
Human Nutrition Research in the Data Era: Results of 11 Reports on the Effects of a Multiple-Micronutrient-Intervention Study

Abstract
:
1. Introduction

2. Rationale and General Experimental Design
2.1. Rationale for the Intervention
2.2. General Statistical Analysis
3. Primary Results [21]
3.1. Population-Level Results
- The studied population in two years (n = 280) had an average age of 11.7 ± 1.1 years old, 55% were female, 43.2% were overweight or obese, and 73.6% were at pubertal status 2 and 3.
- The average total Brazilian healthy-eating index (BHEI-R) score for all included individuals was 54.8 ± 7.5 (53.7 in the first year and 54.5 in the second year). One hundred and fifty-two participants in 2013 and 2014 (91%) had a total BHEI-R below 65, which is considered a “poor diet”, specifically poor in vegetables, legumes, fruits, whole grains, milk and dairy, and rich in sugar and saturated fat; 15 (9%) were classified in the intermediary category, with scores between 65 and 84; and none were in the “good diet” category (above 85) [22]. No statistical differences were found in food-intake patterns across all three visits.
- The percent insufficiencies (in parenthesis) obtained by aggregating data from all participants in both arms of the study were in folate (43%), retinol (24%), α-tocopherol (25.8%), γ-tocopherol (100%), thiamine (99%), vitamin B12 (43%), nicotinamide (8%), pantothenic acid (99%), and pyridoxal (76%).
- A total of 16% of the population were classified as having dyslipidemia.
- The blood level of thymidine monophosphate was positively correlated with percentage of European ancestry, while the levels of vitamin B12 and folate were negatively correlated with percentage of Native American ancestry.
Relevance
3.2. Population-Level Analysis of Clinical and Omic Measures Post-Intervention
Relevance
3.3. Inter-Individual Variability
Relevance
3.4. Predicting Responses to the Intervention
Relevance
4. Secondary Results
4.1. Food-Intake Studies
4.1.1. Healthy-Eating-Index Biomarkers [46]
4.1.2. FFQ Biomarkers
4.1.3. Relevance
4.2. Impact on Lipid Species [55]
Relevance
4.3. Lipoproteins, Polyunsaturated Fatty Acids (PUFAs), 1-Carbon Pathway Metabolites, and B Vitamin Associations
4.3.1. Vitamins and One-Carbon Metabolites [66]
4.3.2. Lipoproteins, PUFAs, and B Vitamin Associations [66]
4.3.3. Relevance
4.4. PUFAs and DNA Damage [68,69]
Relevance
4.5. Metabo Groups
4.5.1. Lipid Profile and Proteomic Metabotypes
4.5.2. Vitamins—Inflammatory Biomarker Metabotypes
4.5.3. Relevance
4.6. Identification of Vitamin B12 Genetic-Risk Score [96]
- LMBRD1 (lysosomal cobalamin transport escort protein), CUBN (cubilin), TCN1 (transcobalamin 1), TCN2 (transcobalamin 2), and ABCD4 (lysosomal cobalamin transporter ABCD4) are involved in the transport of or in the lysosomal release of vitamin B12 into the cytoplasm.
- CUBN, ABCD4 with MTR (exosome RNA helicase) participate in reactions that catalyze the transfer of a methyl group from methyl-cobalamin to homocysteine.
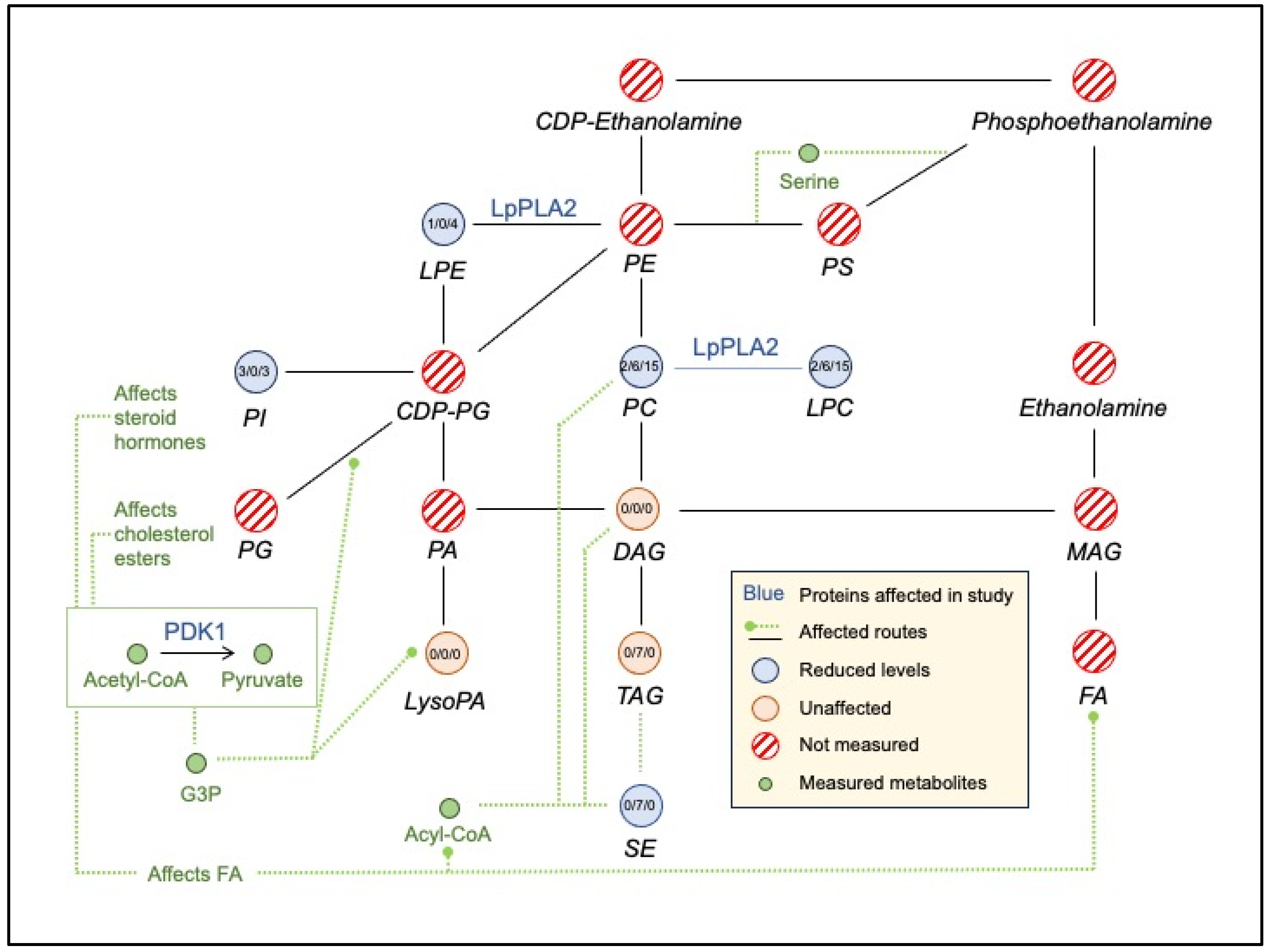
- A small interconnected lipid metabolism network included (i) NDUFAB1 (mitochondrial acyl carrier), an acyl carrier protein of the growing fatty acid chain in fatty acid biosynthesis, (ii) MVK (mevalonate kinase), a regulatory site in the cholesterol biosynthetic pathway, (iii) PEMT (phosphatidylethanolamine N-methyltransferase) which catalyzes the three sequential steps of the methylation pathway involving phosphatidylethanolamine (PE), phosphatidylmonomethylethanolaimne (PMME), phosphatidyldimethylethanolamine (PDME), phosphatidylcholine (PC), and SLC27A4, a fatty-acid transport protein. These interactions are consistent with the changes in lipidemia ([55] and Lipidemia section).
- Vitamin B12 levels may also have a role in the vitamin D metabolic processes through the low-density lipoprotein receptor-related protein 2 (CUBN, LRP2) and in a-amino acid metabolic process (CBS, FPGS, MTR, PEMT, SARS).
