
The Impact of Diet on miRNA Regulation and Its Implications for Health: A Systematic Review


Abstract
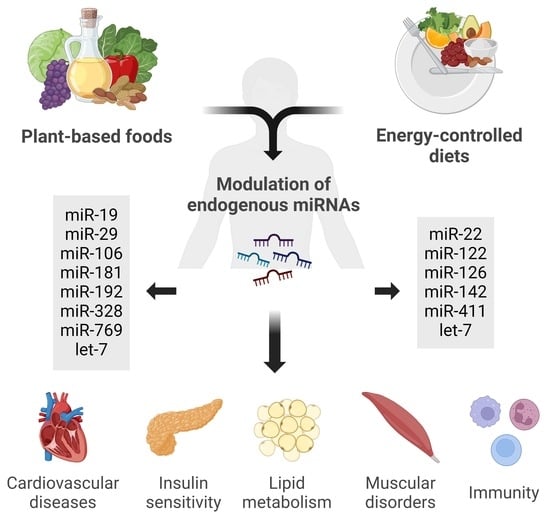
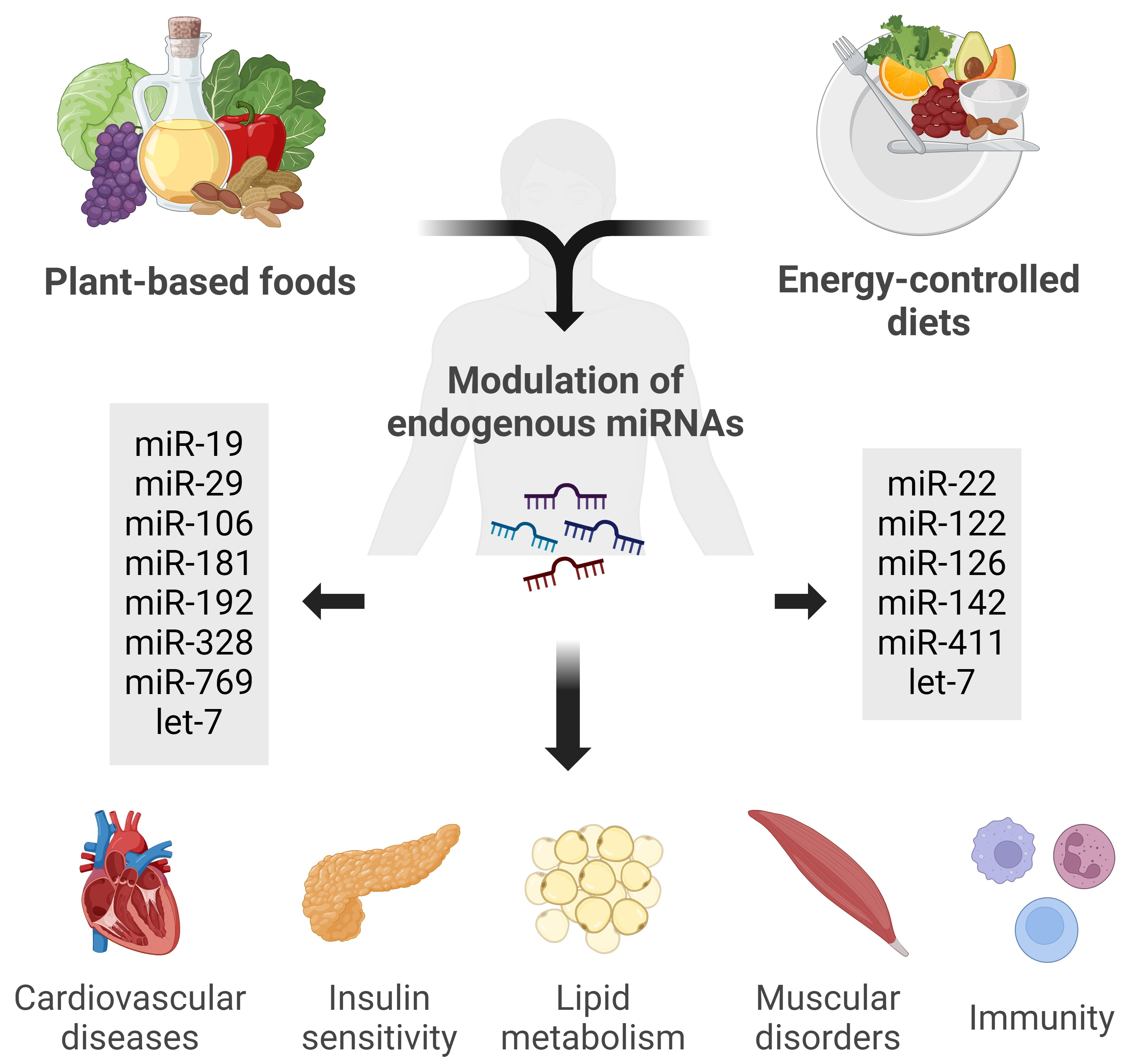
:
1. Introduction
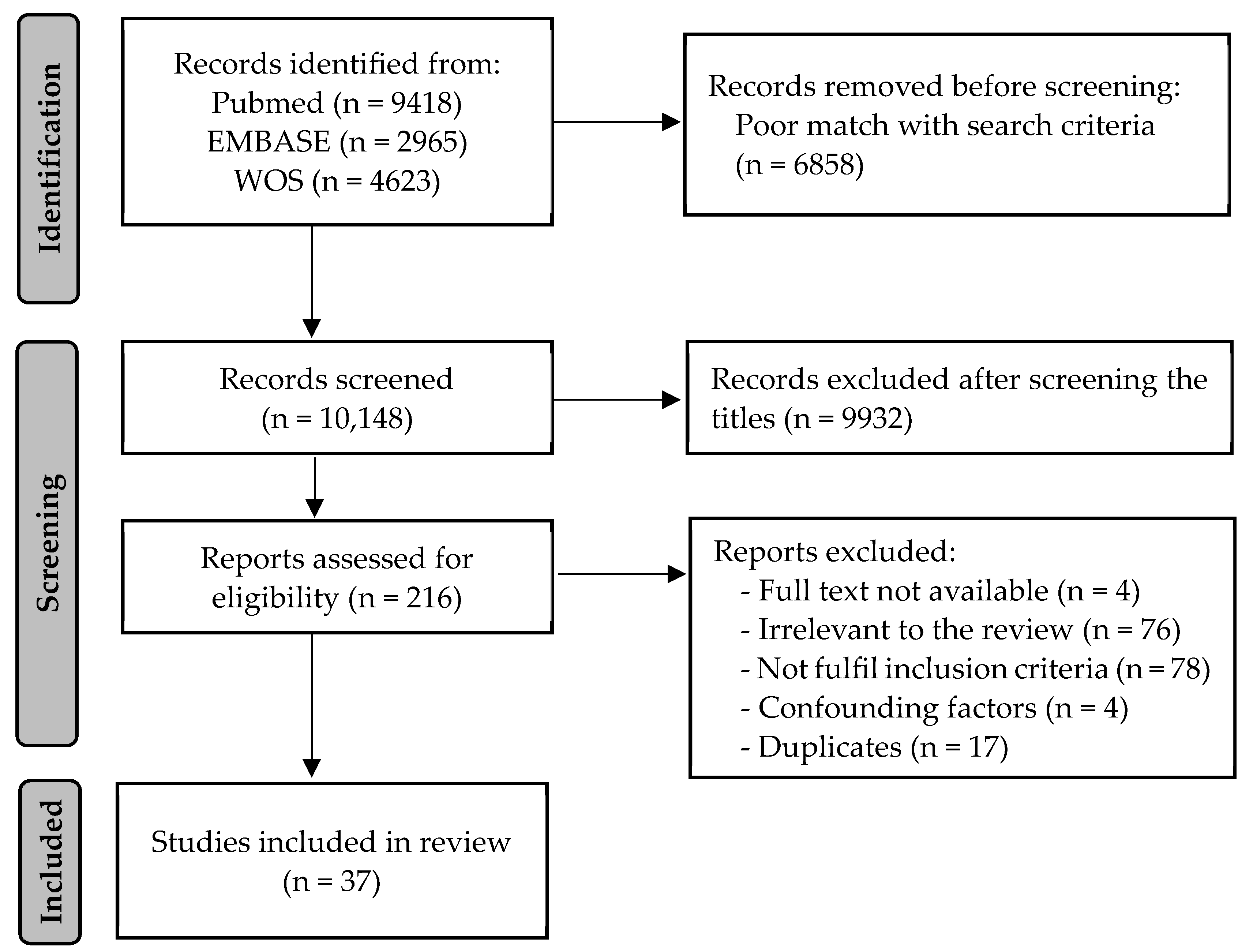
2. Materials and Methods
3. Results
3.1. Characterization of the Included Studies
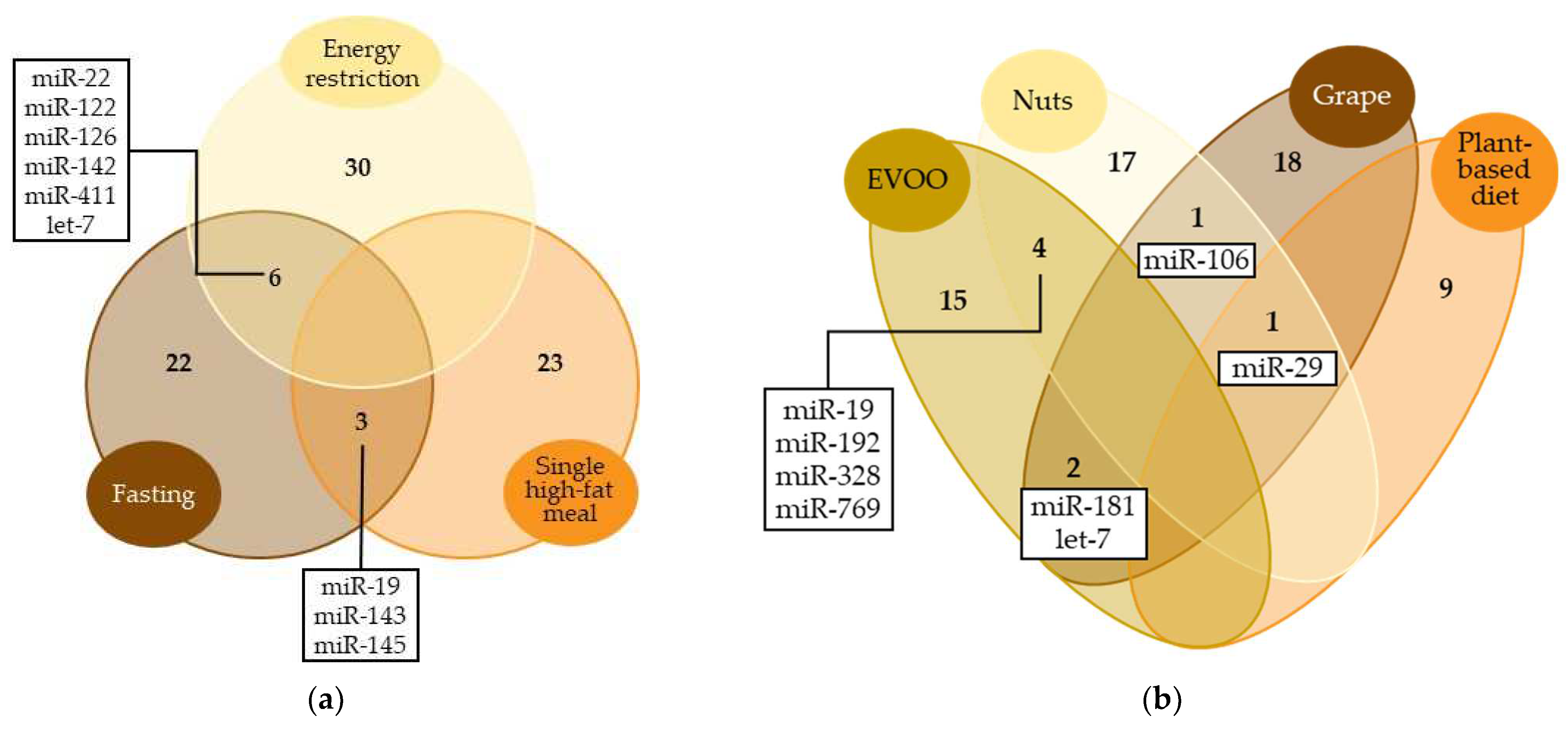
3.2. Modulation of miRNA Expression by Energy-Controlled Diets
3.3. Modulation of miRNA Expression by Fat Intake
3.4. Dietary Patterns Related to miRNA Modulation in Observational Studies
3.5. Other Dietary Patterns Related to miRNA Modulation
4. Discussion
4.1. Influence of Energy Intake on miRNA Regulation
4.2. Influence of a Mediterranean Diet and Plant-Based Foods on miRNA Regulation
4.3. Biological Effects of Diet-Modulated miRNAs
4.4. Exogenous miRNAs from Dietary Sources
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Noncommunicable Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases (accessed on 10 June 2023).
- Hunter, D.J.; Reddy, K.S. Noncommunicable Diseases. N. Engl. J. Med. 2013, 369, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT-Lancet Commission on Healthy Diets from Sustainable Food Systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, E.A.; McKay, S. The Role of Specific Components of a Plant-Based Diet in Management of Dyslipidemia and the Impact on Cardiovascular Risk. Nutrients 2020, 12, 2671. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, L.J.; Di Bella, G.; Veronese, N.; Barbagallo, M. Impact of Mediterranean Diet on Chronic Non-Communicable Diseases and Longevity. Nutrients 2021, 13, 2028. [Google Scholar] [CrossRef] [PubMed]
- Bruins, M.J.; Van Dael, P.; Eggersdorfer, M. The Role of Nutrients in Reducing the Risk for Noncommunicable Diseases during Aging. Nutrients 2019, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean Bioactive Peptides and Their Functional Properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhou, B.; Ross, S.A.; Zempleni, J. Nutrition, MicroRNAs, and Human Health. Adv. Nutr. 2017, 8, 105–112. [Google Scholar] [CrossRef]
- Milenkovic, D.; Jude, B.; Morand, C. MiRNA as Molecular Target of Polyphenols Underlying Their Biological Effects. Free Radic. Biol. Med. 2013, 64, 40–51. [Google Scholar] [CrossRef]
- Kura, B.; Parikh, M.; Slezak, J.; Pierce, G.N. The Influence of Diet on MicroRNAs That Impact Cardiovascular Disease. Molecules 2019, 24, 1509. [Google Scholar] [CrossRef]
- Otsuka, K.; Yamamoto, Y.; Matsuoka, R.; Ochiya, T. Maintaining Good MiRNAs in the Body Keeps the Doctor Away?: Perspectives on the Relationship between Food-Derived Natural Products and MicroRNAs in Relation to Exosomes/Extracellular Vesicles. Mol. Nutr. Food Res. 2018, 62, 1700080. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous Plant MIR168a Specifically Targets Mammalian LDLRAP1: Evidence of Cross-Kingdom Regulation by MicroRNA. Cell Res. 2012, 22, 107. [Google Scholar] [CrossRef]
- Lukasik, A.; Zielenkiewicz, P. In Silico Identification of Plant MiRNAs in Mammalian Breast Milk Exosomes—A Small Step Forward? PLoS ONE 2014, 9, 99963. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; He, F.; Ma, L.; Cao, M.; Zhou, Z.; Wei, Z.; Xue, Y.; Sang, X.; Chong, H.; Tian, C.; et al. The Potential Atheroprotective Role of Plant MIR156a as a Repressor of Monocyte Recruitment on Inflamed Human Endothelial Cells. J. Nutr. Biochem. 2018, 57, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Debnath, T.; Deb Nath, N.C.; Kim, E.K.; Lee, K.G. Role of Phytochemicals in the Modulation of MiRNA Expression in Cancer. Food Funct. 2017, 8, 3432–3442. [Google Scholar] [CrossRef] [PubMed]
- Saquib, M.; Agnihotri, P.; Monu, X.; Biswas, S. Exogenous MiRNA: A Perspective Role as Therapeutic in Rheumatoid Arthritis. Curr. Rheumatol. Rep. 2021, 23, 43. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, K.J.; Conlon, M.A.; Young, G.P.; Topping, D.L.; Hu, Y.; Winter, J.M.; Bird, A.R.; Cobiac, L.; Kennedy, N.A.; Michael, M.Z.; et al. Dietary Manipulation of Oncogenic MicroRNA Expression in Human Rectal Mucosa: A Randomized Trial. Cancer Prev. Res. 2014, 7, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An Overview of MicroRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Bernardo, B.C.; Ooi, J.Y.Y.; Lin, R.C.Y.; Mcmullen, J.R. MiRNA Therapeutics: A New Class of Drugs with Potential Therapeutic Applications in the Heart. Future Med. Chem. 2015, 7, 1771–1792. [Google Scholar] [CrossRef]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A Brief Review on the Mechanisms of MiRNA Regulation. Genom. Proteom. Bioinform. 2009, 7, 147–154. [Google Scholar] [CrossRef]
- Ruiz, G.P.; Camara, H.; Fazolini, N.P.B.; Mori, M.A. Extracellular MiRNAs in Redox Signaling: Health, Disease and Potential Therapies. Free Radic. Biol. Med. 2021, 173, 170–187. [Google Scholar] [CrossRef]
- Kalayinia, S.; Arjmand, F.; Maleki, M.; Malakootian, M.; Singh, C.P. MicroRNAs: Roles in Cardiovascular Development and Disease. Cardiovasc. Pathol. 2021, 50, 107296. [Google Scholar] [CrossRef] [PubMed]
- Çakmak, H.A.; Demir, M. MicroRNA and Cardiovascular Diseases. Balk. Med. J. 2020, 37, 60–71. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Kuang, G.; Wu, Y.; Ou, C. Emerging Roles of Exosomal MiRNAs in Diabetes Mellitus. Clin. Transl. Med. 2021, 11, 11. [Google Scholar] [CrossRef]
- Vasu, S.; Kumano, K.; Darden, C.M.; Rahman, I.; Lawrence, M.C.; Naziruddin, B. MicroRNA Signatures as Future Biomarkers for Diagnosis of Diabetes States. Cells 2019, 8, 1533. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Huang, F.; Gu, X.; Zhang, M.; Wen, J.; Wang, X.; You, L.; Cui, X.; Ji, C.; Guo, X. Adipogenic MiRNA and Meta-Signature MiRNAs Involved in Human Adipocyte Differentiation and Obesity. Oncotarget 2016, 7, 40830–40845. [Google Scholar] [CrossRef]
