domains play an important role in receptor binding and co-stimulatory properties. Int. Immunol. 2003, 15, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Suvas, S.; Singh, V.; Sahdev, S.; Vohra, H.; Agrewala, J.N. Distinct role of CD80 and CD86 in the regulation of the activation of B cell and B cell lymphoma. J. Biol. Chem. 2002, 277, 7766–7775. [Google Scholar] [CrossRef]
- Mann, J.R.; Backlund, M.G.; DuBois, R.N. Mechanisms of disease: Inflammatory mediators and cancer prevention. Nat. Clin. Pract. Oncol. 2005, 2, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.M.; Fowkes, F.G.R.; Belch, J.F.F.; Ogawa, H.; Warlow, C.P.; Meade, T.W. Effect of Daily Aspirin on Long-Term Risk of Death Due to Cancer: Analysis of Individual Patient Data From Randomized Trials. Obstet. Gynecol. Surv. 2011, 66, 222–223. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer’s Name | Sequences (5′–3′) |
|---|---|
| CD19-F | AGACTGTGTTGTAACCCATCCC |
| CD19-R | AGGCAAGGGGGGGGTGATGTAAAC |
| CD80-F | ACCCACAACATAACTGAGTCT |
| CD80-R | TTCCAACCAAGAGAAGCGGAGG |
| CD86-F | TAAGCAAGGTCACCGAAA |
| CD86-R | AGAACACACACACGGTCATATG |
| CD138-F | ACCAGCAGACACCGAGAC |
| CD138-R | TGGGAGCCGAGTCTCATGG |
| β-action F | GGCTGTATTCCCCTCCATCG |
| β-actin-R | CCAGTTGGTAACAATGCCATGT |
| Group | Number | Lymphocyte Count | Rosette Forming Rate | Average Value | ||
|---|---|---|---|---|---|---|
| Control | 3 | 200 | 34% | 40% | 38% | 37.3% |
| EPF | 3 | 200 | 20% | 17% | 21% | 19.3% |
| EPF + Antibody | 3 | 200 | 30% | 31% | 34% | 31.7% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Ren, H.; Wang, H.; Li, H.; Guo, J.; Xiao, Y.; Guo, Y.; Liu, M.; Duan, F.; Hu, P.; et al. The Inhibitory Effect of Early Pregnancy Factor on Red Meat Neu5Gc-Mediated Antibody Production in CMAH−/− Mice. Nutrients 2024, 16, 905. https://doi.org/10.3390/nu16060905
Wang C, Ren H, Wang H, Li H, Guo J, Xiao Y, Guo Y, Liu M, Duan F, Hu P, et al. The Inhibitory Effect of Early Pregnancy Factor on Red Meat Neu5Gc-Mediated Antibody Production in CMAH−/− Mice. Nutrients. 2024; 16(6):905. https://doi.org/10.3390/nu16060905
Chicago/Turabian StyleWang, Cong, Honglin Ren, Han Wang, Haosong Li, Jian Guo, Yiran Xiao, Yuxi Guo, Mengdi Liu, Fuchun Duan, Pan Hu, and et al. 2024. "The Inhibitory Effect of Early Pregnancy Factor on Red Meat Neu5Gc-Mediated Antibody Production in CMAH−/− Mice" Nutrients 16, no. 6: 905. https://doi.org/10.3390/nu16060905