
Walnut (Juglans regia L.) Oligopeptides Alleviate Alcohol-Induced Acute Liver Injury through the Inhibition of Inflammation and Oxidative Stress in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animals and WOPs Administration
2.3. Blood Ethanol Concentration
2.4. Righting Reflex Experiment
2.5. Assay for Serum Biochemical Markers
2.6. Histopathological Examination
2.7. Detection for Oxidative Stress Biomarkers
2.8. Enzyme-Linked Immunosorbent Assay
2.9. Western Blot Analysis
2.10. Statistics
3. Results
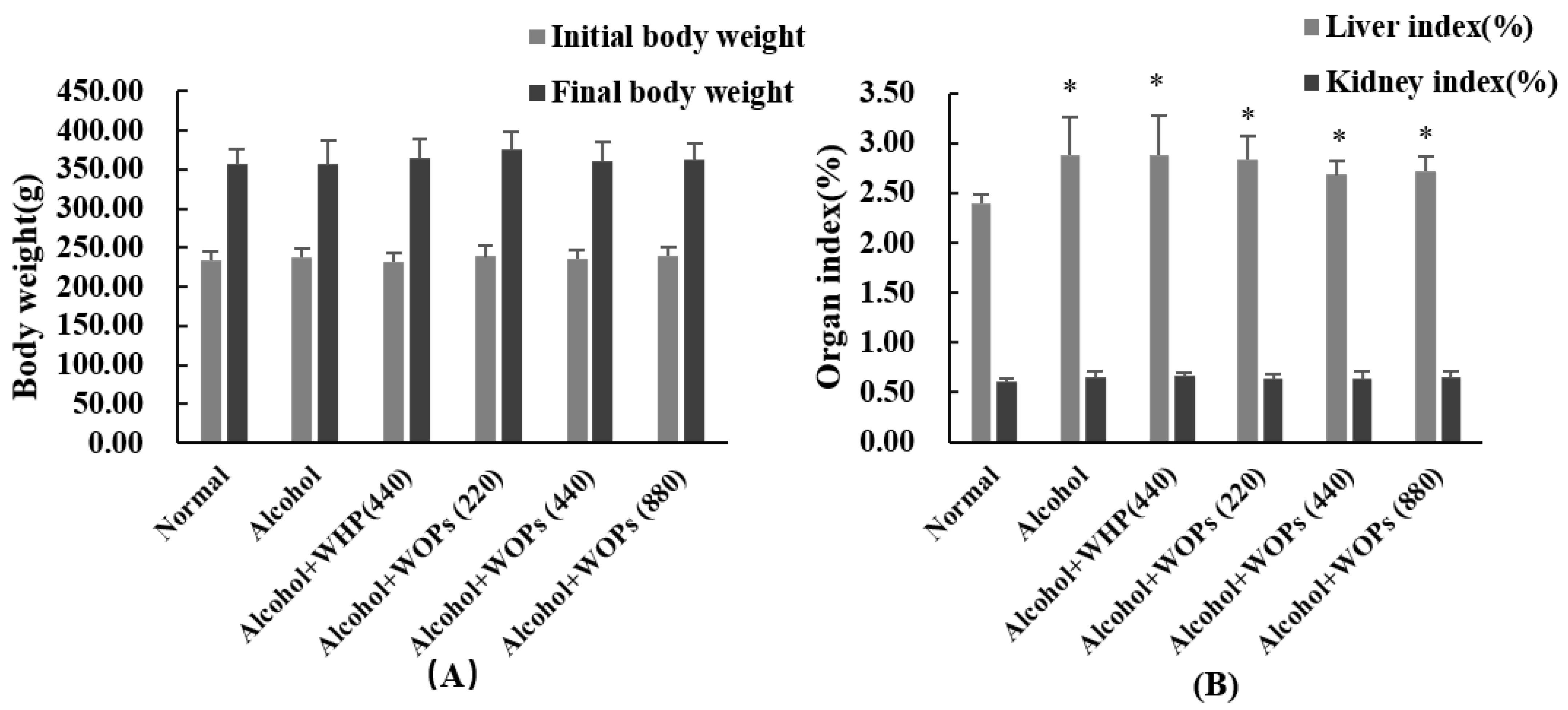
3.1. Effect of WOPs on Body Weight and Organ Index
3.2. The Loss of Righting Reflex (LORR) Assay
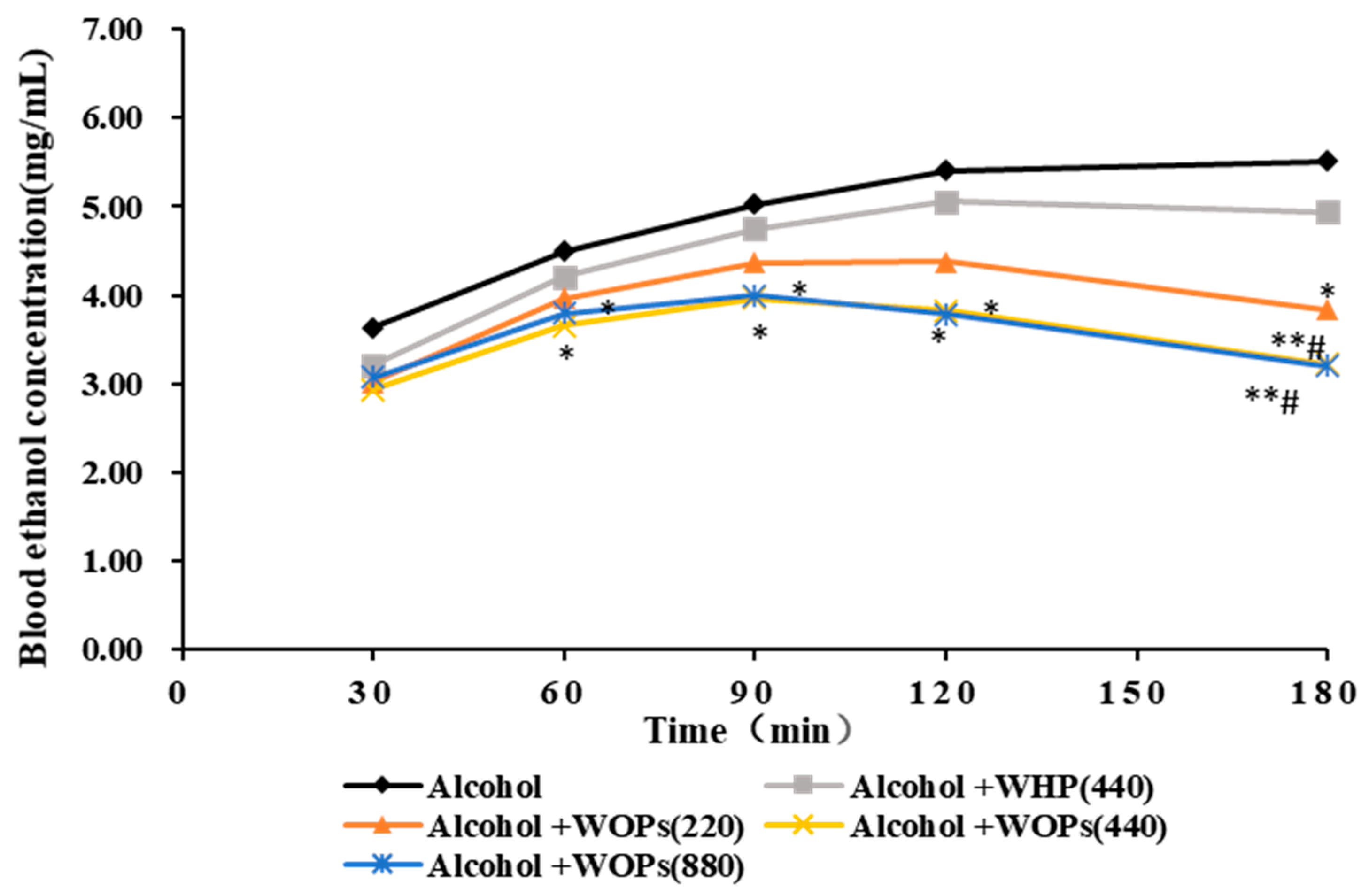
3.3. Effect of WOPs on Blood Ethanol Concentration of Mice
3.4. Effect of WOPs on Serum Biochemical Parameters
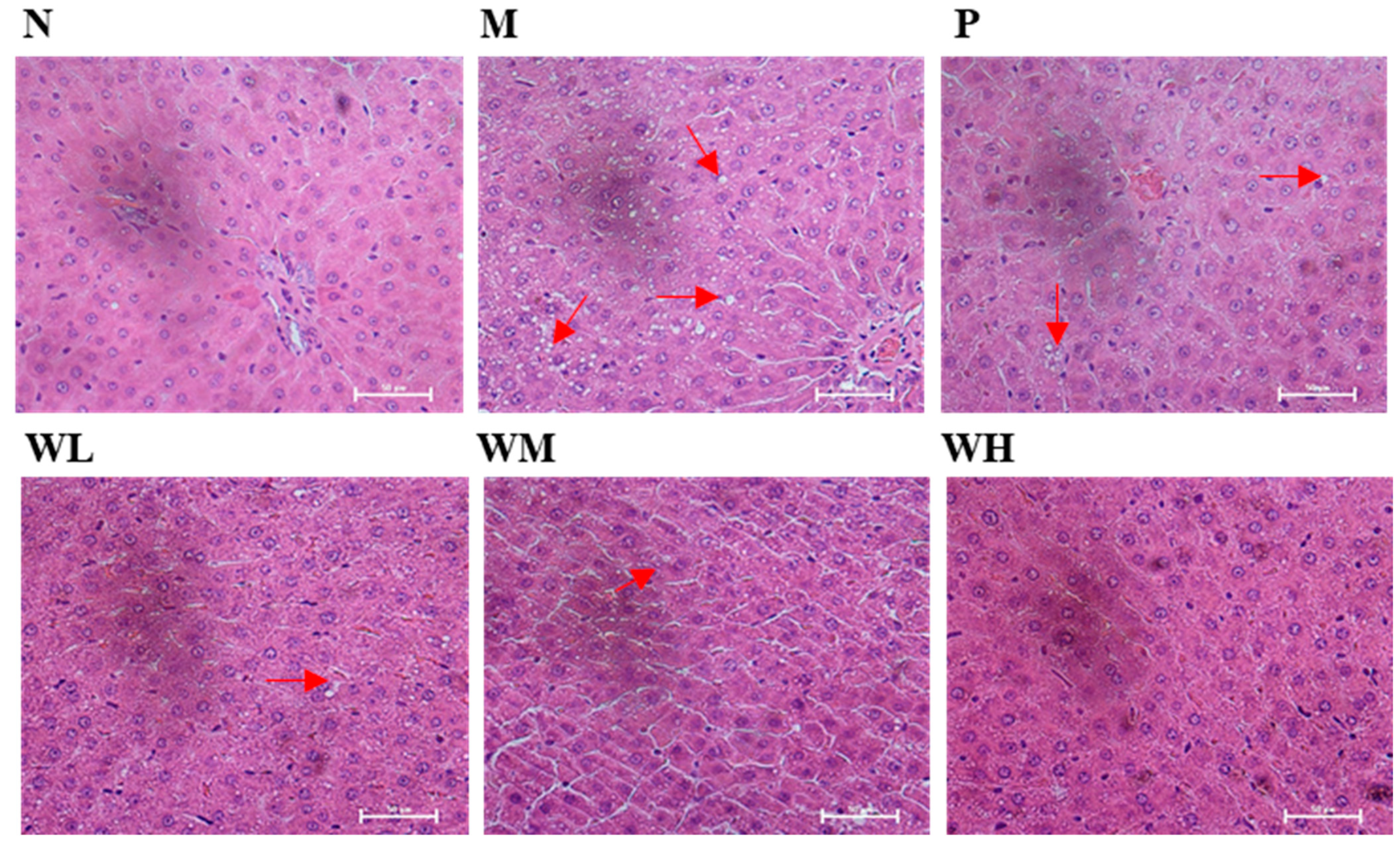
3.5. Pathological Observations and Steatosis Grade
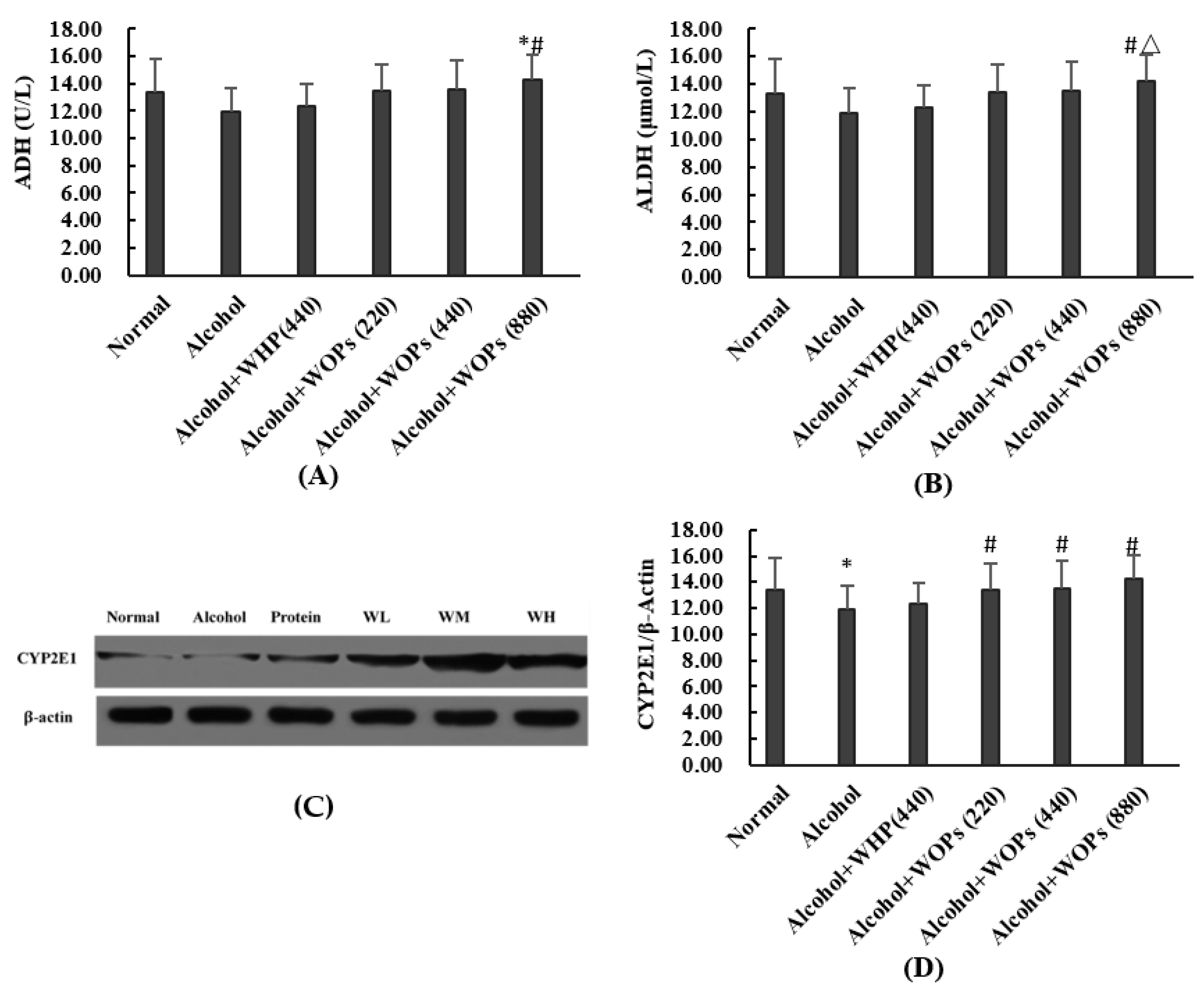
3.6. Effect of WOPs on Alcohol-Metabolic Enzymes in Rat Liver
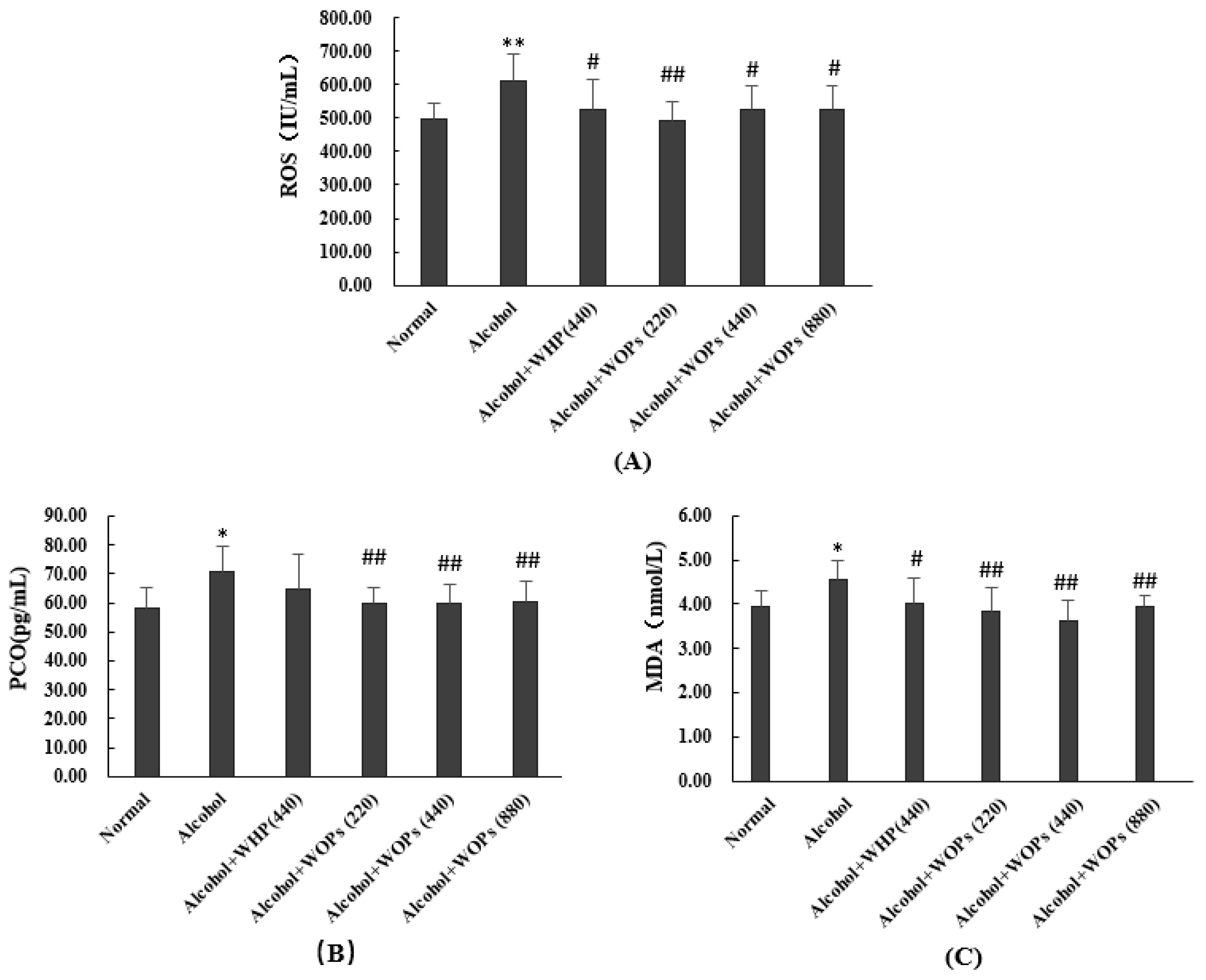
3.7. Effect of WOPs on Oxidative Stress Products in Liver of Rats
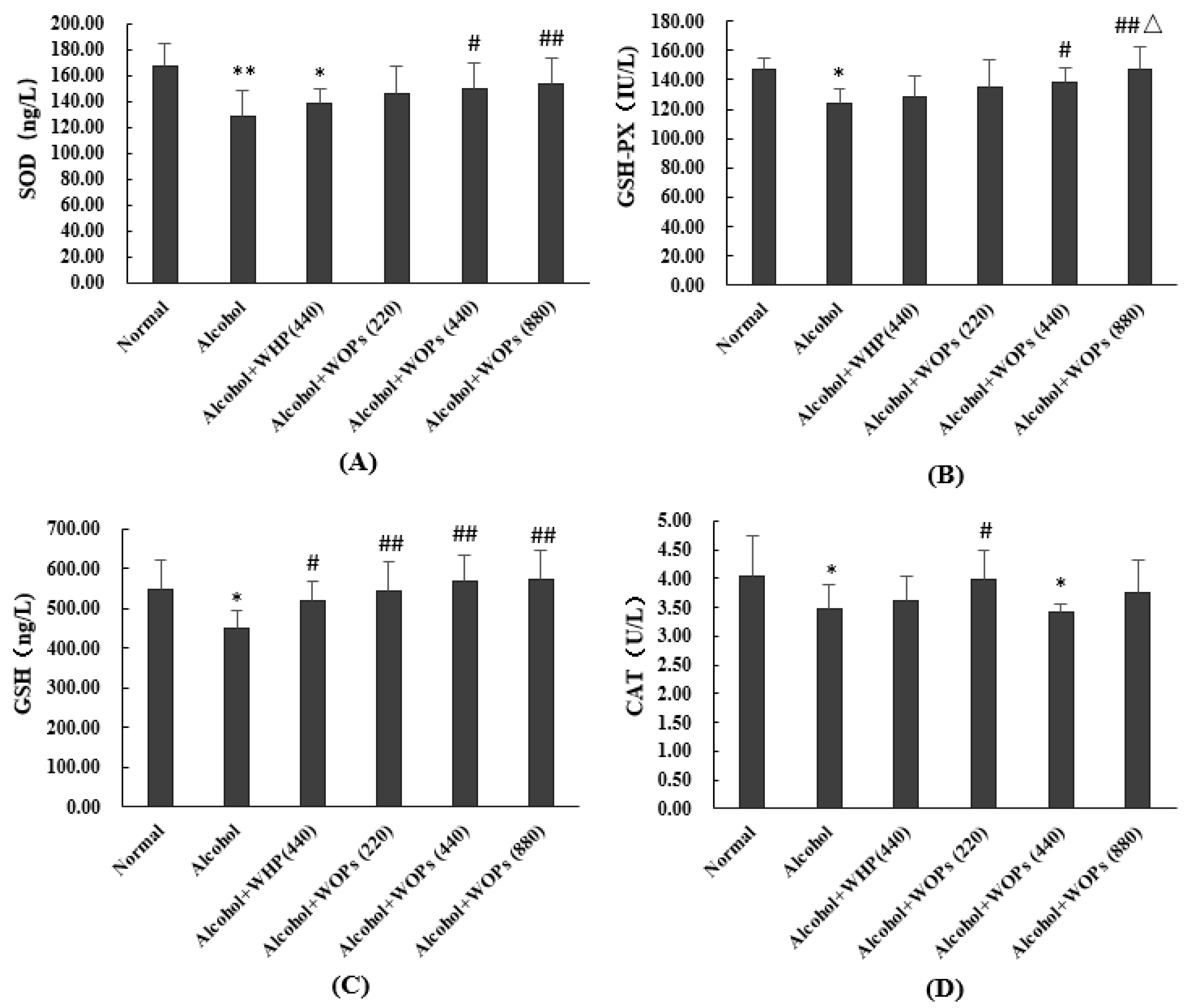
3.8. Effect of WOPs on Anti-Oxidation Capacity in Livers of Rats
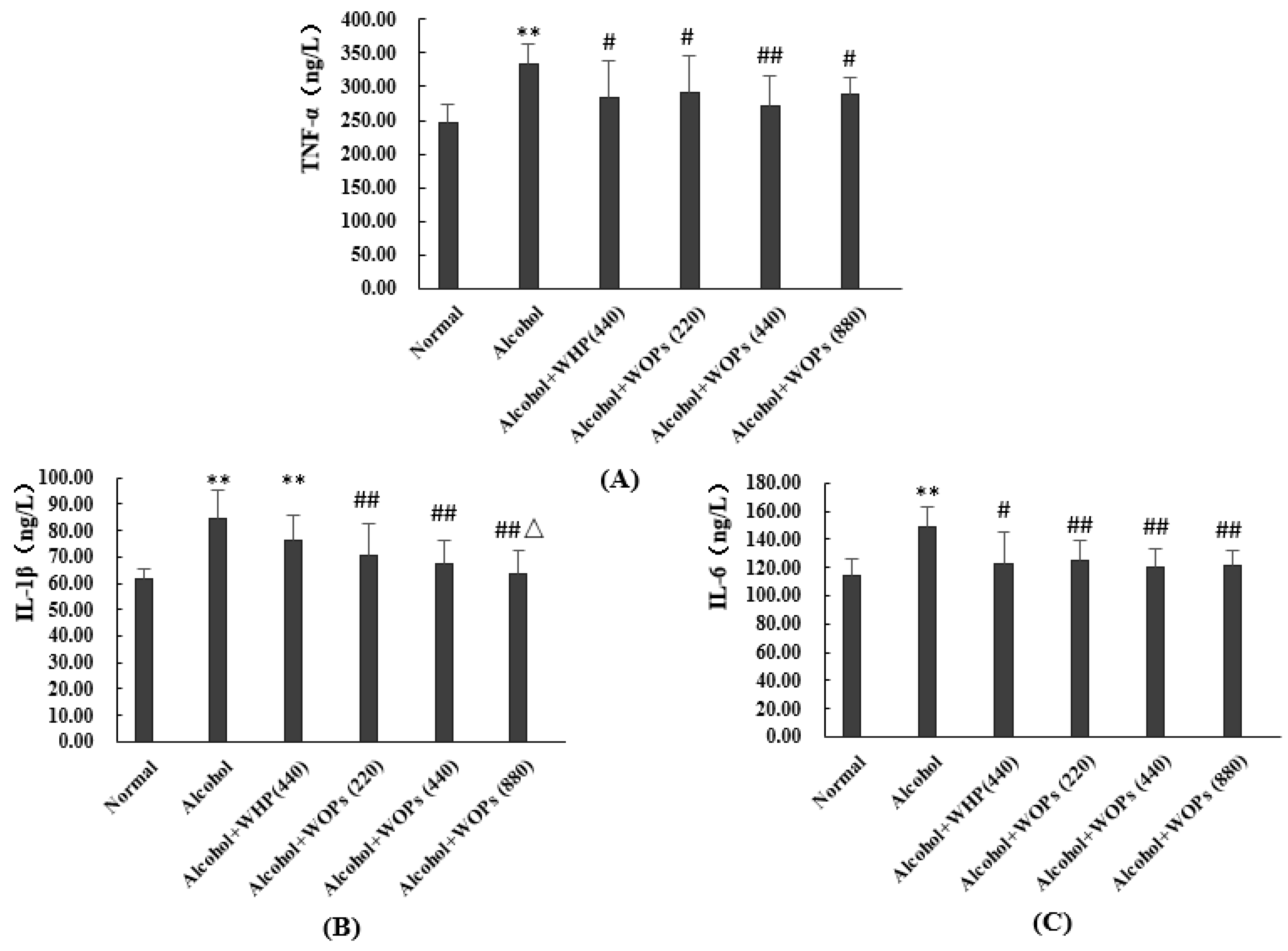
3.9. Effect of WOPs on Serum Pro-Inflammatory Cytokine Levels in Rats
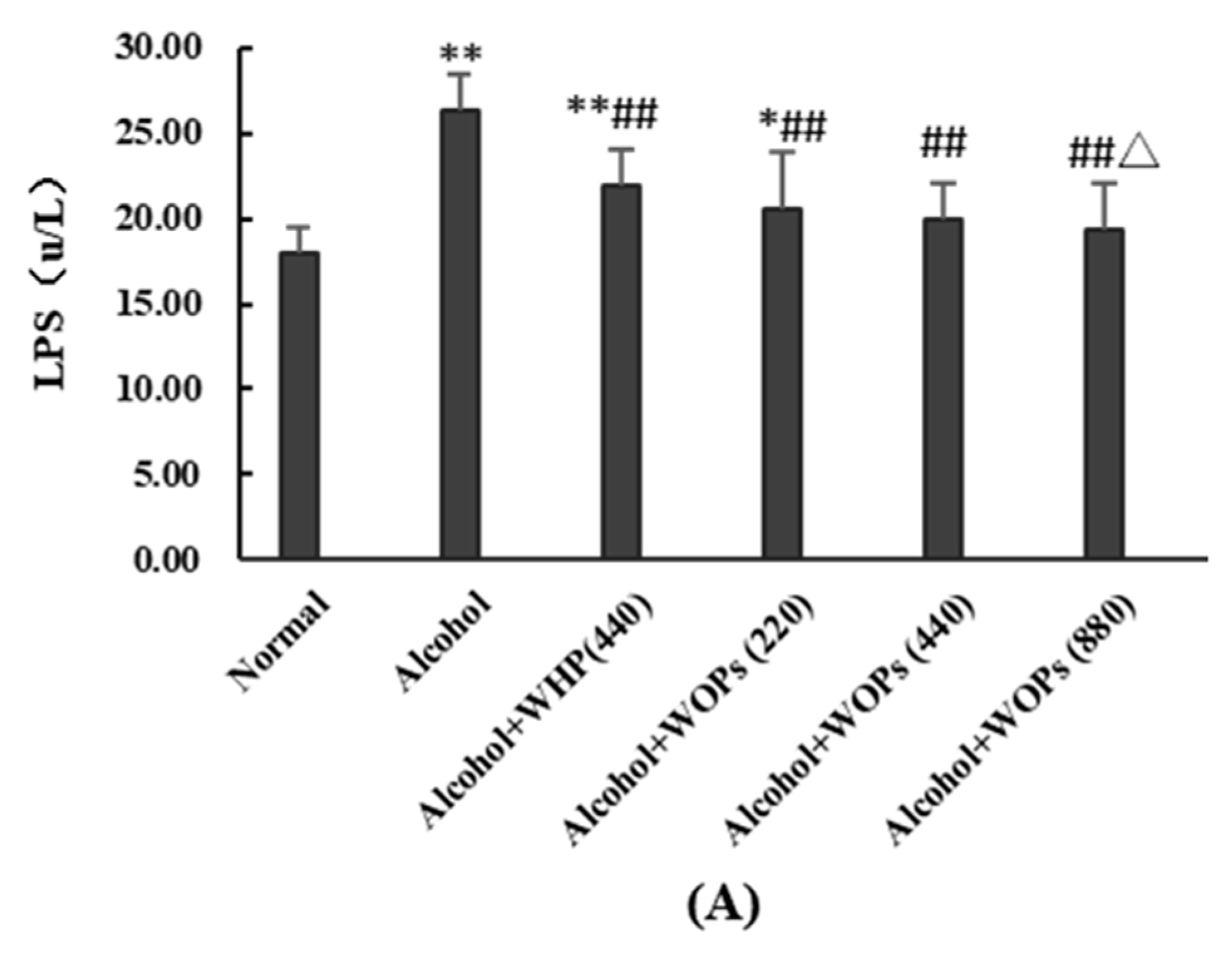
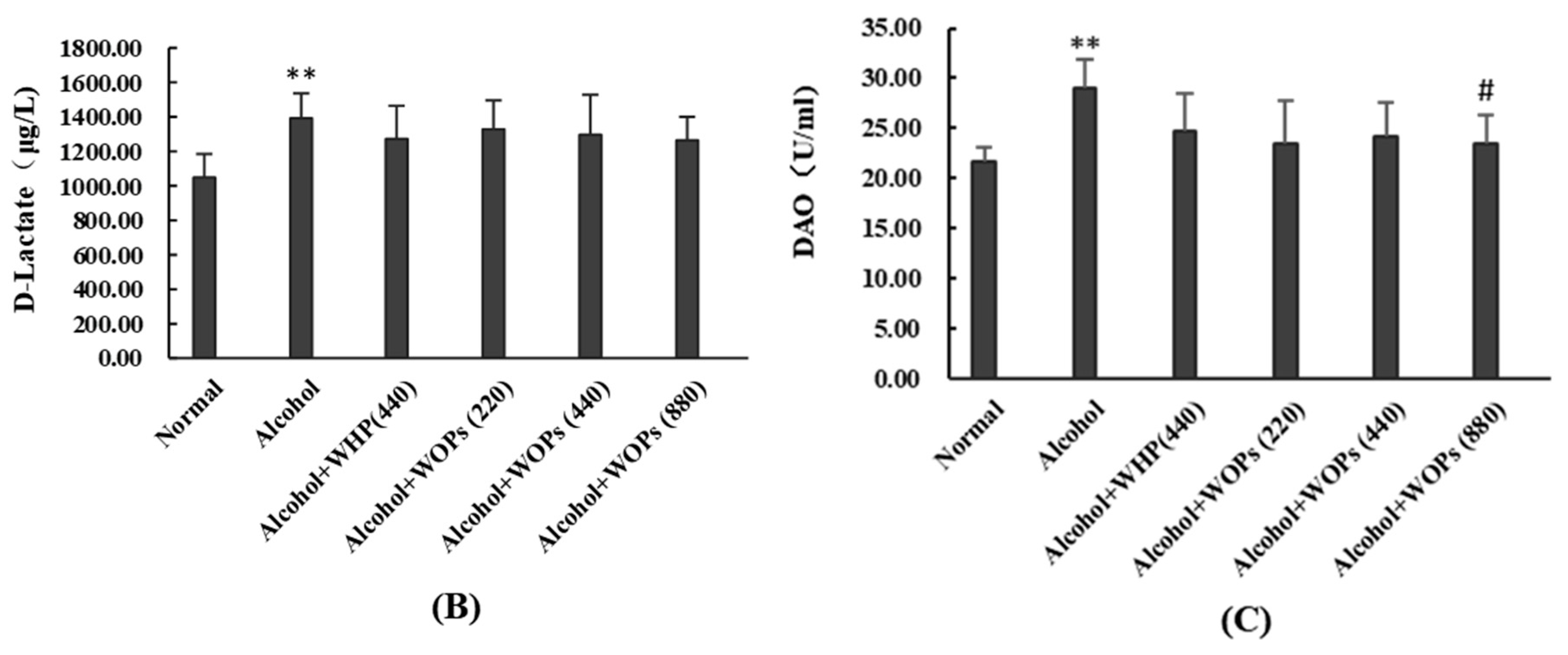
3.10. Effect of WOPs on Intestinal Permeability in Rats
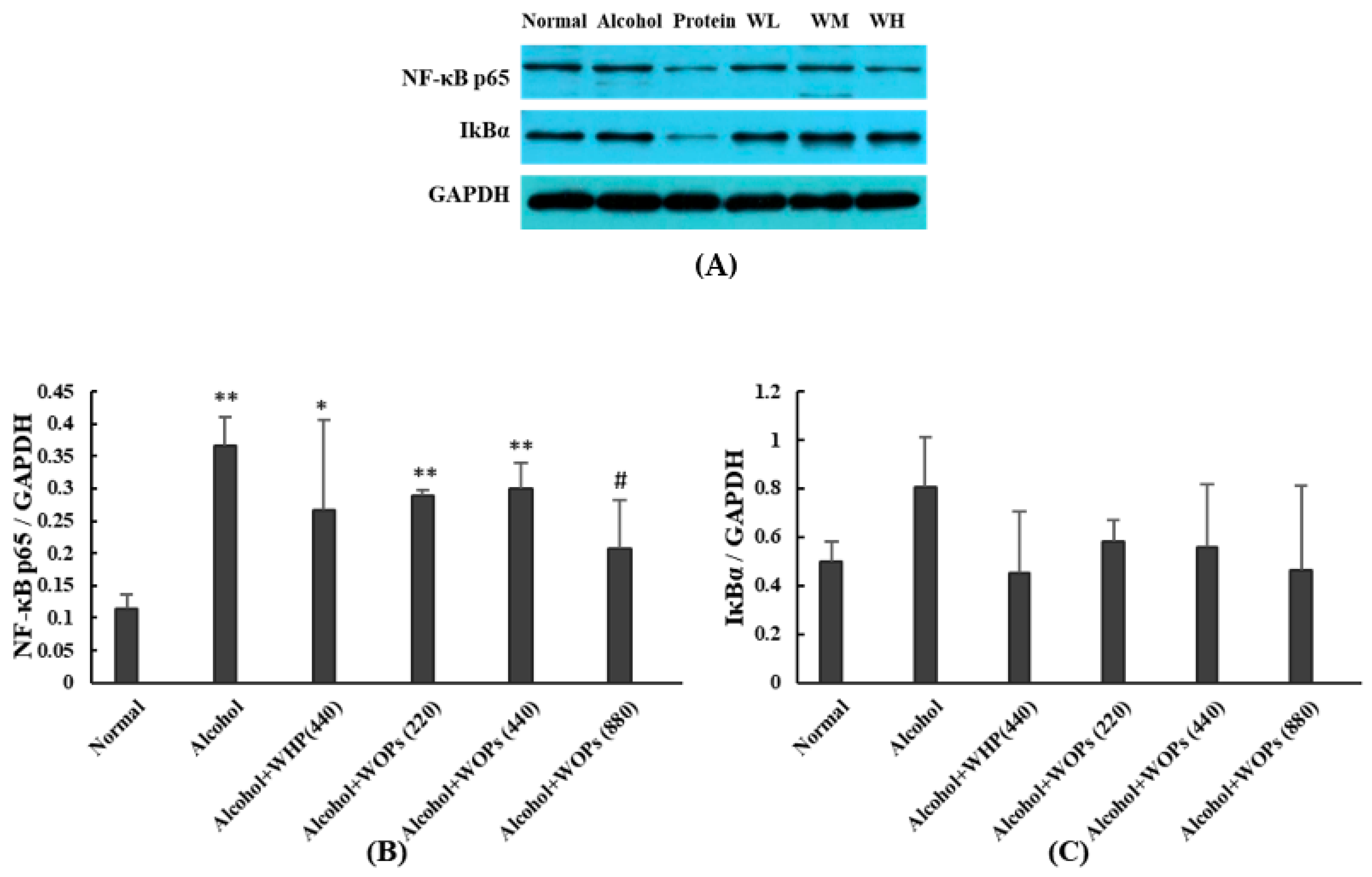
3.11. Effect of WOPs on the Expression of Hepatic NF-κB p65 and IκBα in Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic liver disease. Nat. Rev. Dis. Primers 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Status Report on Alcohol and Health 2018. 2018. Available online: https://www.who.int/publications/i/item/9789241565639 (accessed on 13 January 2023).