- Zaiou, M.; El Amri, H.; Bakillah, A. The Clinical Potential of Adipogenesis and Obesity-Related MicroRNAs. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Pomar, C.A.; Castro, H.; Pico, C.; Serra, F.; Palou, A.; Sanchez, J. Cafeteria Diet Consumption during Lactation in Rats, Rather than Obesity Per Se, Alters MiR-222, MiR-200a, and MiR-26a Levels in Milk. Mol. Nutr. Food Res. 2019, 63, 1800928. [Google Scholar] [CrossRef] [PubMed]
- Guedes, E.C.; da Silva, I.B.; Lima, V.M.; Miranda, J.B.; Albuquerque, R.P.; Ferreira, J.C.B.; Barreto-Chaves, M.L.M.; Diniz, G.P. High Fat Diet Reduces the Expression of MiRNA-29b in Heart and Increases Susceptibility of Myocardium to Ischemia/Reperfusion Injury. J. Cell. Physiol. 2019, 234, 9399–9407. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, B.; Wang, X.; Liu, Z.; Li, J.; Zhang, S.; Gu, X.; Jia, M.; Guo, H.; Feng, N.; et al. P53/PANK1/MiR-107 Signalling Pathway Spans the Gap between Metabolic Reprogramming and Insulin Resistance Induced by High-Fat Diet. J. Cell. Mol. Med. 2020, 24, 3611–3624. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. MiRNet 2.0: Network-Based Visual Analytics for MiRNA Functional Analysis and Systems Biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Giardina, S.; Hernández-Alonso, P.; Díaz-López, A.; Salas-Huetos, A.; Salas-Salvadó, J.; Bulló, M. Changes in Circulating MiRNAs in Healthy Overweight and Obese Subjects: Effect of Diet Composition and Weight Loss. Clin. Nutr. 2019, 38, 438–443. [Google Scholar] [CrossRef]
- Margolis, L.M.; Rivas, D.A.; Pasiakos, S.M.; McClung, J.P.; Ceglia, L.; Fielding, R.A. Upregulation of Circulating MyomiR Following Short-Term Energy Restriction Is Inversely Associated with Whole Body Protein Synthesis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R298–R304. [Google Scholar] [CrossRef] [PubMed]
- Assmann, T.S.; Riezu-Boj, J.I.; Milagro, F.I.; Martínez, J.A. Circulating Adiposity-Related MicroRNAs as Predictors of the Response to a Low-Fat Diet in Subjects with Obesity. J. Cell. Mol. Med. 2020, 24, 2956–2967. [Google Scholar] [CrossRef] [PubMed]
- Duggan, C.; Tapsoba, J.d.D.; Scheel, J.; Wang, C.Y.; McTiernan, A. Weight Loss Reduces Circulating Micro-RNA Related to Obesity and Breast Cancer in Postmenopausal Women. Epigenetics 2022, 17, 2082–2095. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Xue, Q.; Rood, J.; Bray, G.A.; Sacks, F.M.; Qi, L. Circulating Thrifty MicroRNA Is Related to Insulin Sensitivity, Adiposity, and Energy Metabolism in Adults with Overweight and Obesity: The POUNDS Lost Trial. Am. J. Clin. Nutr. 2023, 117, 121–129. [Google Scholar] [CrossRef]
- Hess, A.L.; Larsen, L.H.; Udesen, P.B.; Sanz, Y.; Larsen, T.M.; Dalgaard, L.T. Levels of Circulating MiR-122 Are Associated with Weight Loss and Metabolic Syndrome. Obesity 2020, 28, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Jayasooriya, V.; Johnson, N.; Bradley, A.; Kotarsky, C.; Jepng’etich, L.; Friesner, D.; Stastny, S.; Hackney, K.J.; Nawarathna, D. A Miniaturized MicroRNA Sensor Identifies Targets Associated with Weight Loss in a Diet and Exercise Intervention among Healthy Overweight Individuals. Sensors 2022, 22, 6758. [Google Scholar] [CrossRef]
- Kristensen, M.M.; Davidsen, P.K.; Vigelsø, A.; Hansen, C.N.; Jensen, L.J.; Jessen, N.; Bruun, J.M.; Flemming, D.; Helge, J.W. MiRNAs in Human Subcutaneous Adipose Tissue: Effects of Weight Loss Induced by Hypocaloric Diet and Exercise. Obesity 2017, 25, 572–580. [Google Scholar] [CrossRef]
- Milagro, F.I.; Miranda, J.; Portillo, M.P.; Fernandez-Quintela, A.; Campión, J.; Martínez, J.A. High-Throughput Sequencing of MicroRNAs in Peripheral Blood Mononuclear Cells: Identification of Potential Weight Loss Biomarkers. PLoS ONE 2013, 8, e54319. [Google Scholar] [CrossRef]
- Müller, S.; Wallner, S.; Schmitz, G.; Loew, T.; Stempfl, T.; Möhle, C.; Strack, C.; Sag, S.; Baessler, A.; Fischer, M. SNP Dependent Modulation of Circulating MiRNAs from the MiR25/93/106 Cluster in Patients Undergoing Weight Loss. Gene 2020, 753, 144787. [Google Scholar] [CrossRef] [PubMed]
- Parr, E.B.; Camera, D.M.; Burke, L.M.; Phillips, S.M.; Coffey, V.G.; Hawley, J.A. Circulating MicroRNA Responses between ‘High’ and ‘Low’ Responders to a 16-Wk Diet and Exercise Weight Loss Intervention. PLoS ONE 2016, 11, e0152545. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Torres, L.F.C.; Ong, K.L.; Shrestha, S.; Choteau, S.A.; Barter, P.J.; Clifton, P.; Rye, K.A. High-Density Lipoprotein-Associated MiR-223 Is Altered after Diet-Induced Weight Loss in Overweight and Obese Males. PLoS ONE 2016, 11, e0151061. [Google Scholar] [CrossRef] [PubMed]
- Lilja, S.; Stoll, C.; Krammer, U.; Hippe, B.; Duszka, K.; Debebe, T.; Höfinger, I.; König, J.; Pointner, A.; Haslberger, A. Five Days Periodic Fasting Elevates Levels of Longevity Related Christensenella and Sirtuin Expression in Humans. Int. J. Mol. Sci. 2021, 22, 2331. [Google Scholar] [CrossRef] [PubMed]
- Ravanidis, S.; Grundler, F.; de Toledo, F.W.; Dimitriou, E.; Tekos, F.; Skaperda, Z.; Kouretas, D.; Doxakis, E. Fasting-Mediated Metabolic and Toxicity Reprogramming Impacts Circulating MicroRNA Levels in Humans. Food Chem. Toxicol. 2021, 152, 112187. [Google Scholar] [CrossRef] [PubMed]
- Saini, S.K.; Singh, A.; Saini, M.; Gonzalez-Freire, M.; Leeuwenburgh, C.; Anton, S.D. Time-Restricted Eating Regimen Differentially Affects Circulatory MiRNA Expression in Older Overweight Adults. Nutrients 2022, 14, 1843. [Google Scholar] [CrossRef] [PubMed]
- Lopez, S.; Bermudez, B.; Montserrat-de la Paz, S.; Abia, R.; Muriana, F.J.G. A MicroRNA Expression Signature of the Postprandial State in Response to a High-Saturated-Fat Challenge. J. Nutr. Biochem. 2018, 57, 45–55. [Google Scholar] [CrossRef]