Relevance
5. Main Results, Strengths, and Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reider, C.A.; Chung, R.Y.; Devarshi, P.P.; Grant, R.W.; Mitmesser, S.H. Inadequacy of immune health nutrients: Intakes in us adults, the 2005–2016 NHANES. Nutrients 2020, 12, 1735. [Google Scholar] [CrossRef]
- Wunderle, C.; Gomes, F.; Schuetz, P.; Stumpf, F.; Austin, P.; Ballesteros-Pomar, M.D.; Cederholm, T.; Fletcher, J.; Laviano, A.; Norman, K.; et al. ESPEN guideline on nutritional support for polymorbid medical inpatients. Clin. Nutr. 2023, 42, 1545–1568. [Google Scholar] [CrossRef]
- Warsi, O.M.; Dykhuizen, D.E. Evolutionary implications of Liebig’s law of the minimum: Selection under low concentrations of two nonsubstitutable nutrients. Ecol. Evol. 2017, 7, 5296–5309. [Google Scholar] [CrossRef]
- Scott-Boyer, M.P.; Lacroix, S.; Scotti, M.; Morine, M.J.; Kaput, J.; Priami, C. A network analysis of cofactor-protein interactions for analyzing associations between human nutrition and diseases. Sci. Rep. 2016, 6, 19633. [Google Scholar] [CrossRef]
- Kuhn, T.S. The Structure of Scientific Revolutions; 50th Anniversary; University of Chicago Press: Chicago, IL, USA, 1962. [Google Scholar]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized nutrition by prediction of glycemic responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef]
- Berry, S.; Valdes, A.; Davies, R.; Delahanty, L.; Drew, D.; Chan, A.T.; Segata, N.; Franks, P.; Spector, T. Predicting Personal Metabolic Responses to Food Using Multi-omics Machine Learning in over 1000 Twins and Singletons from the UK and US: The PREDICT I Study (OR31-01-19). Curr. Dev. Nutr. 2019, 3, 1324. [Google Scholar] [CrossRef]
- Kaput, J. Lessons from application of data science strategies in nutritional research. Am. J. Clin. Nutr. 2020, 111, 4–5. [Google Scholar] [CrossRef]
- Instituto Brasileiro de Geografia e Estatística. Pesquisa de Orçamentos Familiares 2008–2009: Antropometria e Estado Nutricional de Crianças, Adolescentes e Adultos no Brasil; Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, Brazil, 2010. [Google Scholar]
- Israel, B.A.; Eng, E.; Schulz, A.J.; Parker, E.A. Methods in Community-Based Participatory Research for Health; John Wiley & Sons: Hoboken, NJ, USA, 2005; 479p. [Google Scholar]
- McCabe-Sellers, B.; Lovera, D.; Nuss, H.; Wise, C.; Ning, B.; Teitel, C.; Clark, B.S.; Toennessen, T.; Green, B.; Bogle, M.L.; et al. Personalizing nutrigenomics research through community based participatory research and omics technologies. Omi. A J. Integr. Biol. 2008, 12, 263–272. [Google Scholar] [CrossRef]
- van Ommen, B.; Keijer, J.; Heil, S.G.; Kaput, J. Challenging homeostasis to define biomarkers for nutrition related health. Mol. Nutr. Food Res. 2009, 53, 795–804. [Google Scholar] [CrossRef]
- Stroeve, J.; van Wietmarschen, H.; Kremer, B.; Ben van Ommen, B.; Wopereis, S. Phenotypic Flexibility as a measure of health: The optimal nutritional stress response test. Genes Nutr. 2015, 15, 13. [Google Scholar] [CrossRef]
- Mengersen, K.; McGree, J.M.; Schmid, C.H. 12 Chapter: Statistical Analysis of N-of-1 Trials. In The Essential Guide to N-of-1 Trials in Health; Nikles, J., Mitchell, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 135–154. [Google Scholar]
- Schmid, C.H.; Duan, N. Chapter 4. Statistical Design and Analytic Considerations for N-of-1 Trials. In Design and Implementation of N-of-1 Trials: A User’s Guide; Kravitz, R.L., Duan, N., Eds.; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2014; pp. 33–53. Available online: https://effectivehealthcare.ahrq.gov/products/n-1-trials/research-2014-5 (accessed on 12 November 2023).
- Hendrickson, R.C.; Thomas, R.G.; Schork, N.J.; Raskind, M.A. Optimizing Aggregated N-Of-1 Trial Designs for Predictive Biomarker Validation: Statistical Methods and Theoretical Findings. Front. Digit. Health 2020, 2, 13. [Google Scholar] [CrossRef]
- Daza, E.J. Causal Analysis of Self-tracked Time Series Data Using a Counterfactual Framework for N-of-1 Trials. Methods Inf. Med. 2018, 57, 10–21. [Google Scholar] [CrossRef]
- Zoh, R.S.; Esteves, B.H.; Yu, X.; Fairchild, A.J.; Vazquez, A.I.; Chapple, A.G.; Brown, A.W.; George, B.; Gordon, D.; Landsittel, D.; et al. Design, analysis, and interpretation of treatment response heterogeneity in personalized nutrition and obesity treatment research. Obes. Rev. 2023, 24, e13635. [Google Scholar] [CrossRef]
- Ostermann, T.; Willich, S.N.; Lüdtke, R. Regression toward the mean—A detection method for unknown population mean based on Mee and Chua’s algorithm. BMC Med. Res. Methodol. 2008, 8, 52. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef]
- Mathias, M.G.; Coelho-Landell, C.d.A.; Scott-Boyer, M.P.; Lacroix, S.; Morine, M.J.; Salomão, R.G.; Toffano, R.B.D.; Almada, M.O.R.D.V.; Camarneiro, J.M.; Hillesheim, E.; et al. Clinical and Vitamin Response to a Short-Term Multi-Micronutrient Intervention in Brazilian Children and Teens: From Population Data to Interindividual Responses. Mol. Nutr. Food Res. 2018, 62, 1700613. [Google Scholar] [CrossRef]