- Cioarca-Nedelcu, R.; Atanasiu, V.; Stoian, I. Alcoholic liver disease-from steatosis to cirrhosis—A biochemistry approach. J. Med. Life 2021, 14, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Chang, B.; Sun, Y.; Lin, H.; Li, B.; Teng, G.; Zou, Z.S. Disease spectrum of alcoholic liver disease in Beijing 302 Hospital from 2002 to 2013: A large tertiary referral hospital experience from 7422 patients. Medicine 2017, 96, e6163. [Google Scholar] [CrossRef] [PubMed]
- Axley, P.D.; Richardson, C.T.; Singal, A.K. Epidemiology of Alcohol Consumption and Societal Burden of Alcoholism and Alcoholic Liver Disease. Clin. Liver Dis. 2019, 23, 39–50. [Google Scholar] [CrossRef]
- Singal, A.K.; Mathurin, P. Diagnosis and Treatment of Alcohol-Associated Liver Disease: A Review. Jama 2021, 326, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Hamley, I.W. Small Bioactive Peptides for Biomaterials Design and Therapeutics. Chem. Rev. 2017, 117, 14015–14041. [Google Scholar] [CrossRef] [PubMed]
- Bougle, D.; Bouhallab, S. Dietary bioactive peptides: Human studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 335–343. [Google Scholar] [CrossRef]
- Gao, S.; Jiang, X.; Wang, L.; Jiang, S.; Luo, H.; Chen, Y.; Peng, C. The pathogenesis of liver cancer and the therapeutic potential of bioactive substances. Front. Pharmacol. 2022, 13, 1029601. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, W.; Wang, Y.; Jia, F.; Wang, Y.; Ma, Y.; Gu, R.; Lu, J. Functions and Applications of Bioactive Peptides from Corn Gluten Meal. Adv. Food Nutr. Res. 2019, 87, 1–41. [Google Scholar] [CrossRef]
- Nie, W.; Zhou, K.; Wang, Y.; Wang, Z.M.; Xie, Y.; Zhou, H.; Xu, B.C. Isolation and identification of bioactive peptides from Xuanwei ham that rescue oxidative stress damage induced by alcohol in HHL-5 hepatocytes. Food Funct. 2020, 11, 9710–9720. [Google Scholar] [CrossRef]