- Quintanilha, B.J.; Chaves, D.F.S.; Brasili, E.; Corrêa, T.A.F.; Capetini, V.C.; Ferreira, F.M.; Castro, I.A.; Hassimotto, N.M.A.; Rogero, M.M.; Lajolo, F.M. Ingestion of Orange Juice Prevents Hyperglycemia and Increases Plasma MiR-375 Expression. Clin. Nutr. ESPEN 2022, 47, 240–245. [Google Scholar] [CrossRef]
- Quintanilha, B.J.; Pinto Ferreira, L.R.; Ferreira, F.M.; Neto, E.C.; Sampaio, G.R.; Rogero, M.M. Circulating Plasma MicroRNAs Dysregulation and Metabolic Endotoxemia Induced by a High-Fat High-Saturated Diet. Clin. Nutr. 2020, 39, 554–562. [Google Scholar] [CrossRef]
- Gil-Zamorano, J.; Cofán, M.; de las Hazas, M.C.L.; García-Blanco, T.; García-Ruiz, A.; Doménech, M.; Serra-Mir, M.; Roth, I.; Valls-Pedret, C.; Rajaram, S.; et al. Interplay of Walnut Consumption, Changes in Circulating MiRNAs and Reduction in LDL-Cholesterol in Elders. Nutrients 2022, 14, 1473. [Google Scholar] [CrossRef]
- López de las Hazas, M.C.; Gil-Zamorano, J.; Cofán, M.; Mantilla-Escalante, D.C.; Garcia-Ruiz, A.; del Pozo-Acebo, L.; Pastor, O.; Yañez-Mo, M.; Mazzeo, C.; Serra-Mir, M.; et al. One-Year Dietary Supplementation with Walnuts Modifies Exosomal MiRNA in Elderly Subjects. Eur. J. Nutr. 2021, 60, 1999–2011. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.J.; Cardona-Alvarado, M.I.; Mercader, J.M.; Moreno-Navarrete, J.M.; Moreno, M.; Sabater, M.; Fuentes-Batllevell, N.; Ramírez-Chávez, E.; Ricart, W.; Molina-Torres, J.; et al. Circulating Profiling Reveals the Effect of a Polyunsaturated Fatty Acid-Enriched Diet on Common MicroRNAs. J. Nutr. Biochem. 2015, 26, 1095–1101. [Google Scholar] [CrossRef]
- Reis, B.Z.; Duarte, G.B.S.; Vargas-Mendez, E.; Ferreira, L.R.P.; Barbosa, F.; Cercato, C.; Rogero, M.M.; Cozzolino, S.M.F. Brazil Nut Intake Increases Circulating MiR-454-3p and MiR-584-5p in Obese Women. Nutr. Res. 2019, 67, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Salas-Huetos, A.; Moraleda, R.; Giardina, S.; Anton, E.; Blanco, J.; Salas-Salvadó, J.; Bulló, M. Effect of Nut Consumption on Semen Quality and Functionality in Healthy Men Consuming a Western-Style Diet: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2018, 108, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Daimiel, L.; Micó, V.; Valls, R.M.; Pedret, A.; Motilva, M.J.; Rubió, L.; Fitó, M.; Farrás, M.; Covas, M.I.; Solá, R.; et al. Impact of Phenol-Enriched Virgin Olive Oils on the Postprandial Levels of Circulating MicroRNAs Related to Cardiovascular Disease. Mol. Nutr. Food Res. 2020, 64, e2000049. [Google Scholar] [CrossRef] [PubMed]
- D’Amore, S.; Vacca, M.; Cariello, M.; Graziano, G.; D’Orazio, A.; Salvia, R.; Sasso, R.C.; Sabbà, C.; Palasciano, G.; Moschetta, A. Genes and MiRNA Expression Signatures in Peripheral Blood Mononuclear Cells in Healthy Subjects and Patients with Metabolic Syndrome after Acute Intake of Extra Virgin Olive Oil. Biochim. Biophys. Acta 2016, 1861, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Cannataro, R.; Perri, M.; Gallelli, L.; Caroleo, M.C.; De Sarro, G.; Cione, E. Ketogenic Diet Acts on Body Remodeling and MicroRNAs Expression Profile. Microrna 2019, 8, 116–126. [Google Scholar] [CrossRef]
- Desgagné, V.; Guay, S.P.; Guérin, R.; Corbin, F.; Couture, P.; Lamarche, B.; Bouchard, L. Variations in HDL-Carried MiR-223 and MiR-135a Concentrations after Consumption of Dietary Trans Fat Are Associated with Changes in Blood Lipid and Inflammatory Markers in Healthy Men—An Exploratory Study. Epigenetics 2016, 11, 438–448. [Google Scholar] [CrossRef]
- Ferrero, G.; Carpi, S.; Polini, B.; Pardini, B.; Nieri, P.; Impeduglia, A.; Grioni, S.; Tarallo, S.; Naccarati, A. Intake of Natural Compounds and Circulating MicroRNA Expression Levels: Their Relationship Investigated in Healthy Subjects With Different Dietary Habits. Front. Pharmacol. 2021, 11, 619200. [Google Scholar] [CrossRef]
- Liu, T.; Gatto, N.M.; Chen, Z.; Qiu, H.; Lee, G.; Duerksen-Hughes, P.; Fraser, G.; Wang, C. Vegetarian Diets, Circulating MiRNA Expression and Healthspan in Subjects Living in the Blue Zone. Precis. Clin. Med. 2020, 3, 245–259. [Google Scholar] [CrossRef]
- Tarallo, S.; Pardini, B.; Mancuso, G.; Rosa, F.; Di Gaetano, C.; Rosina, F.; Vineis, P.; Naccarati, A. MicroRNA Expression in Relation to Different Dietary Habits: A Comparison in Stool and Plasma Samples. Mutagenesis 2014, 29, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Tarallo, S.; Ferrero, G.; De Filippis, F.; Francavilla, A.; Pasolli, E.; Panero, V.; Cordero, F.; Segata, N.; Grioni, S.; Pensa, R.G.; et al. Stool MicroRNA Profiles Reflect Different Dietary and Gut Microbiome Patterns in Healthy Individuals. Gut 2022, 71, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Fontalba-Romero, M.I.; López-Enriquez, S.; Lago-Sampedro, A.; Garcia-Escobar, E.; Pastori, R.L.; Domínguez-Bendala, J.; Alvarez-Cubela, S.; Valdés, S.; Rojo-Martinez, G.; García-Fuentes, E.; et al. Association between the Mediterranean Diet and Metabolic Syndrome with Serum Levels of MiRNA in Morbid Obesity. Nutrients 2021, 13, 436. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, F.; Mitchell, C.J.; Milan, A.M.; Schierding, W.; Zeng, N.; Sharma, P.; Mitchell, S.M.; D’Souza, R.F.; Knowles, S.O.; Roy, N.C.; et al. Comprehensive Profiling of the Circulatory MiRNAome Response to a High Protein Diet in Elderly Men: A Potential Role in Inflammatory Response Modulation. Mol. Nutr. Food Res. 2019, 63, e1800811. [Google Scholar] [CrossRef] [PubMed]
- Shin, P.K.; Kim, M.S.; Park, S.J.; Kwon, D.Y.; Kim, M.J.; Yang, H.J.; Kim, S.H.; Kim, K.; Chun, S.; Lee, H.J.; et al. A Traditional Korean Diet Alters the Expression of Circulating MicroRNAs Linked to Diabetes Mellitus in a Pilot Trial. Nutrients 2020, 12, 2558. [Google Scholar] [CrossRef] [PubMed]