- Hann, C.S.; Rock, C.L.; King, I.; Drewnowski, A. Validation of the healthy eating index with use of plasma biomarkers in a clinical sample of women. Am. J. Clin. Nutr. 2001, 74, 479–486. [Google Scholar] [CrossRef]
- Haider, B.A.; Bhutta, Z.A. Multiple-micronutrient supplementation for women during pregnancy. Cochrane Database Syst. Rev. 2017, 11, 14. [Google Scholar] [CrossRef]
- Earnest, C.; Cooper, K.H.; Marks, A.; Mitchell, T.L. Efficacy of a complex multivitamin supplement. Nutrition 2002, 18, 738–742. [Google Scholar] [CrossRef]
- Ashraf, A.P.; Alvarez, J.A.; Gower, B.A.; Saenz, K.H.; McCormick, K.L. Associations of serum 25-hydroxyvitamin D and components of the metabolic syndrome in obese adolescent females. Obesity 2011, 19, 2214–2221. [Google Scholar] [CrossRef]
- Kelishadi, R.; Farajzadegan, Z.; Bahreynian, M. Association between vitamin D status and lipid profile in children and adolescents: A systematic review and meta-analysis. Int. J. Food Sci. Nutr. 2014, 65, 404–410. [Google Scholar] [CrossRef]
- Birjmohun, R.S.; Hutten, B.A.; Kastelein, J.J.P.; Stroes, E.S.G. Efficacy and safety of high-density lipoprotein cholesterol-increasing compounds. J. Am. Coll. Cardiol. 2005, 45, 185–197. [Google Scholar] [CrossRef]
- Partida-Hernández, G.; Arreola, F.; Fenton, B.; Cabeza, M.; Román-Ramos, R.; Revilla-Monsalve, M.C. Effect of zinc replacement on lipids and lipoproteins in type 2-diabetic patients. Biomed. Pharmacother. 2006, 60, 161–168. [Google Scholar] [CrossRef]
- Barbagallo, M.; Dominguez, L.J.; Galioto, A.; Ferlisi, A.; Cani, C.; Malfa, L.; Pineo, A.; Busardo’, A.; Paolisso, G. Role of magnesium in insulin action, diabetes and cardio-metabolic syndrome X. Mol. Asp. Med. 2003, 24, 39–52. [Google Scholar] [CrossRef]
- Heng, E.C.; Karsani, S.A.; Abdul Rahman, M.; Abdul Hamid, N.A.; Hamid, Z.; Wan Ngah, W.Z. Supplementation with tocotrienol-rich fraction alters the plasma levels of Apolipoprotein A-I precursor, Apolipoprotein E precursor, and C-reactive protein precursor from young and old individuals. Eur. J. Nutr. 2013, 52, 1811–1820. [Google Scholar] [CrossRef]
- Burdeos, G.C.; Nakagawa, K.; Kimura, F.; Miyazawa, T. Tocotrienol Attenuates Triglyceride Accumulation in HepG2 Cells and F344 Rats. Lipids 2012, 47, 471–481. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Sami, S.A.; Salser, W.A.; Khan, F.A. Synergistic effect of tocotrienol-rich fraction (TRF(25)) of rice bran and lovastatin on lipid parameters in hypercholesterolemic humans. J. Nutr. Biochem. 2001, 12, 318–329. [Google Scholar] [CrossRef]
- Wu, S.-J.; Liu, P.-L.; Ng, L.-T. Tocotrienol-rich fraction of palm oil exhibits anti-inflammatory property by suppressing the expression of inflammatory mediators in human monocytic cells. Mol. Nutr. Food Res. 2008, 52, 921–929. [Google Scholar] [CrossRef]
- Al-Daghri, N.; Al-Attas, O.; Al-Daghri, N.; Alokail, M.; Abd-Alrahman, S.; Vinodson, B.; Sabico, S. Metabolic Benefits of Six-month Thiamine Supplementation in Patients with and without Diabetes Mellitus Type 2. Clin. Med. Insights Endocrinol. Diabetes 2014, 7. [Google Scholar] [CrossRef]
- Doshi, S.N.; McDowell, I.F.W.; Moat, S.J.; Payne, N.; Durrant, H.J.; Lewis, M.J.; Goodfellow, J. Folic acid improves endothelial function in coronary artery disease via mechanisms largely independent of homocysteine lowering. Circulation 2002, 105, 22–26. [Google Scholar] [CrossRef]
- O’Hanesian, M.A.; Rosner, B.; Bishop, L.M.; Sacks, F.M. Effects of inherent responsiveness to diet and day-to-day diet variation on plasma lipoprotein concentrations. Am. J. Clin. Nutr. 1996, 64, 53–59. [Google Scholar] [CrossRef]
- Williams, R.P. Biochemical Individuality: The Basis for the Genetotrophic Concept; Keats Publishing: New Canaan, CT, USA, 1956. [Google Scholar]
- Rundblad, A.; Sandoval, V.; Holven, K.B.; Ordovás, J.M.; Ulven, S.M. Omega-3 fatty acids and individual variability in plasma triglyceride response: A mini-review. Redox Biol. 2023, 63, 102730. [Google Scholar] [CrossRef]
- Abdullah, M.M.H.; Vazquez-Vidal, I.; Baer, D.J.; House, J.D.; Jones, P.J.H.; Desmarchelier, C. Common genetic variations involved in the inter-individual variability of circulating cholesterol concentrations in response to diets: A narrative review of recent evidence. Nutrients 2021, 13, 695. [Google Scholar] [CrossRef]
- Kaput, J. Developing the pathway to personalized health: The potential of N-of-1 studies for personalizing nutrition. J. Nutr. 2021, 151, 2863–2864. [Google Scholar] [CrossRef]
- Hillesheim, E.; Yin, X.; Sundaramoorthy, G.P.; Brennan, L. Using a Metabotype Framework to Deliver Personalized Nutrition Improves Dietary Quality and Metabolic Health Parameters: A 12-Week Randomized Controlled Trial. Mol. Nutr. Food Res. 2023, 67, 2200620. [Google Scholar] [CrossRef]
- Hughes, R.L.; Kable, M.E.; Marco, M.; Keim, N.L. The Role of the Gut Microbiome in Predicting Response to Diet and the Development of Precision Nutrition Models. Part II: Results. Adv. Nutr. 2019, 10, 979–998. [Google Scholar] [CrossRef]
- Hughes, R.L.; Marco, M.L.; Hughes, J.P.; Keim, N.L.; Kable, M.E. The Role of the Gut Microbiome in Predicting Response to Diet and the Development of Precision Nutrition Models—Part I: Overview of Current Methods. Adv. Nutr. 2019, 10, 953–978. [Google Scholar] [CrossRef]