- Wei, K.; Wei, Y.; Xu, W.; Lu, F.; Ma, H. Corn peptides improved obesity-induced non-alcoholic fatty liver disease through relieving lipid metabolism, insulin resistance and oxidative stress. Food Funct. 2022, 13, 5782–5793. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Zhou, L.; Gao, J.; Jia, R.; Zheng, Y.; Zhao, S.; Zhao, M.; Toldrá, F. Musculus senhousei as a promising source of bioactive peptides protecting against alcohol-induced liver injury. Food Chem. Toxicol. 2023, 174, 113652. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.; Sussmuth, R.D. Bioactive Peptide Natural Products as Lead Structures for Medicinal Use. Acc. Chem. Res. 2017, 50, 1566–1576. [Google Scholar] [CrossRef] [PubMed]
- Ros, E.; Izquierdo-Pulido, M.; Sala-Vila, A. Beneficial effects of walnut consumption on human health: Role of micronutrients. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 498–504. [Google Scholar] [CrossRef]
- Uddin, Y.; Khan, N.M.; Ali, F.; Ahamd, S.; Wang, J. Estimation of Various Physicochemical Properties of Walnut Oil from Different Areas of Northern Kpk, Pakistan. J. Mex. Chem. Society. 2021, 65, 572–581. [Google Scholar] [CrossRef]
- Ullah, S.F.; Khan, N.M.; Ali, F.; Ahmad, S.; Muhammad, N. Effects of Maillard reaction on physicochemical and functional properties of walnut protein isolate. Food Sci. Biotechnol. 2019, 28, 1391–1399. [Google Scholar] [CrossRef]
- Liu, R.; Hao, Y.T.; Zhu, N.; Liu, X.R.; Kang, J.W.; Mao, R.X.; Hou, C.; Li, Y. The Gastroprotective Effect of Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) against Ethanol-Induced Gastric Mucosal Injury in Rats. Nutrients 2020, 12, 1138. [Google Scholar] [CrossRef]
- Liu, R.; Wu, L.; Du, Q.; Ren, J.W.; Chen, Q.H.; Li, D.; Mao, R.X.; Liu, X.R.; Li, Y. Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) and Their Anti-Fatigue Effects in Mice. Molecules 2018, 24, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Liu, R.; He, L.-X.; Mao, R.-X.; Liu, X.-R.; Zhang, T.; Hao, Y.-T.; Fan, R.; Xu, M.-H.; Li, Y. Radioprotective Effect of Walnut Oligopeptides Against Gamma Radiation-Induced Splenocyte Apoptosis and Intestinal Injury in Mice. Molecules 2019, 24, 1582. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Zhang, J.; Yao, J.; Zhang, B.; Duan, W.; Zhao, C.; Du, P.; Song, J.; Zheng, Y.; Wang, M. Shanxi Aged Vinegar Protects against Alcohol-Induced Liver Injury via Activating Nrf2-Mediated Antioxidant and Inhibiting TLR4-Induced Inflammatory Response. Nutrients 2018, 10, 805. [Google Scholar] [CrossRef] [PubMed]