- Tutino, V.; De Nunzio, V.; Milella, R.A.; Gasparro, M.; Cisternino, A.M.; Gigante, I.; Lanzilotta, E.; Iacovazzi, P.A.; Lippolis, A.; Lippolis, T.; et al. Impact of Fresh Table Grape Intake on Circulating MicroRNAs Levels in Healthy Subjects: A Significant Modulation of Gastrointestinal Cancer-Related Pathways. Mol. Nutr. Food Res. 2021, 65, 2100428. [Google Scholar] [CrossRef]
- Capetini, V.C.; Quintanilha, B.J.; de Oliveira, D.C.; Nishioka, A.H.; de Matos, L.A.; Ferreira, L.R.P.; Ferreira, F.M.; Sampaio, G.R.; Hassimotto, N.M.A.; Lajolo, F.M.; et al. Blood Orange Juice Intake Modulates Plasma and PBMC MicroRNA Expression in Overweight and Insulin-Resistant Women: Impact on MAPK and NFκB Signaling Pathways. J. Nutr. Biochem. 2023, 112, 109240. [Google Scholar] [CrossRef]
- Desvignes, T.; Batzel, P.; Berezikov, E.; Eilbeck, K.; Eppig, J.T.; McAndrews, M.S.; Singer, A.; Postlethwait, J.H. MicroRNA Nomenclature: A View Incorporating Genetic Origins, Biosynthetic Pathways, and Sequence Variants. Trends Genet. 2015, 31, 613. [Google Scholar] [CrossRef]
- Ameling, S.; Kacprowski, T.; Chilukoti, R.K.; Malsch, C.; Liebscher, V.; Suhre, K.; Pietzner, M.; Friedrich, N.; Homuth, G.; Hammer, E.; et al. Associations of Circulating Plasma MicroRNAs with Age, Body Mass Index and Sex in a Population-Based Study. BMC Med. Genom. 2015, 8, 61. [Google Scholar] [CrossRef]
- Ban, E.; Song, E.J. Considerations and Suggestions for the Reliable Analysis of MiRNA in Plasma Using QRT-PCR. Genes 2022, 13, 328. [Google Scholar] [CrossRef]
- Wong, R.K.Y.; MacMahon, M.; Woodside, J.V.; Simpson, D.A. A Comparison of RNA Extraction and Sequencing Protocols for Detection of Small RNAs in Plasma. BMC Genom. 2019, 20, 446. [Google Scholar] [CrossRef] [PubMed]
- Meerson, A.; Traurig, M.; Ossowski, V.; Fleming, J.M.; Mullins, M.; Baier, L.J. Human Adipose MicroRNA-221 Is Upregulated in Obesity and Affects Fat Metabolism Downstream of Leptin and TNF-α. Diabetologia 2013, 56, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Shan, A.; Su, Y.; Cheng, Y.; Ji, H.; Yang, Q.; Lei, Y.; Liu, B.; Wang, W.; Ning, G.; et al. MiR-221/222 Inhibit Insulin Production of Pancreatic β-Cells in Mice. Endocrinology 2020, 161, bqz027. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of Tissue-Specific MicroRNAs from Mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, H.X.; Jena, P.K.; Sheng, L.; Ali, M.R.; Wan, Y.J.Y. MiR-22 Inhibition Reduces Hepatic Steatosis via FGF21 and FGFR1 Induction. JHEP Rep. 2020, 2, 100093. [Google Scholar] [CrossRef]
- Panella, R.; Petri, A.; Desai, B.N.; Fagoonee, S.; Cotton, C.A.; Nguyen, P.K.; Lundin, E.M.; Wagshal, A.; Wang, D.Z.; Näär, A.M.; et al. MicroRNA-22 Is a Key Regulator of Lipid and Metabolic Homeostasis. Int. J. Mol. Sci. 2023, 24, 12870. [Google Scholar] [CrossRef]
- Agbu, P.; Carthew, R.W. MicroRNA-Mediated Regulation of Glucose and Lipid Metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef]
- Hartig, S.M.; Hamilton, M.P.; Bader, D.A.; McGuire, S.E. The MicroRNA Interactome in Metabolic Homeostasis. Trends Endocrinol. Metab. 2015, 26, 733. [Google Scholar] [CrossRef]
- Kabekkodu, S.P.; Shukla, V.; Varghese, V.K.; D’Souza, J.; Chakrabarty, S.; Satyamoorthy, K. Clustered MiRNAs and Their Role in Biological Functions and Diseases. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1955–1986. [Google Scholar] [CrossRef]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. In Methods Mol Biol; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1509, pp. 1–10. [Google Scholar]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Zhou, S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y.; Cai, L. MiRNAS in Cardiovascular Diseases: Potential Biomarkers, Therapeutic Targets and Challenges. Acta Pharmacol. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Guo, X. The Clinical Potential of Circulating MicroRNAs in Obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Foscolou, A.; Critselis, E.; Panagiotakos, D. Olive Oil Consumption and Human Health: A Narrative Review. Maturitas 2018, 118, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Nocella, C.; Cammisotto, V.; Fianchini, L.; D’Amico, A.; Novo, M.; Castellani, V.; Stefanini, L.; Violi, F.; Carnevale, R. Extra Virgin Olive Oil and Cardiovascular Diseases: Benefits for Human Health. Endocr. Metab. Immune Disord.-Drug Targets 2018, 18, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Marrone, G.; Guerriero, C.; Palazzetti, D.; Lido, P.; Marolla, A.; Di Daniele, F.; Noce, A. Vegan Diet Health Benefits in Metabolic Syndrome. Nutrients 2021, 13, 817. [Google Scholar] [CrossRef] [PubMed]
- He, P.P.; OuYang, X.P.; Li, Y.; Lv, Y.C.; Wang, Z.B.; Yao, F.; Xie, W.; Tan, Y.L.; Li, L.; Zhang, M.; et al. MicroRNA-590 Inhibits Lipoprotein Lipase Expression and Prevents Atherosclerosis in ApoE Knockout Mice. PLoS ONE 2015, 10, e0138788. [Google Scholar] [CrossRef]
- He, P.P.; Ouyang, X.P.; Tang, Y.Y.; Liao, L.; Wang, Z.B.; Lv, Y.C.; Tian, G.P.; Zhao, G.J.; Huang, L.; Yao, F.; et al. MicroRNA-590 Attenuates Lipid Accumulation and pro-Inflammatory Cytokine Secretion by Targeting Lipoprotein Lipase Gene in Human THP-1 Macrophages. Biochimie 2014, 106, 81–90. [Google Scholar] [CrossRef]