- Boeing, H. Nutritional epidemiology: New perspectives for understanding the diet-disease relationship ? Eur. J. Clin. Nutr. 2013, 67, 424–429. [Google Scholar] [CrossRef]
- O’Gorman, A.; Brennan, L. The role of metabolomics in determination of new dietary biomarkers. Proc. Nutr. Soc. 2017, 76, 295–302. [Google Scholar] [CrossRef]
- Toffano, R.B.D.; Hillesheim, E.; Mathias, M.G.M.G.; Coelho-Landell, C.A.; Salomão, R.G.; Almada, M.O.R.V.; Camarneiro, J.M.; Barros, T.T.T.; Camelo-Junior, J.S.; Rezzi, S.; et al. Validation of the brazilian healthy eating index-revised using biomarkers in children and adolescents. Nutrients 2018, 10, 154. [Google Scholar] [CrossRef]
- Hillesheim, E.; Toffano, R.B.D.; de Barros, T.T.; Salomão, R.G.; Mathias, M.G.; Coelho-Landell, C.d.A.; Almada, M.O.R.d.V.; Camarneiro, J.M.; Camelo-Junior, J.S.; Ued, F.d.V.; et al. Biomarker-based validity of a food frequency questionnaire estimating intake in Brazilian children and adolescents. Int. J. Food Sci. Nutr. 2020, 72, 236–247. [Google Scholar] [CrossRef]
- Neufingerl, N.; Eilander, A. Nutrient Intake and Status in Adults Consuming Plant-Based Diets Compared to Meat-Eaters: A Systematic Review. Nutrients 2022, 14, 29. [Google Scholar] [CrossRef]
- Neufingerl, N.; Eilander, A. Nutrient Intake and Status in Children and Adolescents Consuming Plant-Based Diets Compared to Meat-Eaters: A Systematic Review. Nutrients 2023, 15, 4341. [Google Scholar] [CrossRef]
- Cuparencu, C.; Praticó, G.; Hemeryck, L.Y.; Sri Harsha, P.S.C.; Noerman, S.; Rombouts, C.; Xi, M.; Vanhaecke, L.; Hanhineva, K.; Brennan, L.; et al. Biomarkers of meat and seafood intake: An extensive literature review. Genes Nutr. 2019, 14, 35. [Google Scholar] [CrossRef]
- Li, K.J.; Brouwer-brolsma, E.M.; Burton-pimentel, K.J.; Vergères, G.; Feskens, E.J.M. A systematic review to identify biomarkers of intake for fermented food products. Genes Nutr. 2021, 16, 5. [Google Scholar] [CrossRef]
- Vázquez-Fresno, R.; Rosana, A.R.R.; Sajed, T.; Onookome-Okome, T.; Wishart, N.A.; Wishart, D.S. Herbs and Spices- Biomarkers of Intake Based on Human Intervention Studies—A Systematic Review. Genes Nutr. 2019, 14, 18. [Google Scholar] [CrossRef]
- Landberg, R.; Hanhineva, K.; Tuohy, K.; Garcia-Aloy, M.; Biskup, I.; Llorach, R.; Yin, X.; Brennan, L.; Kolehmainen, M. Biomarkers of cereal food intake. Genes Nutr. 2019, 14, 28. [Google Scholar] [CrossRef]
- Praticò, G.; Gao, Q.; Manach, C.; Dragsted, L.O. Biomarkers of food intake for Allium vegetables. Genes Nutr. 2018, 13, 34. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Eiden, M.; Morin-Rivron, D.; Christinat, N.; Monteiro, J.P.; Kaput, J.; Masoodi, M. Impact of multi-micronutrient supplementation on lipidemia of children and adolescents. Clin. Nutr. 2020, 39, 2211–2219. [Google Scholar] [CrossRef]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef]
- Sauvant, P.; Cansell, M.; Atgie, C. Vitamin A and lipid metabolism: Relationship between hepatic stellate cells (HSCs) and adipocytes. J. Physiol. Biocehm. 2011, 67, 4870496. [Google Scholar] [CrossRef]
- Adaikalakoteswari, A.; Jayashri, R.; Sukumar, N.; Venkataraman, H.; Pradeepa, R.; Gokulakrishnan, K.; Anjana, R.M.; Mcternan, P.G.; Tripathi, G.; Patel, V.; et al. Vitamin B12 deficiency is associated with adverse lipid profile in Europeans and Indians with type 2 diabetes. Cardiovasc. Diabetol. 2014, 13, 129. [Google Scholar] [CrossRef]
- Sakuragi, T. The function of vitamin B6 in fat metabolism. J. Am. Oil Chem. Soc. 1959, 36, 251–253. [Google Scholar] [CrossRef]
- Desmarchelier, C.; Borel, P.; Lairon, D.; Maraninchi, M.; Valero, R. Effect of Nutrient and Micronutrient Intake on Chylomicron Production and Postprandial Lipemia. Nutrients 2019, 11, 1299. [Google Scholar] [CrossRef]
- Da Silva, L.; Collino, S.; Cominetti, O.; Martin, F.-P.; Montoliu, I.; Moreno, S.O.; Corthesy, J.; Kaput, J.; Kussmann1, M.; Monteiro, J.P.; et al. High-throughput method for the quantitation of metabolites and co-factors from homocysteine—Methionine cycle for nutritional status assessment. Bioanalysis 2016, 8, 1937–1949. [Google Scholar] [CrossRef]
- Finkelstein, J.D. Metabolic regulatory properties of S-adenosylmethionine and S-adenosylhomocysteine. Clin. Chem. Lab. Med. 2007, 45, 1694–1699. [Google Scholar] [CrossRef]
- Lennard, L. Methyltransferases. Compr. Toxicol. 2010, 4, 453–457. [Google Scholar]
- Monteiro, J.P.; Wise, C.; Morine, M.J.M.; Teitel, C.; Pence, L.; Williams, A.; McCabe-Sellers, B.; Champagne, C.; Turner, J.; Shelby, B.; et al. Methylation Potential Associated with Diet, Genotype, Protein, and Metabolite Levels in the Delta Obesity Vitamin Study. Genes Nutr. 2014, 9, 403. [Google Scholar] [CrossRef]
- Morine, M.J.M.J.; Monteiro, J.P.J.P.; Wise, C.; Teitel, C.; Pence, L.; Williams, A.; Ning, B.; McCabe-Sellers, B.; Champagne, C.; Turner, J.; et al. Genetic associations with micronutrient levels identified in immune and gastrointestinal networks. Genes Nutr. 2014, 9, 408. [Google Scholar] [CrossRef]