- Kourkoumpetis, T.; Sood, G. Pathogenesis of Alcoholic Liver Disease: An Update. Clin. Liver Dis. 2019, 23, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chen, Q.H.; Ren, J.W.; Sun, B.; Cai, X.X.; Li, D.; Mao, R.X.; Wu, X.; Li, Y. Ginseng (Panax ginseng Meyer) Oligopeptides Protect Against Binge Drinking-Induced Liver Damage through Inhibiting Oxidative Stress and Inflammation in Rats. Nutrients 2018, 10, 1665. [Google Scholar] [CrossRef] [PubMed]
- Massey, V.L.; Arteel, G.E. Acute alcohol-induced liver injury. Front. Physiol. 2012, 3, 193. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Lemasters, J.J. A Unifying Hypothesis Linking Hepatic Adaptations for Ethanol Metabolism to the Proinflammatory and Profibrotic Events of Alcoholic Liver Disease. Alcohol. Clin. Exp. Res. 2018, 42, 2072–2089. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yu, X.; Zhu, N.; Xu, M.; Li, Y. Beneficial effects of whey protein peptides on muscle loss in aging mice models. Front. Nutr. 2022, 9, 897821. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, Q.; Lin, B.; Yu, Y.; Ding, Y.; Dai, X.; Li, Y. Comparative studies of oral administration of marine collagen peptides from Chum Salmon (Oncorhynchus keta) pre- and post-acute ethanol intoxication in female Sprague-Dawley rats. Food Funct. 2014, 5, 2078–2085. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, J.; Li, Y. Corn oligopeptides protect against early alcoholic liver injury in rats. Food Chem. Toxicol. 2012, 50, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Bao, L.; Wang, N.; Xu, M.; Mao, R.; Li, Y. Dietary Nucleotides Supplementation and Liver Injury in Alcohol-Treated Rats: A Metabolomics Investigation. Molecules 2016, 21, 435. [Google Scholar] [CrossRef]