- Liu, X.L.; Cao, H.X.; Wang, B.C.; Xin, F.Z.; Zhang, R.N.; Zhou, D.; Yang, R.X.; Zhao, Z.H.; Pan, Q.; Fan, J.G. MiR-192-5p Regulates Lipid Synthesis in Non-Alcoholic Fatty Liver Disease through SCD-1. World J. Gastroenterol. 2017, 23, 8140–8151. [Google Scholar] [CrossRef]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-Associated Exosomal MiRNAs Modulate Glucose and Lipid Metabolism in Mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Gao, X.; Zhang, R.; Zhang, Y.; Liang, H.; Xu, C.; Du, W.; Zhang, Y.; Liu, X.; et al. MicroRNA-328 as a Regulator of Cardiac Hypertrophy. Int. J. Cardiol. 2014, 173, 268–276. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.D.; Rong, J.; Huan, T.; Lacey, S.; Tanriverdi, K.; Munson, P.J.; Larson, M.G.; Joehanes, R.; Murthy, V.; Shah, R.; et al. Messenger RNA and MicroRNA Transcriptomic Signatures of Cardiometabolic Risk Factors. BMC Genom. 2017, 18, 139. [Google Scholar] [CrossRef]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in Metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, A.; Ezekiel, U. Phytochemical Modulation of MiRNAs in Colorectal Cancer. Medicines 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Larrosa, M.; Yáñez-Gascón, M.J.; Dávalos, A.; Gil-Zamorano, J.; Gonzálvez, M.; García-Almagro, F.J.; Ruiz Ros, J.A.; Tomás-Barberán, F.A.; Espín, J.C.; et al. One-Year Supplementation with a Grape Extract Containing Resveratrol Modulates Inflammatory-Related MicroRNAs and Cytokines Expression in Peripheral Blood Mononuclear Cells of Type 2 Diabetes and Hypertensive Patients with Coronary Artery Disease. Pharmacol. Res. 2013, 72, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Sanchita; Trivedi, R.; Asif, M.H.; Trivedi, P.K. Dietary Plant MiRNAs as an Augmented Therapy: Cross-Kingdom Gene Regulation. RNA Biol. 2018, 15, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W. XenomiRs and MiRNA Homeostasis in Health and Disease: Evidence That Diet and Dietary MiRNAs Directly and Indirectly Influence Circulating MiRNA Profiles. RNA Biol. 2012, 9, 1147. [Google Scholar] [CrossRef]
- Jia, M.; He, J.; Bai, W.; Lin, Q.; Deng, J.; Li, W.; Bai, J.; Fu, D.; Ma, Y.; Ren, J.; et al. Cross-Kingdom Regulation by Dietary Plant MiRNAs: An Evidence-Based Review with Recent Updates. Food Funct. 2021, 12, 9549–9562. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front. Nutr. 2021, 8, 586564. [Google Scholar] [CrossRef]
- Li, J.; Lei, L.; Ye, F.; Zhou, Y.; Chang, H.; Zhao, G. Nutritive Implications of Dietary MicroRNAs: Facts, Controversies, and Perspectives. Food Funct. 2019, 10, 3044–3056. [Google Scholar] [CrossRef]
- Title, A.; Denzler, R.; Stoffel, M. Reply to Diet-Responsive MicroRNAs Are Likely Exogenous. J. Biol. Chem. 2015, 290, 25198. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.W.; Hale, A.E.; Isaacs, S.K.; Baggish, A.L.; Chan, S.Y. Ineffective Delivery of Diet-Derived MicroRNAs to Recipient Animal Organisms. RNA Biol. 2013, 10, 1107–1116. [Google Scholar] [CrossRef]
- Cintio, M.; Polacchini, G.; Scarsella, E.; Montanari, T.; Stefanon, B.; Colitti, M. MicroRNA Milk Exosomes: From Cellular Regulator to Genomic Marker. Animals 2020, 10, 1126. [Google Scholar] [CrossRef]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs Are Absorbed in Biologically Meaningful Amounts from Nutritionally Relevant Doses of Cow Milk and Affect Gene Expression in Peripheral Blood Mononuclear Cells, HEK-293 Kidney Cell Cultures, and Mouse Livers. J. Nutr. 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Izumi, H.; Sekine, K.; Ochiya, T. MicroRNA as a New Immune-Regulatory Agent in Breast Milk. Silence 2010, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Du, X.; Li, J.; Lönnerdal, B. Human Milk Exosomes and Their MicroRNAs Survive Digestion in Vitro and Are Taken up by Human Intestinal Cells. Mol. Nutr. Food Res. 2017, 61, 1700082. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Lozano, E.; Sebastián-Valles, F.; Knott-Torcal, C. Circulating MicroRNAs in Breast Milk and Their Potential Impact on the Infant. Nutrients 2020, 12, 66. [Google Scholar] [CrossRef]
- Langley-Evans, S.C. Nutrition in Early Life and the Programming of Adult Disease: A Review. J. Hum. Nutr. Diet. 2015, 28, 1–14. [Google Scholar] [CrossRef]
- Carolan-Olah, M.; Duarte-Gardea, M.; Lechuga, J. A Critical Review: Early Life Nutrition and Prenatal Programming for Adult Disease. J. Clin. Nurs. 2015, 24, 3716–3729. [Google Scholar] [CrossRef]
- Picó, C.; Palou, M.; Priego, T.; Sánchez, J.; Palou, A.; Sanchez, J.; Palou, A. Metabolic Programming of Obesity by Energy Restriction during the Perinatal Period: Different Outcomes Depending on Gender and Period, Type and Severity of Restriction. Front. Physiol. 2012, 3, 436. [Google Scholar] [CrossRef]
- Alonso-Bernáldez, M.; Asensio, A.; Palou-March, A.; Sánchez, J.; Palou, A.; Serra, F.; Palou, M. Breast Milk MicroRNAs Related to Leptin and Adiponectin Function Can Be Modulated by Maternal Diet and Influence Offspring Phenotype in Rats. Int. J. Mol. Sci. 2022, 23, 7237. [Google Scholar] [CrossRef] [PubMed]
- Pomar, C.A.; Serra, F.; Palou, A.; Sánchez, J. Lower MiR-26a Levels in Breastmilk Affect Gene Expression in Adipose Tissue of Offspring. FASEB J. 2021, 35, e21924. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C.; Stremmel, W.; Weiskirchen, R.; John, S.M.; Schmitz, G. Exosome-Derived MicroRNAs of Human Milk and Their Effects on Infant Health and Development. Biomolecules 2021, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- Golan-Gerstl, R.; Elbaum Shiff, Y.; Moshayoff, V.; Schecter, D.; Leshkowitz, D.; Reif, S. Characterization and Biological Function of Milk-Derived MiRNAs. Mol. Nutr. Food Res. 2017, 61, 1700009. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-Related MicroRNAs Are Abundant in Breast Milk Exosomes. Int. J. Biol. Sci. 2012, 8, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.B.; Chernausek, S.D.; Garman, L.D.; Pezant, N.P.; Plows, J.F.; Kharoud, H.K.; Demerath, E.W.; Fields, D.A. Human Milk Exosomal MicroRNA: Associations with Maternal Overweight/Obesity and Infant Body Composition at 1 Month of Life. Nutrients 2021, 13, 1091. [Google Scholar] [CrossRef] [PubMed]