- Ued, F.V.; Mathias, M.G.; Toffano, R.B.D.; Barros, T.T.; Almada, M.O.R.V.; Salomão, R.G.; Coelho-Landell, C.A.; Hillesheim, E.; Camarneiro, J.M.; Camelo-Junior, J.S.; et al. Vitamin B2 and folate concentrations are associated with ARA, EPA and DHA fatty acids in red blood cells of Brazilian children and adolescents. Nutrients 2019, 11, 2918. [Google Scholar] [CrossRef]
- Zhu, J.; Xun, P.C.; Kolencik, M.; Yang, K.F.; Fly, A.D.; Kahe, K. Do B Vitamins Enhance the Effect of Omega-3 Polyunsaturated Fatty Acids on Cardiovascular Diseases? A Systematic Review of Clinical Trials. Nutrients 2022, 14, 1608. [Google Scholar] [CrossRef]
- De Barros, T.T.; Venâncio, V.D.P.; Hernandes, L.C.; Greggi Antunes, L.M.; Hillesheim, E.; Salomão, R.G.; Mathias, M.G.; Coelho-Landell, C.A.; Toffano, R.B.D.; Do Vale Almada, M.O.R.; et al. DNA damage is inversely associated to blood levels of DHA and EPA fatty acids in Brazilian children and adolescents. Food Funct. 2020, 11, 5115–5121. [Google Scholar] [CrossRef]
- de Barros, T.T.; Venancio, V.d.P.; Hernandes, L.C.; Antunes, L.M.G.; Hillesheim, E.; Salomão, R.G.; Mathias, M.G.; Coelho-Landell, C.A.; Toffano, R.B.D.; Almada, M.O.R.D.V.; et al. Dna damage, n-3 long-chain pufa levels and proteomic profile in brazilian children and adolescents. Nutrients 2021, 13, 2483. [Google Scholar] [CrossRef]
- Burlinson, B.; Tice, R.R.; Speit, G.; Agurell, E.; Brendler-Schwaab, S.Y.; Collins, A.R.; Escobar, P.; Honma, M.; Kumaravel, T.S.; Nakajima, M.; et al. Fourth International Workgroup on Genotoxicity testing: Results of the in vivo Comet assay workgroup. Mutat. Res. 2007, 627, 31–35. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Müllner, E.; Brath, H.; Pleifer, S.; Schiermayr, C.; Baierl, A.; Wallner, M.; Fastian, T.; Millner, Y.; Paller, K.; Henriksen, T.; et al. Vegetables and PUFA-rich plant oil reduce DNA strand breaks in individuals with type 2 diabetes. Mol. Nutr. Food Res. 2013, 57, 328–338. [Google Scholar] [CrossRef]
- Ames, B.N. DNA damage from micronutrient deficiencies is likely to be a major cause of cancer. Mutat. Res. 2001, 475, 7–20. [Google Scholar] [CrossRef]
- Bull, C.; Fenech, M. Genome-health nutrigenomics and nutrigenetics: Nutritional requirements or “nutriomes” for chromosomal stability and telomere maintenance at the individual level. Proc. Nutr. Soc. 2008, 67, 146–156. [Google Scholar] [CrossRef]
- Reszka, E.; Wasowicz, W.; Gromadzinska, J. Genetic polymorphism of xenobiotic metabolising enzymes, diet and cancer susceptibility. Br. J. Nutr. 2006, 96, 609–619. [Google Scholar]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat Inflamm Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- Ames, B.N. The Metabolic Tune-up: Metabolic Harmony and Disease Prevention. J. Nutr. 2003, 133, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.P.J.P.; Kussmann, M.; Kaput, J. The genomics of micronutrient requirements. Genes Nutr. 2015, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Hillesheim, E.; Brennan, L. Metabotyping and its role in nutrition research. Nutr. Res. Rev. 2020, 33, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Celis-Morales, C.; Livingstone, K.M.; Marsaux, C.F.M.; Macready, A.L.; Fallaize, R.; O’Donovan, C.B.; Woolhead, C.; Forster, H.; Walsh, M.C.; Navas-Carretero, S.; et al. Effect of personalized nutrition on health-related behaviour change: Evidence from the Food4Me European randomized controlled trial. Int. J. Epidemiol. 2017, 46, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Landell, C.A.; Salomão, R.G.; Almada, M.O.R.d.V.; Mathias, M.G.; Toffano, R.B.D.; Hillesheim, E.; Barros, T.T.; Camarneiro, J.M.; Camelo-Junior, J.S.; Rosa, J.C.; et al. Metabo groups in response to micronutrient intervention: Pilot study. Food Sci. Nutr. 2020, 8, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Almada, M.O.R.D.V.; Almeida, A.C.F.; Ued, F.d.V.; Mathias, M.G.; Coelho-Landell, C.d.A.; Salomão, R.G.; Toffano, R.B.D.; Camarneiro, J.M.; Hillesheim, E.; de Barros, T.T.; et al. Metabolic groups related to blood vitamin levels and inflammatory biomarkers in Brazilian children and adolescents. J. Nutr. Sci. Vitaminol. 2020, 66, 515–525. [Google Scholar] [CrossRef]
- Carter, K.; Worwood, M. Haptoglobin: A review of the major allele frequenciesworldwide and their association with diseases. Int. Lab. Hem. 2007, 29, 92–110. [Google Scholar] [CrossRef] [PubMed]
- Toonen, E.J.; Mirea, A.M.; Tack, C.J.; Stienstra, R.; Ballak, D.B.; Van Diepen, J.A.; Hijmans, A.; Chavakis, T.; Dokter, W.H.; Pham, C.T.; et al. Activation of proteinase 3 contributes to nonalcoholic fatty liver disease and insulin resistance. Mol. Med. 2016, 22, 202–214. [Google Scholar] [CrossRef]
- Davis, A.E.I.; Meja, P.; Lu, F. Biological Activities of C1 Inhibitor. Mol. Immunol. 2008, 45, 4057–4063. [Google Scholar] [CrossRef]
- Luo, Z.; Lei, H.; Sun, Y.; Liu, X.; Su, D.F. Orosomucoid, an acute response protein with multiple modulating activities. J. Physiol. Biochem. 2015, 71, 329–340. [Google Scholar] [CrossRef]
- Tesseromatis, C.; Alevizou, A.; Tigka, E.; Kotsiou, A. Acute Phase Proteins—Regulation and Functions of Acute Phase Proteins; Veas, F., Ed.; InTech: Rijeka, Croatia, 2011; pp. 247–260. [Google Scholar]