- Buyco, D.G.; Martin, J.; Jeon, S.; Hooks, R.; Lin, C.; Carr, R. Experimental models of metabolic and alcoholic fatty liver disease. World J. Gastroenterol. 2021, 27, 1–18. [Google Scholar] [CrossRef]
- Ward, L.D.; Tu, H.C.; Quenneville, C.B.; Tsour, S.; Flynn-Carroll, A.O.; Parker, M.M.; Deaton, A.M.; Haslett, P.A.J.; Lotta, L.A.; Verweij, N.; et al. GWAS of serum ALT and AST reveals an association of SLC30A10 Thr95Ile with hypermanganesemia symptoms. Nat. Commun. 2021, 12, 4571. [Google Scholar] [CrossRef]
- Kwo, P.Y.; Cohen, S.M.; Lim, J.K. ACG Clinical Guideline: Evaluation of Abnormal Liver Chemistries. Am. J. Gastroenterol. 2017, 112, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.Z.; Chandimali, N.; Han, Y.H.; Lee, D.H.; Kim, J.S.; Kim, S.U.; Kim, T.D.; Jeong, D.K.; Sun, H.N.; Lee, D.S.; et al. Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease. Int. J. Mol. Sci. 2019, 20, 2712. [Google Scholar] [CrossRef] [PubMed]
- Duparc, T.; Plovier, H.; Marrachelli, V.G.; Van Hul, M.; Essaghir, A.; Ståhlman, M.; Matamoros, S.; Geurts, L.; Pardo-Tendero, M.M.; Druart, C.; et al. Hepatocyte MyD88 affects bile acids, gut microbiota and metabolome contributing to regulate glucose and lipid metabolism. Gut 2017, 66, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Lai, Y.; Huang, P.; Xie, L.; Lin, H.; Zhou, Z.; Mo, C.; Deng, G.; Yan, W.; Gao, Z.; et al. Naringin attenuates alcoholic liver injury by reducing lipid accumulation and oxidative stress. Life Sci. 2019, 216, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Chu, H.; Brandl, K.; Jiang, L.; Zeng, S.; Meshgin, N.; Papachristoforou, E.; Argemi, J.; Mendes, B.G.; Wang, Y.; et al. CRIg on liver macrophages clears pathobionts and protects against alcoholic liver disease. Nat. Commun. 2021, 12, 7172. [Google Scholar] [CrossRef]
- Li, L.X.; Guo, F.F.; Liu, H.; Zeng, T. Iron overload in alcoholic liver disease: Underlying mechanisms, detrimental effects, and potential therapeutic targets. Cell. Mol. Life Sci. 2022, 79, 201. [Google Scholar] [CrossRef]