- Torres, L.F.; Cogliati, B.; Otton, R. Green Tea Prevents NAFLD by Modulation of MiR-34a and MiR-194 Expression in a High-Fat Diet Mouse Model. Oxid. Med. Cell. Longev. 2019, 2019, 4168380. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
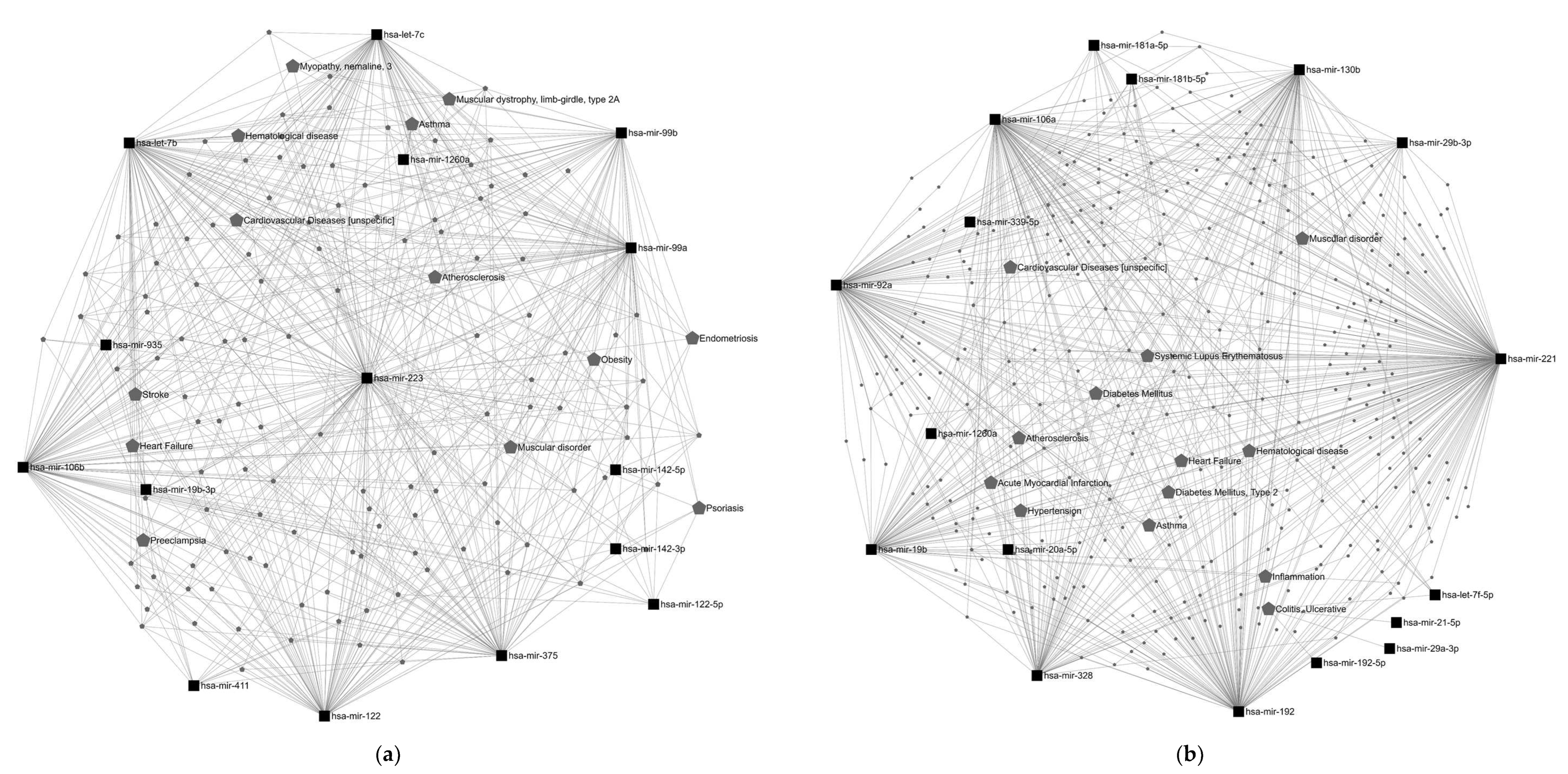{kind=link}
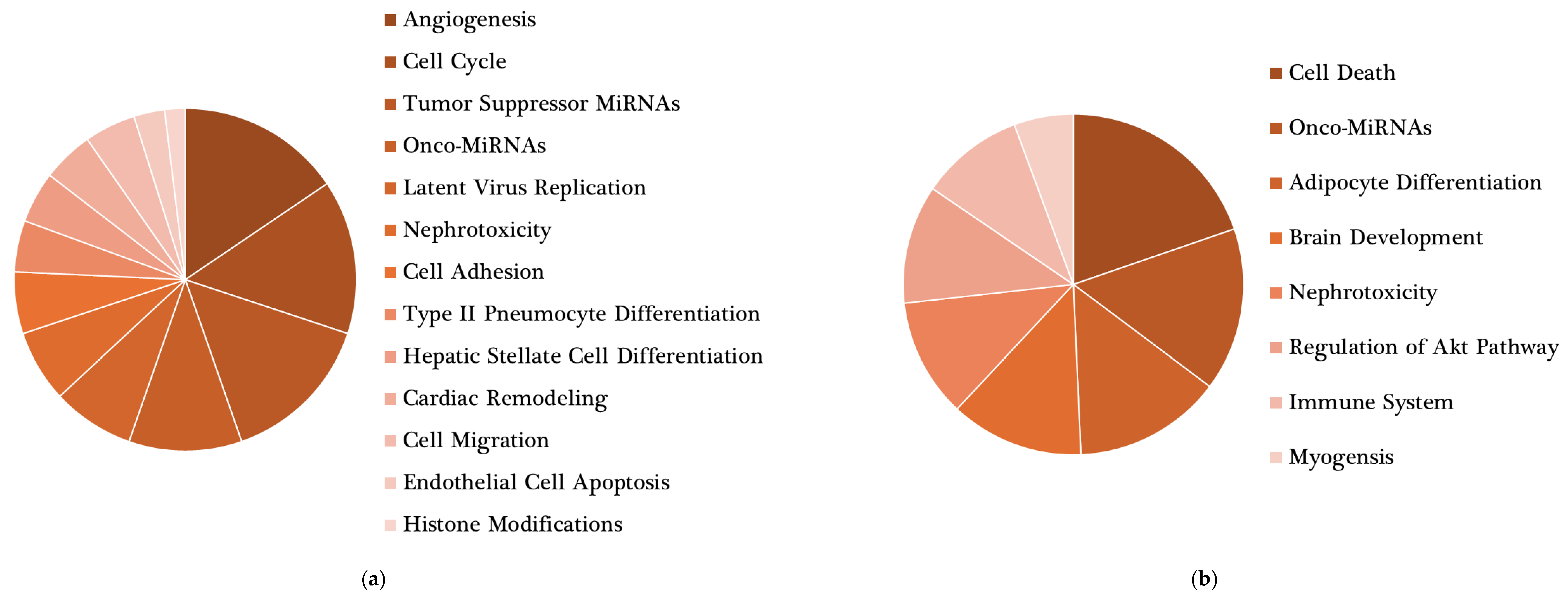{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| Studies conducted in humans | Studies not conducted in humans |
| Observational or interventional studies | Reviews and other design types |
| Studies evaluating dietary modulation of miRNA expression | Studies evaluating supplementation rather than food intake |
| Studies performed in adults | Population age under 18 |
| Studies on healthy population with or without obesity | Study population with any other disease |
| Author and Publication Date | Population (F/M) | Health Status | Study Design | Dietary Strategy | Method of Analysis | Sample Analyzed | Outcomes |
|---|---|---|---|---|---|---|---|
| Giardina, 2019 [33] | 103 adults (82/21), 30–60 years | With overweight/obesity (27–35 kg/m2) | Randomized, parallel-group, controlled trial | 6-month study, three groups of energy-restricted diets (500 kcal/day restriction): moderate-carbohydrate and low-glycemic-index diet (LGI), moderate-carbohydrate and high-glycemic-index diet (HGI) and low-fat and high-glycemic-index diet (LF) | Array screening and validation using qPCR | Plasma | LGI vs. HGI: ↓ hsa-miR-361 After LGI: ↓ hsa-miR-139-3p ↓ hsa-miR-411 ↓ hsa-miR-432 ↓ hsa-miR-99b ↓ hsa-miR-340 ↓ hsa-miR-423-5p ↓ hsa-miR-361 ↓ hsa-let-7c After HGI: ↓ hsa-miR-139-3p ↓ hsa-miR-340 After LF: ↓ hsa-miR-139-3p ↓ hsa-miR-432 ↓ hsa-miR-423-5p |
| Margolis, 2017 [34] | 16 men, 60–75 years | With overweight (25–35 kg/m2) | Randomized, parallel-group, triple-blinded trial | 35-day study: 7-day weight maintenance and 28-day 30% energy restriction periods | qPCR | Serum | Energy restriction: ↑ miR-133a-3p ↑ miR-133b |
| Assmann, 2020 [35] | 103 adults | Intervention groups with obesity (30–40 kg/m2) and control group with normal weight | Randomized, parallel-group trial | 16-week weight loss intervention, three groups with 30% energy restriction: moderately high-protein diet (n = 38, 40% carbohydrate, 30% protein, 30% fat), low-fat diet (n = 40, 60% carbohydrate, 18% protein, 22% fat) and control (n = 25) | qPCR array | Plasma | Responders vs. non-responders to low-fat diet: ↓ hsa-miR-130a-3p ↓ hsa-miR-142-5p ↓ hsa-miR-144-5p ↓ hsa-miR-15a-5p ↓ hsa-miR-221-3p ↓ hsa-miR-29c-3p ↑ hsa-miR-22-3p |
| Duggan, 2022 [36] | 192 postmenopausal women, 50–75 years | With overweight/obesity | Randomized, parallel-group, single-blinded, controlled trial | 12-month weight loss intervention in four groups: restricted diet, exercise, diet + exercise and control (no intervention) | NanoString nCounter technology | Plasma | Weight loss intervention vs. control: ↓ miR-122 |
| Heianza, 2022 [37] | 495 adults | Healthy and with overweight | Randomized trial | 2-year study, four groups of weight loss diets: low-fat average-protein diet, low-fat high-protein diet, high-fat average-protein diet and high-fat high-protein diet | RNA-seq | Plasma | High-fat and high-protein vs. low-fat and average-protein: ↓ hsa-miR-128-1-5p |
| Hess, 2020 [38] | 85 adults (55/30), 18–60 years | With overweight/obesity (28–45 kg/m2) | Randomized, parallel-group, double-blinded trial | 12-week study, two weight loss intervention groups, both with an energy deficit of 500 kcal/day: fiber supplementation (20 g/day) and control (no supplement) | qPCR array | Serum | After both diets: ↓ hsa-miR-122-5p ↓ hsa-miR-193a-5p ↑ hsa-miR-126a-3p ↑ hsa-miR-222-3p |
| Jayasooriya, 2022 [39] | 27 adults, 18–60 years | With overweight (25–34.9 kg/m2) | Interventional single-arm pilot study | 6-week weight loss intervention: diet (250 kcal/d energy restriction) and exercise | miRNA sensor iLluminate | Serum | Post vs. pre: ↓ hsa-let-7b ↓ hsa-miR-99a |
| Kristensen, 2017 [40] | 19 adults (10/9) | With morbid obesity (≥40 kg/m2) | Interventional study | 15-week weight loss intervention: exercise and hypocaloric diet | Array screening and validation using qPCR | Subcutaneous adipose tissue | ↑ hsa-miR-29a-3p ↑ hsa-miR-29a-5p ↓ hsa-miR-20b-5p |
| Milagro, 2013 [41] | 10 women | With obesity (35.6 kg/m2) | Interventional study | 8-week weight loss intervention (800–880 kcal/day). Two groups: responders (weight loss > 5%) and non-responders (weight loss < 5%) | RNA-seq screening and validation using qPCR | PBMC | Non-responders: ↑ hsa-miR-935 ↑ hsa-miR-4772-3p ↓ hsa-miR-223 ↓ hsa-miR-224 ↓ hsa-miR-376b |
| Müller, 2020 [42] | 46 adults (26/20), 18–72 years | With obesity (>30 kg/m2) | Interventional study | 3-month weight loss intervention: 800 kcal/day (only protein shake) | RNA-seq screening and validation using qPCR | Plasma | ↓ hsa-miR-25-3p ↓ hsa-miR-93-5p ↓ hsa-miR-106b-3p |
| Parr, 2016 [43] | 40 adults (26/14), 35–59 years | With obesity (27–40 kg/m2) | Randomized, parallel-group, controlled trial | 16-week weight loss intervention: 250 kcal/day energy restriction and exercise. Two groups: HiRes (>10% body mass loss, n = 22) and LoRes (<5% body mass loss, n = 18) | qPCR array | Plasma | LoRes vs. HiRes: ↑ hsa-miR-935 Both groups: ↑ hsa-miR-221-3p ↑ hsa-miR-223-3p |
| Tabet, 2016 [44] | 47 men, 20–65 years | With obesity (32 kg/m2) | Randomized, parallel-group, controlled trial | 12-week weight loss intervention. Two groups of hypocaloric diets: high-protein diet (30% of energy, n = 20) and normal-protein diet (20% of energy, n = 27) | qPCR | HDL-fraction of serum samples | High-protein diet: ↓ miR-223 |