- Gruys, E.; Toussaint, M.J.M.; Niewold, T.A.; Koopmans, S.J. Acute phase reaction and acute phase proteins. J. Zhejiang Univ. Sci. 2005, 6, 1045–1056. [Google Scholar] [CrossRef]
- Perl, M.L.; Finkelstein, A.; Revivo, M.; Berliner, S.; Herz, I.; Rabinovich, I.; Ziv-Baran, T.; Gotler, D.; Keren, G.; Bana, S.; et al. Variance in biomarker usefulness as indicators for carotid and coronary atherosclerosis. Isr. Med. Assoc. J. 2016, 18, 80–84. [Google Scholar]
- Poredoš, P.; Ježovnik, M.K. Markers of preclinical atherosclerosis and their clinical relevance. Vasa—Eur. J. Vasc. Med. 2015, 44, 247–256. [Google Scholar] [CrossRef]
- Aleman, M.M.; Walton, B.L.; Byrnes, J.R.; Wolberg, A.S. Fibrinogen and red blood cells in venous thrombosis. Thromb Res. 2014, 133, S38–S40. [Google Scholar] [CrossRef]
- Ferrari, M.; Cuenca-García, M.; Valtueña, J.; Moreno, L.A.; Censi, L.; González-Gross, M.; Androutsos, O.; Gilbert, C.C.; Huybrechts, I.; Dallongeville, J.; et al. Inflammation profile in overweight/obese adolescents in Europe: An analysis in relation to iron status. Eur. J. Clin. Nutr. 2015, 69, 247–255. [Google Scholar] [CrossRef]
- Peng, H.Y.; Man, C.F.; Xu, J.; Fan, Y. Elevated homocysteine levels and risk of cardiovascular and all-cause mortality: A meta-analysis of prospective studies. J. Zhejiang Univ. Sci. B 2015, 16, 78–86. [Google Scholar] [CrossRef]
- Funk, C.D. Leukotriene modifiers as potential therapeutics for cardiovascular disease. Nat. Rev. Drug Discov. 2005, 4, 664–672. [Google Scholar] [CrossRef]
- Fiamoncini, J.; Rundle, M.; Gibbons, H.; Thomas, E.L.; Geillinger-Kästle, K.; Bunzel, D.; Trezzi, J.-P.; Kiselova-Kaneva, Y.; Wopereis, S.; Wahrheit, J.; et al. Plasma metabolome analysis identifies distinct human metabotypes in the postprandial state with different susceptibility to weight loss–mediated metabolic improvements. FASEB J. 2018, 32, 5447–5458. [Google Scholar] [CrossRef]
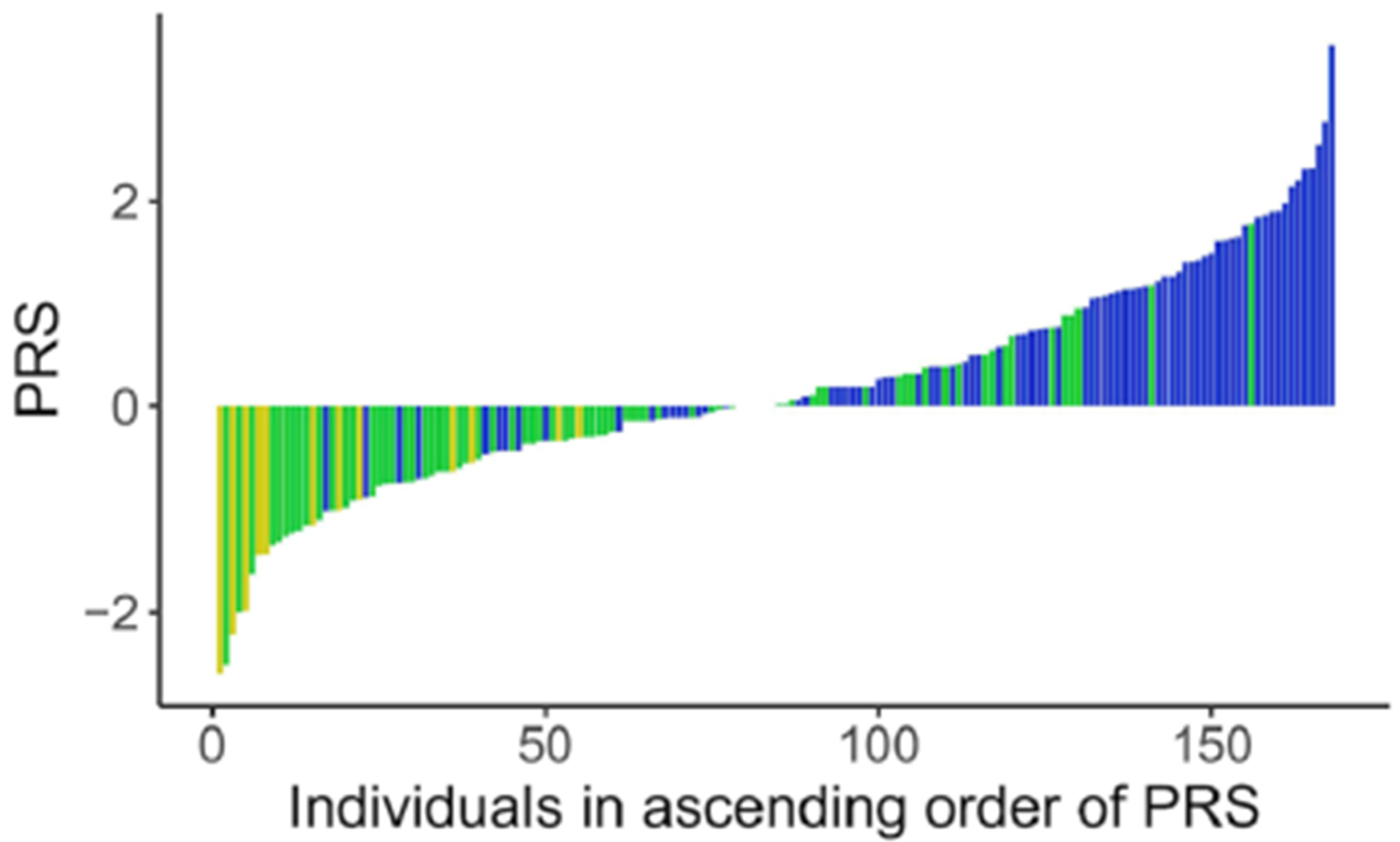
- Fuzo, C.A.; da Veiga Ued, F.; Moco, S.; Cominetti, O.; Métairon, S.; Pruvost, S.; Charpagne, A.; Carayol, J.; Torrieri, R.; Silva, W.A.; et al. Contribution of genetic ancestry and polygenic risk score in meeting vitamin B12 needs in healthy Brazilian children and adolescents. Sci. Rep. 2021, 11, 11992. [Google Scholar] [CrossRef]
- Hellwege, J.N.; Keaton, J.M.; Giri, A.; Gao, X.; Velez Edwards, D.R.; Edwards, T.L. Population Stratification in Genetic Association Studies. Curr. Protoc. Hum. Genet. 2017, 95, 1.22.1–1.22.23. [Google Scholar] [CrossRef]
- Guan, Y. Detecting structure of haplotypes and local ancestry. Genetics 2014, 196, 625–642. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Torkamani, A.; Wineinger, N.E.; Topol, E.J. The personal and clinical utility of polygenic risk scores. Nat. Rev. Genet. 2018, 19, 581–590. [Google Scholar] [CrossRef]
- Grimaldi, K.A.; van Ommen, B.; Ordovas, J.M.; Parnell, L.D.; Mathers, J.C.; Bendik, I.; Brennan, L.; Celis-Morales, C.; Cirillo, E.; Daniel, H.; et al. Proposed guidelines to evaluate scientific validity and evidence for genotype-based dietary advice. Genes Nutr. 2017, 12, 35. [Google Scholar] [CrossRef]
- Tishkoff, S.A.; Reed, F.A.; Ranciaro, A.; Voight, B.F.; Babbitt, C.C.; Silverman, J.S.; Powell, K.; Mortensen, H.M.; Hirbo, J.B.; Osman, M.; et al. Convergent adaptation of human lactase persistence in Africa and Europe. Nat. Genet 2007, 39, 31–40. [Google Scholar] [CrossRef]
- Kaput, J.; Perozzi, G.; Radonjic, M.; Virgili, F. Propelling the paradigm shift from reductionism to systems nutrition. Genes Nutr. 2017, 12, 3. [Google Scholar] [CrossRef]
- Schork, N.J.; Goetz, L.H. Single subject (N-of-1) Studies in Translational Nutrition Research. Annu. Rev. Nutr. 2017, 37, 395–422. [Google Scholar] [CrossRef]
- Kirk, D.; Catal, C.; Tekinerdogan, B. Precision nutrition: A systematic literature review. Comput. Biol. Med. 2021, 133, 104365. [Google Scholar] [CrossRef]
- Horowitz, C.R.; Robinson, M.; Seifer, S. Community-based participatory research from the margin to the mainstream: Are researchers prepared? Circulation 2009, 119, 2633–2642. [Google Scholar] [CrossRef]
- Bogart, L.M.; Uyeda, K. Community-based participatory research: Partnering with communities for effective and sustainable behavioral health interventions. Health Psychol. 2009, 28, 391–393. [Google Scholar] [CrossRef]
- Kaput, J.; Ordovas, J.M.; Ferguson, L.; van Ommen, B.; Rodriguez, R.L.; Allen, L.; Ames, B.N.; Dawson, K.; German, B.; Krauss, R.; et al. The case for strategic international alliances to harness nutritional genomics for public and personal health. Br. J. Nutr. 2005, 94, 623–632. [Google Scholar] [CrossRef]
- Kaput, J.; van Ommen, B.; Kremer, B.; Priami, C.; Monteiro, J.P.; Morine, M.; Pepping, F.; Diaz, Z.; Fenech, M.; He, Y.; et al. Consensus statement—Understanding health and malnutrition through a systems approach: The ENOUGH program for early life. Genes Nutr. 2014, 9, 378. [Google Scholar] [CrossRef]



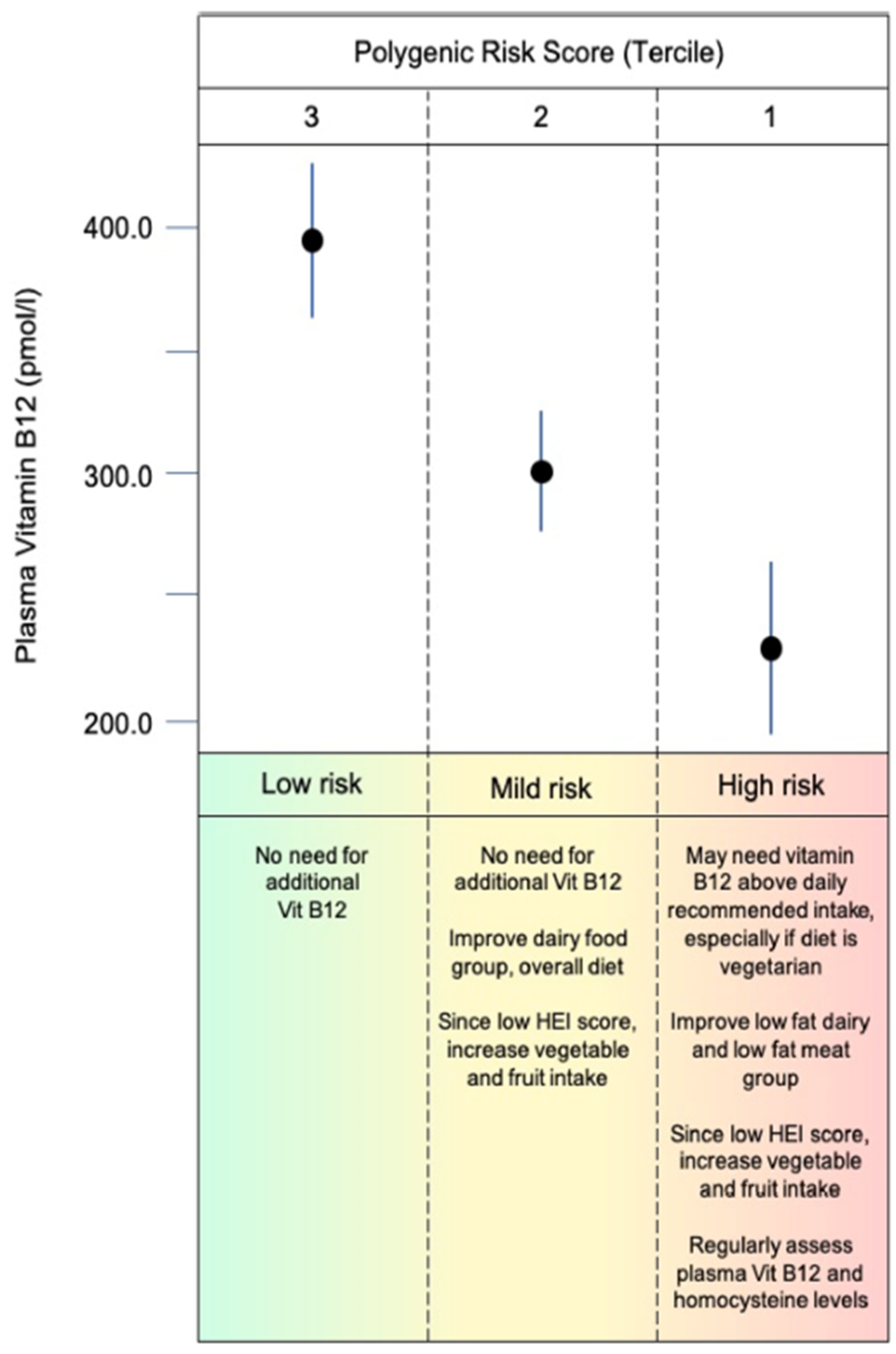
 , normal
, normal  and high
and high  .
, normal and high .
.
, normal and high .