- Namachivayam, A.; A Valsala Gopalakrishnan. A review on molecular mechanism of alcoholic liver disease. Life Sci. 2021, 274, 119328. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Tan, W.; Liu, X.; Deng, L.; Huang, L.; Wang, X.; Gao, X. New insight and potential therapy for NAFLD: CYP2E1 and flavonoids. Biomed. Pharmacother. 2021, 137, 111326. [Google Scholar] [CrossRef]
- Harjumäki, R.; Pridgeon, C.S.; Ingelman-Sundberg, M. CYP2E1 in Alcoholic and Non-Alcoholic Liver Injury. Roles of ROS, Reactive Intermediates and Lipid Overload. Int. J. Mol. Sci. 2021, 22, 8221. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Lee, Y.S.; Sim, J.; Seo, S.; Seo, W. Alcoholic liver disease: A new insight into the pathogenesis of liver disease. Arch. Pharm. Res. 2022, 45, 447–459. [Google Scholar] [CrossRef]
- Yuan, R.; Tao, X.; Liang, S.; Pan, Y.; He, L.; Sun, J.; Wenbo, J.; Li, X.; Chen, J.; Wang, C. Protective effect of acidic polysaccharide from Schisandra chinensis on acute ethanol-induced liver injury through reducing CYP2E1-dependent oxidative stress. Biomed. Pharmacother. 2018, 99, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Nie, Y.; Luo, M.; Chen, Z.; He, B. Natural Compounds: A Potential Treatment for Alcoholic Liver Disease? Front. Pharmacol. 2021, 12, 694475. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.M.; Cho, Y.E.; Hwang, S. Crosstalk between Oxidative Stress and Inflammatory Liver Injury in the Pathogenesis of Alcoholic Liver Disease. Int. J. Mol. Sci. 2022, 23, 774. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S. Alcohol, liver disease and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.E.; Kim, D.K.; Seo, W.; Gao, B.; Yoo, S.H.; Song, B.J. Fructose Promotes Leaky Gut, Endotoxemia, and Liver Fibrosis Through Ethanol-Inducible Cytochrome P450-2E1-Mediated Oxidative and Nitrative Stress. Hepatology 2021, 73, 2180–2195. [Google Scholar] [CrossRef]