| Lilja, 2021 [45] | 54 adults (35/16), 23–75 years | Healthy | Interventional study | 5-day study, two groups: fasting (5 days of only liquids) and control (non-fasting) | qPCR | Stool | Fasting: ↑ let-7b-5p ↓ miR-34a-5p |
| Ravanidis, 2021 [46] | 32 adults (10/22), 18–70 years | With overweight/obesity (28 kg/m2) | Interventional single-arm study | 10-day fasting period (250 kcal/day) | qPCR | Plasma | ↓ hsa-miR-19b-3p ↓ hsa-miR-22-3p ↓ hsa-miR-142-3p ↓ hsa-miR-143-3p ↓ hsa-miR-145-5p ↑ hsa-miR-122-5p ↑ hsa-miR-126-3p |
| Saini, 2022 [47] | 9 older adults (6/3), ≥65 years | With overweight | Interventional single-arm study | 4 weeks with 16 h per day of fasting | RNA-seq | Serum | After intervention: ↑ miR-623 ↑ miR-4303 ↑ miR-7162-3p ↑ miR-411-5p ↑ miR-5682 ↑ miR-4513 ↓ miR-4649-5p ↓ miR-2467-3p ↓ miR-543 ↓ miR-301a-3p ↓ miR-3132 ↓ miR-19a-5p ↓ miR-495-3p ↓ miR-4761-3p |
| Lopez, 2018 [48] | 9 men, 18–23 years | Healthy | Randomized, crossover, double-blinded trial | A single high-saturated-fat meal (800 kcal, 77% fat, 23% carbohydrate) | Array screening and validation using qPCR | PBMC | Postprandial: ↓ hsa-miR-613 ↓ hsa-miR-629-3p ↓ hsa-miR-24-2-5p ↓ hsa-miR-555 ↓ hsa-miR-148a-5p ↓ hsa-miR-621 ↓ hsa-miR-875-3p ↓ hsa-miR-513c-5p ↓ hsa-miR-1226 ↑ hsa-miR-653 ↑ hsa-miR-19b-1-5p ↑ hsa-miR-363-5p ↑ hsa-miR-885-3p ↑ hsa-miR-339-3p ↑ hsa-miR-938 ↑ hsa-miR-148b-5p ↑ hsa-miR-593-5p ↑ hsa-miR-200b-5p |
| Quintanilha, 2022 [49] | 12 adults (7/5), 25–45 years | Healthy | Randomized, crossover trial | A single high-fat high-carbohydrate meal + water/orange juice/isocaloric beverage with 1-week washouts | qPCR | Plasma | Meal + orange juice vs. water: ↑ hsa-miR-375 Meal + glucose vs. water: ↓ hsa-miR-205-3p |
| Quintanilha, 2020 [50] | 11 women, 20–40 years | Healthy | Interventional trial | A single high-fat high-saturated meal (1067 kcal) | qPCR array | Plasma | Postprandial: ↑ hsa-miR-200c-3p ↑ hsa-miR-143-5p ↑ hsa-miR-200b-3p ↑ hsa-miR-143-3p ↑ hsa-miR-375 ↑ hsa-miR-145-5p ↓ hsa-miR-1260a ↓ hsa-miR-92b-3p ↓ hsa-miR-205-5p |
| Gil-Zamorano, 2022 [51] | 8 adults, 63–79 years | Healthy | Randomized, parallel-group, single-blinded, controlled trial | 1-year study, two groups: walnut supplementation (30–60 g/day) and control (abstaining from walnuts) | Array screening and validation using qPCR | Serum | ↑ hsa-miR-551a |
| López de las Hazas, 2021 [52] | 211 subjects, 63–79 years | Healthy | Randomized, parallel-group, single-blinded, controlled trial | 1-year study, two groups: walnut supplementation (n = 101, 30–60 g/day) and control (n = 110, abstaining from walnuts) | Screening in 40 pools of samples and validation using qPCR | Plasma | Walnuts: ↑ hsa-miR-32-5p ↑ hsa-miR-29b-3p |
| Ortega, 2015 [53] | 30 adults (22/8), 30–50 years | With obesity (30–35 kg/m2) | Interventional study | 8-week study: normocaloric diet enriched with PUFA (30 g/day almonds and walnuts) | Array screening and validation using qPCR | Plasma | ↓ miR-328 ↓ miR-330-3p ↓ miR-221 ↓ miR-125a-5p ↑ miR-192 ↑ miR-486-5p ↑ miR-19b ↑ miR-106a ↑ miR-130b ↑ miR-18a ↑ miR-769-5p |
| Reis, 2019 [54] | 54 women, 18–55 years | With overweight/obesity (≥27.5 kg/m2) | Randomized, parallel-group, controlled trial | 2-month study. Two groups: Brazil nut (1 Brazil nut/day, n = 29) and control (no Brazil nuts, n = 25) | qPCR | Plasma | Brazil nut intake: ↑ miR-454-3p ↑ miR-584-5p |
| Salas-Huetos, 2018 [55] | 98 men, 18–35 years | Healthy | Randomized, parallel-group, single-blinded, controlled trial | 14-week study, two groups: nuts (Western diet + 60 g/day nuts) and control (Western diet avoiding nuts) | Array for screening and validation | Sperm | Nuts: ↓ hsa-miR-34b-3p |
| Daimiel, 2020 [56] | 12 adults (6/6), 22–60 years | Healthy | Randomized, crossover, double-blinded, controlled trial | 30 mL of 3 polyphenol-enriched EVOOs after 12 h of fasting: low-EVOO (250 mg/kg of oil), medium-EVOO (500 mg/kg) and high-EVOO (750 mg/kg) diets | qPCR array | Plasma | EVOO: ↓ l hsa-let-7e-5p ↓ hsa-miR-328a-3p ↓ hsa-miR-10a-5p ↓ hsa-miR-21-5p ↓ hsa-miR-26b-5p ↑ hsa-miR-17-5p ↑ hsa-miR-20a-5p ↑ hsa-miR-192-5p |
| D’Amore, 2016 [57] | 24 adults (12/12) | 12 healthy and 12 with metabolic syndrome | Interventional study | Two interventions: single intake of high-polyphenol EVOO and low-polyphenol EVOO (50 mL) after 12 h fasting and a 1-week washout with no olive oil intake | Array screening and validation using qPCR | PBMC | High-polyphenol EVOO in healthy adults: ↑ miR-23b-3p ↑ miR-519b-39 ↓ miR-146b-5p ↓ miR-19a-3p ↓ miR-181b-5p ↓ miR-107 ↓ miR-769-5p ↓ miR-192-5p |
| Cannataro, 2019 [58] | 36 adults (18/18), 31–58 years | With obesity (>30 kg/m2) | Interventional study | 6 weeks of a ketogenic diet | NanoString nCounter technology | Serum | Ketogenic diet: ↑ hsa-let-7b-5p ↑ hsa-miR-143-3p ↓ hsa-miR-504-5p |
| Desgagné, 2016 [59] | 9 men, 20–59 years | Healthy | Randomized, crossover, double-blinded, controlled trial | Three 4-week interventions with 3-week washout periods: high-iTFA (10.2 g industrial TFA/2500 kcal, 3.7% energy), high-rTFA (10.2 g dairy and meat TFA/2500 kcal, 3.7% energy) and low-TFA (2.2 g/2500 kcal, 0.8% energy) | qPCR | HDL plasma-fraction | No significant results |
| Ferrero, 2021 [60] | 120 adults (72/48) | Healthy | Observational study | Equal % of vegans, vegetarians and omnivores, diet > 1 year. Food frequency questionnaires | RNA-seq | Plasma | No significant results |
| Liu, 2020 [61] | 96 adults (53/43), ≥60 years | Healthy | Observational study | 31 non-vegetarians, 15 vegans, 32 lacto-vegetarians and 18 semi-vegetarians. Food frequency questionnaires | RNA-seq | Plasma | Vegetarians: ↑ hsa-miR-3661 ↑ hsa-miR-320c ↑ hsa-miR-29a-3p ↑ hsa-miR-320b ↑ hsa-miR-204-3p ↓ hsa-miR-132-5p |