{kind=link}
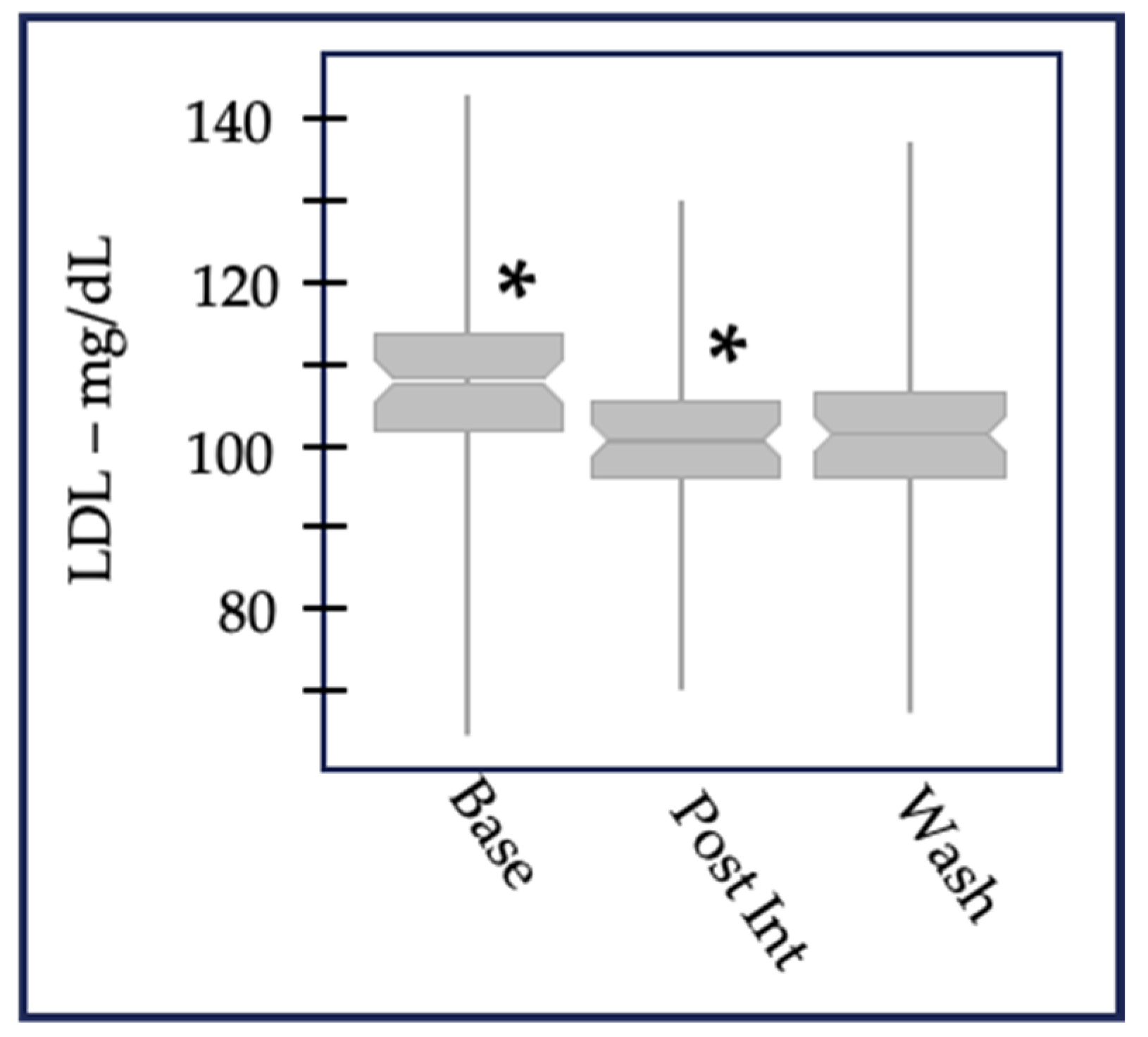{kind=link}
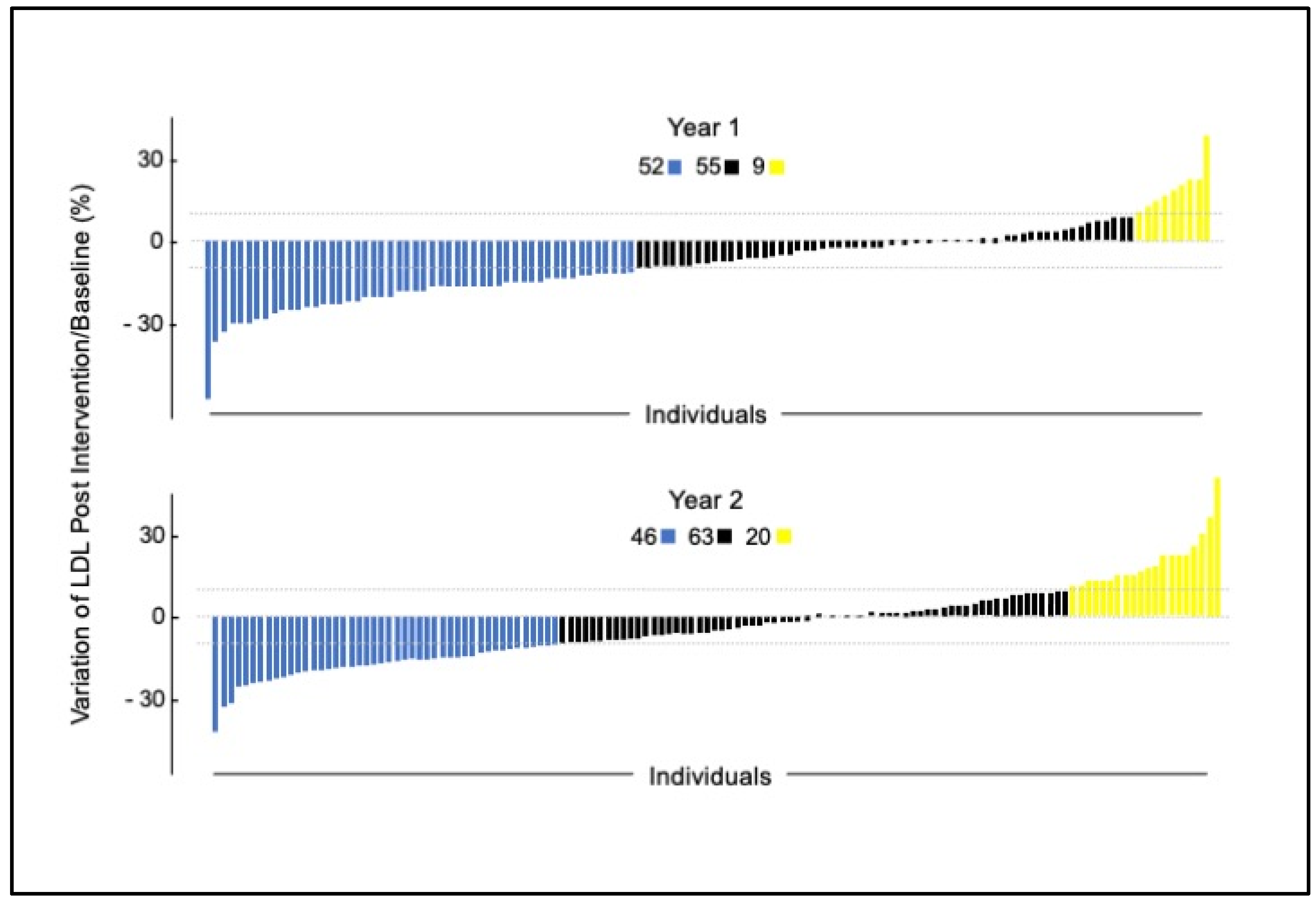{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Number of Variables |
|---|---|
| Anthropometry | 7 |
| Body Mass Index (BMI) | 1 |
| 24 h recall | 28 |
| Food Frequency Questionnaire | 28 |
| Physical Activity | 20 |
| Social Economic Status | 22 |
| Clinical Biochemistry | 18 |
| Hematology | 13 |
| Tanner Classification | 1 |
| Proteomics (plasma-Somalogic) | 1129 |
| Proteomics iTRAQ | 20 |
| Metabolomics (plasma-NMR) | 24 |
| Lipidomics | 76 |
| Fatty acids (Red Blood Cells) | 25 |
| Amino acids (Red Blood Cells) | 25 |
| Comet Assay (whole blood) | 9 |
| Vitamins (plasma) | 36 |
| Methionine Pathway (RBC) | 13 |
| Healthy Eating Index (HEI) | 13 |
| Whole genome genotyping | >4.3 million |
| Exome (percent of genome) | ~2% |
| GIS Food Access & Quality | ~50 |
| Food price comparison | 16 |
| Dietary Group | Metabolite | R | p Value |
|---|---|---|---|
| Fruit intake | Linoleic acid | 0.23 | 0.003 |
| α-linolenic acid | 0.3 | 0.001 | |
| EPA | 0.26 | 0.001 | |
| DHA | 0.29 | 0.001 | |
| β-carotene | 0.19 | 0.020 | |
| Legumes/Vegetables | Linoleic acid | 0.25 | 0.002 |
| α-linolenic acid | 0.36 | 0.001 | |
| EPA | 0.27 | 0.001 | |
| DHA | 0.34 | 0.001 | |
| β-carotene | 0.25 | 0.002 | |
| All vegetables (including legumes) | Creatine | 0.31 | 0.003 |
| Dark greens and legumes | Creatine | 0.37 | 0.001 |
| Animal protein | Creatine | 0.34 | 0.003 |
| Milk/Dairy | Retinol | 0.19 | 0.001 |
| Pyridoxal | 0.21 | 0.007 |
| Nutrient Intake | Metabolite | R | p Value 1 |
|---|---|---|---|
| Animal protein | Creatine | 0.19 | <0.05 |
| Myristic acid (C14:0) | C14:0 | 0.2 | <0.01 |
| EPA | EPA | 0.15 | <0.05 |
| DHA | DHA | 0.18 | <0.05 |
| β-carotene | β-carotene | 0.31 | <0.001 |
| Folate | Folate | 0.15 | <0.05 |
| Vitamin B3 | Nudifloramide | 0.17 | <0.05 |
| Vitamin B5 | Pantothenic acid | 0.17 | <0.05 |
| Vitamin B6 | Pyridoxal 5′-phosphate | 0.19 | <0.05 |
| Food Groups and Biomarkers | |||
| Fish products | EPA | 0.19 | <0.01 |
| DHA | 0.15 | <0.01 | |
| Milk/Dairy | Myristic acid (C14:0) | 0.20 | <0.01 |
| Pyridoxal 5′-phosphate | 0.32 | <0.001 | |
| Vitamin B12 | 0.23 | <0.001 | |
| Total vegetables | β-carotene | 0.36 | <0.05 |
| Dark green/orange | β-carotene | 0.36 | <0.05 |
| Green vegetables | 5-methyltetrahydrofolate | 0.20 | <0.05 |
| Flour products | Para-aminobenoylglutamic acid | 0.27 | <0.01 |
| Metabolite 1 | Metabolite 2 | Δ Metabolite 1 1 | Δ Metabolite 2 2 |
|---|---|---|---|
| Vitamin B2 | S-adenosylmethionine (SAM) | −1 nmol/L | −1.8 µmol/L |
| Vitamin B2 | SAM:SAH ratio | −1 nmol/L | −0.20 |
| Vitamin B6 | Homocysteine (Hcy) | +1 nmol/L | −0.11 µmol/L |
| Vitamin B12 | Homocysteine (Hcy) | +1 nmol/L | −0.14 µmol/L |
| Hcy | Linoleic acid (LA) | +1 nmol/L | +0.24 mg/dL |
| α-linolenic acid (ALA) | +0.24 mg/dL | ||