- Schiefer, J.; Baron-Stefaniak, J.; Boehm, T.; Wadowski, P.; Berlakovich, G.; Kuessel, L.; Mühlbacher, J.; Jilma-Stohlawetz, P.; Schwameis, M.; Jilma, B.; et al. Regulation of histamine and diamine oxidase in patients undergoing orthotopic liver transplantation. Sci. Rep. 2020, 10, 822. [Google Scholar] [CrossRef]
- Li, H.; Qiu, P.; Wang, J.; Niu, C.; Pan, S. Effects of compound Ginkgo biloba on intestinal permeability in rats with alcohol-induced liver injury. Food Funct. 2015, 6, 470–478. [Google Scholar] [CrossRef]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef]
- Gao, B.; Tsukamoto, H. Inflammation in Alcoholic and Nonalcoholic Fatty Liver Disease: Friend or Foe? Gastroenterology 2016, 150, 1704–1709. [Google Scholar] [CrossRef]
- Cai, X.; Bao, L.; Wang, N.; Ren, J.; Chen, Q.; Xu, M.; Li, D.; Mao, R.; Li, Y. Dietary nucleotides protect against alcoholic liver injury by attenuating inflammation and regulating gut microbiota in rats. Food Funct. 2016, 7, 2898–2908. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Han, J.; Lee, C.; Yoon, M.; Jung, Y. Pathophysiological Aspects of Alcohol Metabolism in the Liver. Int. J. Mol. Sci. 2021, 22, 5717. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Number | LORR Rate (%) | Latency Time of LORR (min) | Duration Time of LORR (min) |
|---|---|---|---|---|
| Alcohol | 10 | 80 | 35.75 ± 32.14 | 711.25 ± 200.63 |
| Alcohol + WHP (440) | 10 | 80 | 63.88 ± 40.00 | 594.37 ± 198.06 |
| Alcohol + WOPs (220) | 10 | 40 | 78.00 ± 61.17 | 369.25 ± 149.50 ## |
| Alcohol + WOPs (440) | 10 | 30 #Δ | 114.33 ± 50.50 # | 331.67 ± 86.05 ##Δ |
| Alcohol + WOPs (880) | 10 | 30 #Δ | 128.33 ± 76.00 ## | 344.64 ± 107.53 ##Δ |
| Parameters | Normal | Alcohol | Alcohol + WHP (440) | Alcohol + WOPs (220) | Alcohol + WOPs (440) | Alcohol + WOPs (880) |
|---|---|---|---|---|---|---|
| ALT (U/L) | 52.67 ± 10.03 | 85.86 ± 21.79 ** | 71.50 ± 22.05 | 63.78 ± 9.72 # | 59.22 ± 15.43 ## | 64.17 ± 10.40 # |
| AST (U/L) | 169.17 ± 34.30 | 264.22 ± 64.26 ** | 234.78 ± 57.35 * | 200.40 ± 50.87 # | 195.00 ± 42.99 # | 193.56 ± 41.21 # |