| Tarallo, 2014 [62] | 24 adults (15/9), 21–60 years | Healthy | Observational study | 8 vegans, 8 vegetarians, 8 omnivorous | qPCR | Stool and plasma | Vegan/vegetarian: ↑ hsa-miR-92a |
| Tarallo, 2022 [63] | 120 adults (72/48) | Healthy | Observational study | Vegan, vegetarian and omnivorous, diet >1 year. Food frequency questionnaires | RNA-seq | Plasma and stool | No significant results in plasma Vegan/vegetarians: ↓ hsa-miR-636 ↓ hsa-miR-4488-3p ↓ hsa-miR-4739 |
| Fontalba-Romero, 2021 [64] | 58 adults (41/17) | With morbid obesity (≥40 kg/m2) | Observational study | MEDAS questionnaire to determine the adherence to a Mediterranean diet | Array screening and validation using qPCR | Serum | High Med diet adherence: ↑ miR-590 |
| Humphreys, 2014 [17] | 23 adults (6/17), 50–75 years | Healthy | Randomized, crossover, controlled trial | Two 4-week interventions with 4-week entry and washouts: HRM (300 g/day meat) and HRM + HAMSB (300 g/day meat + 40 g/day butyrylated high amylose starch) | qPCR | Rectal mucosa | HRM: ↑ hsa-miR-19a-3p ↑ hsa-miR-19b-3p ↑ hsa-miR-21-5p HRM + HAMSB: ↓ hsa-miR-17-5p ↓ hsa-miR-19a-3p ↓ hsa-miR-19b-3p ↓ hsa-miR-20a-5p ↓ hsa-miR-92a |
| Ramzan, 2019 [65] | 31 men, ≥70 years | Healthy | Randomized, parallel-group, single-blinded trial | 10-week study, two groups: RDA (0.8 g protein/kg body weight/day) and 2RDA (1.6 g/kg body weight/day) | RNA-seq screening and validation using qPCR | Plasma | 2RDA: ↓ hsa-miR-125b-5p ↓ hsa-miR-100-5p ↓ hsa-miR-99a-5p ↓ hsa-miR-23b-3p ↓ hsa-miR-203a |
| Shin, 2020 [66] | 10 women, 50–60 years | With overweight 25–30 kg/m2) | Randomized, parallel-group trial | 2-week study. Two groups: k-diet (traditional Korean diet) and control (Westernized Korean diet) | Array screening and validation using qPCR | Plasma and saliva | K-diet, plasma: ↓ hsa-miR-26a-5p ↓ hsa-miR-126-3p Control, plasma: ↓ hsa-miR-25-3p K-diet, saliva: ↓ hsa-miR-92-3p ↓ hsa-miR-122a-5p Control, saliva: ↓ hsa-miR-31-5p |
| Tutino, 2021 [67] | 40 adults (29/11), 30–65 years | With overweight (25–30 kg/m2) | Randomized, parallel-group, single-blinded, controlled trial | 21-day study. Two groups: grape group (5 g/day fresh grape/kg body weight) and control (abstaining from grapes) | qPCR array | Serum | Grape group: ↑ hsa-miR-208a-3p ↑ hsa-miR-33a-5p ↓ hsa-miR-181a-5p ↓ hsa-miR-30e-5p ↓ hsa-miR-30d-5p ↓ hsa-miR-335-5p ↓ hsa-miR-222-3p ↓ hsa-miR-15a-5p ↓ hsa-miR-421 ↓ hsa-miR-339-5p ↓ hsa-miR-378a-3p ↓ hsa-let-7f-5p ↓ hsa-miR-29b-3p ↓ hsa-miR-106b-3p ↓ hsa-miR-324-5p ↓ hsa-miR-1260a ↓ hsa-miR-365a-3p ↓ hsa-miR-155-5p ↓ hsa-miR-335-3p ↓ hsa-miR-200c-3p |
| Capetini, 2023 [68] | 20 women, 18–40 years | With overweight (25–29.9 kg/m2) | Interventional study | 4 weeks consuming 500 mL/d of blood orange juice | Array screening and validation using qPCR | Plasma and PBMC | Plasma: ↑ hsa-miR-144-3p PBMC: ↑ hsa-miR-144-3p ↑ hsa-miR-424-5p ↑ hsa-miR-130b-3p ↓ hsa-let-7f-5p ↓ hsa-miR-126-3p |
| miRNA | Energy Restriction | Fasting | High-Fat Meal | Nuts | EVOO | Vegetarian Diet | Grape | Orange Juice | Red Meat |
|---|---|---|---|---|---|---|---|---|---|
| let-7 | ↓ plasma [33] ↓ serum [39] | ↑ stool [45] | ↓ plasma [56] | ↓ serum [67] | ↓ PBMC [68] | ||||
| miR-19 | ↓ plasma [46] ↓ serum [47] | ↑ PBMC [48] | ↑ plasma [53] | ↓ PBMC [57] | ↑ rectal mucosa [17] | ||||
| miR-29 | ↓ plasma [35] ↑ adipose tissue [40] | ↑ plasma [52] | ↑ plasma [61] | ↓ serum [67] | |||||
| miR-92 | ↓ plasma [50] | ↑ plasma [62] | |||||||
| miR-122 | ↓ serum [38] ↓ plasma [36] | ↑ plasma [46] | |||||||
| miR-126 | ↑ serum [38] | ↑ plasma [46] | ↓ PBMC [68] | ||||||
| miR-20 | ↓ adipose tissue [40] | ↑ plasma [56] | |||||||
| miR-99 | ↓ plasma [33] ↓ serum [39] | ||||||||
| miR-106 | ↓ plasma [42] | ↑ plasma [53] | ↓ serum [67] | ||||||
| miR-130 | ↓ plasma [35] | ↑ plasma [53] | ↑ PBMC [68] | ||||||
| miR-143 | ↓ plasma [46] | ↑ plasma [50] | |||||||
| miR-192 | ↑ plasma [53] | ↑ plasma [56] ↓ PBMC [57] | |||||||
| miR-200 | ↑ PBMC [48] ↑ plasma [50] | ↓ serum [67] | |||||||
| miR-221 | ↓ plasma [35] ↑ plasma [43] | ↓ plasma [53] | |||||||
| miR-223 | ↓ PBMC [41] ↓ serum HDL [44] ↑ plasma [43] | ||||||||
| miR-15 | ↓ plasma [35] | ↓ serum [67] | |||||||
| miR-21 | ↓ plasma [56] | ↑ rectal mucosa [17] | |||||||
| miR-22 | ↑ plasma [35] | ↓ plasma [46] | |||||||
| miR-34 | ↓ stool [45] | ↓ sperm [55] | |||||||
| miR-142 | ↓ plasma [35] | ↓ plasma [46] | |||||||
| miR-144 | ↓ plasma [35] | ↑ PBMC [68] | |||||||
| miR-145 | ↓ plasma [46] | ↑ plasma [50] | |||||||
| miR-181 | ↓ PBMC [57] | ↓ serum [67] | |||||||
| miR-205 | ↓ plasma [50] ↓ plasma [49] | ||||||||
| miR-222 | ↑ serum [38] | ↓ serum [67] | |||||||
| miR-328 | ↓ plasma [53] | ↓ plasma [56] | |||||||
| miR-339 | ↑ PBMC [48] | ↓ serum [67] | |||||||
| miR-375 | ↑ plasma [50] ↑ plasma [49] | ||||||||
| miR-411 | ↓ plasma [33] | ↑ serum [47] | |||||||
| miR-769 | ↑ plasma [53] | ↓ PBMC [57] | |||||||
| miR-935 | ↑ PBMC [41] ↑ plasma [43] | ||||||||
| miR-1260 | ↓ plasma [50] | ↓ serum [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeLucas, M.; Sánchez, J.; Palou, A.; Serra, F. The Impact of Diet on miRNA Regulation and Its Implications for Health: A Systematic Review. Nutrients 2024, 16, 770. https://doi.org/10.3390/nu16060770
DeLucas M, Sánchez J, Palou A, Serra F. The Impact of Diet on miRNA Regulation and Its Implications for Health: A Systematic Review. Nutrients. 2024; 16(6):770. https://doi.org/10.3390/nu16060770
Chicago/Turabian StyleDeLucas, María, Juana Sánchez, Andreu Palou, and Francisca Serra. 2024. "The Impact of Diet on miRNA Regulation and Its Implications for Health: A Systematic Review" Nutrients 16, no. 6: 770. https://doi.org/10.3390/nu16060770