| Arachidonic acid (ARA) | +0.38 mg/dL | ||
| Eicosapentaenoic acid (EPA) | +0.35 mg/dL | ||
| Docosahexenoic Acid (DHA) | +0.49 mg/dL |
| Vitamin | Fatty Acid | Δ B2 1 | Δ Fatty Acid 2 |
|---|---|---|---|
| Vitamin B2 | Linoleic Acid (LA) | +1 nmol/L | +0.15 mg/dL |
| α-linolenic acid (ALA) | +0.15 mg/dL | ||
| Eicosapentaenoic acid (EPA) | +0.19 mg/dL | ||
| Arachidonic acid (ARA) | +0.20 mg/dL | ||
| Docosahexenoic Acid (DHA) | +0.25 mg/dL | ||
| Folate | Linoleic Acid (LA) | +1 ng/mL | +0.15 mg/dL |
| α-linolenic acid (ALA) | +0.15 mg/dL | ||
| Eicosapentaenoic acid (EPA) | +0.14 mg/dL | ||
| Arachidonic acid (ARA) | +0.22 mg/dL | ||
| Docosahexenoic Acid (DHA) | +0.21 mg/dL |
| Vitamin | Fatty Acid | Association | ß-Coefficient | p Value |
|---|---|---|---|---|
| B2 | Palmitoleic | Positive | 0.12 | <0.01 |
| Oleic | Positive | 0.06 | <0.01 | |
| Elaidic trans-Fatty Acid | Negative | 0.10 | <0.01 | |
| B12 | Palmitoleic | Positive | 0.13 | <0.01 |
| Oleic | Positive | 0.06 | <0.01 | |
| Palmitic | Positive | 0.04 | <0.01 | |
| Stearic | Positive | 0.05 | <0.01 | |
| Eicosanoic | Positive | 0.05 | <0.01 | |
| B6 | Linoleic Acid | Negative | 0.07 | 0.02 |
| Alpha linoleic Acid | Negative | 0.10 | <0.01 | |
| Arachidonic acid | Negative | 0.1 | 0.02 | |
| Docosahexenoic acid | Negative | 0.12 | 0.03 |
| Group 1 n = 10 | Group 2 n = 10 | p Value | |
|---|---|---|---|
| Mean Triglycerides mg/dL | 63.3 (39–103.7) | 133.7 (75–220.3) | 0.001 |
| Mean LDL mg/dL | 85.5 (66.7–124) | 114.5 (48–152.3) | 0.143 |
| Mean VLDL mg/dL | 12.7 (8.0–21) | 26.7 (15.7–44) | 0.001 |
| α-tocopherol (Vit E) V2 (µg/mL) | 5.6 (3.1–7.5) | 8.0 (3.1–10.1) | 0.063 |
| Genetic Ancestry | |||
| African (%) | 16.8 (4.8–58.6) | 35.1 (10.4–96.8) | 0. 06 |
| Europe (%) | 71.3 (17–89.9) | 29.1 (0–69) | 0.004 |
| Native America (%) | 7.1 (0–23.7) | 14 (3.1–43.8) | 0.031 |
| Group 1 n = 30 | Group 2 n = 94 | p Value | |
|---|---|---|---|
| Riboflavin nmol/L | 18.1 ± 10.4 | 13.1 ± 8.3 | 0.01 |
| Pyridoxal nmol/L | 8.9 ± 2.9 | 8.0 ± 3.0 | 0.04 |
| Cobalamin pg/mL | 783 ± 210 | 384 ± 192 | 0.01 |
| Linoleic acid mg/dL | 15.3 ± 4.7 | 16.6 ± 4.2 | 0.04 |
| Homocysteine umol/L | 2.5 ± 0.06 | 2.9 ± 0.8 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaput, J.; Monteiro, J.P. Human Nutrition Research in the Data Era: Results of 11 Reports on the Effects of a Multiple-Micronutrient-Intervention Study. Nutrients 2024, 16, 188. https://doi.org/10.3390/nu16020188
Kaput J, Monteiro JP. Human Nutrition Research in the Data Era: Results of 11 Reports on the Effects of a Multiple-Micronutrient-Intervention Study. Nutrients. 2024; 16(2):188. https://doi.org/10.3390/nu16020188
Chicago/Turabian StyleKaput, Jim, and Jacqueline Pontes Monteiro. 2024. "Human Nutrition Research in the Data Era: Results of 11 Reports on the Effects of a Multiple-Micronutrient-Intervention Study" Nutrients 16, no. 2: 188. https://doi.org/10.3390/nu16020188