| TC (mmol/L) | 1.68 ± 0.22 | 1.52 ± 0.15 | 1.60 ± 0.25 | 1.54 ± 0.28 | 1.64 ± 0.25 | 1.42 ± 0.10 |
| TG (mmol/L) | 0.67 ± 0.149 | 1.07 ± 0.16 ** | 1.05 ± 0.31 ** | 1.08 ± 0.20 ** | 0.94 ± 0.25 * | 0.80 ± 0.11 #Δ |
| HDL-C (mmol/L) | 0.74 ± 0.10 | 0.64 ± 0.10 | 0.75 ± 0.11 | 0.75 ± 0.16 | 0.74 ± 0.19 | 0.70 ± 0.14 |
| LDL-C (mmol/L) | 0.27 ± 0.07 | 0.34 ± 0.07 | 0.31 ± 0.07 | 0.28 ± 0.08 * | 0.29 ± 0.07 | 0.24 ± 0.09 ## |
| VLDL (μg/mL) | 175.15 ± 18.81 | 187.32 ± 18.95 | 180.80 ± 127.32 | 177.65 ± 28.39 | 163.07 ± 25.80 | 160.95 ± 16.09 # |
| TP (g/L) | 71.62 ± 5.85 | 55.10 ± 5.05 ** | 58.60 ± 6.50 ** | 60.21 ± 7.11 ** | 63.79 ± 7.87 **## | 63.44 ± 7.47 **## |
| ALB (g/L) | 36.33 ± 2.72 | 30.91 ± 3.52 * | 31.46 ± 3.06 * | 32.19 ± 2.70 * | 33.45 ± 3.93 | 33.64 ± 2.26 |
| GLB (g/L) | 35.28 ± 3.88 | 25.92 ± 4.41 ** | 27.14 ± 3.61 ** | 28.02 ± 4.64 ** | 30.29 ± 4.50 **## | 30.67 ± 4.73 **## |
| A:G ratio | 1.03 ± 0.06 | 1.20 ± 0.13 ** | 1.17 ± 0.07 ** | 1.16 ± 0.12 ** | 1.06 ± 0.10 ##Δ | 1.08 ± 0.11 ## |
| ALP (U/L) | 142.17 ± 25.63 | 278.09 ± 37.95 ** | 276.75 ± 50.33 ** | 206.10 ± 32.51 *## | 205.64 ± 32.50 *## | 203.33 ± 18.17 *##Δ |
| CR (umol/L) | 51.83 ± 7.86 | 42.40 ± 5.27 * | 42.38 ± 5.58 * | 43.13 ± 8.74 * | 44.00 ± 8.37 * | 45.00 ± 5.03 |
| BUN (mmol/L) | 5.93 ± 1.59 | 6.77 ± 1.79 | 6.40 ± 2.23 | 5.75 ± 1.96 | 5.64 ± 1.97 | 5.55 ± 1.15 |
| TBil (umol/L) | 1.68 ± 0.33 | 0.62 ± 0.64 ** | 0.54 ± 0.33 ** | 0.69 ± 0.38 ** | 0.55 ± 0.40 ** | 0.71 ± 0.41 ** |
| Group | Number under Each Steatosis Grade | Average Rank | ||||
|---|---|---|---|---|---|---|
| 0 | I | II | III | IV | ||
| Normal | 6 | 0 | 0 | 0 | 0 | 6.42 |
| Alcohol | 0 | 0 | 1 | 2 | 3 | 30.42 * |
| Alcohol + WHP (440) | 1 | 1 | 2 | 1 | 1 | 23.00 |
| Alcohol + WOPs (220) | 0 | 3 | 2 | 1 | 0 | 17.50 |
| Alcohol + WOPs (440) | 1 | 1 | 2 | 2 | 0 | 18.92 |
| Alcohol + WOPs (880) | 2 | 2 | 1 | 1 | 0 | 14.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Hao, Y.-T.; Zhu, N.; Liu, X.-R.; Mao, R.-X.; Kang, J.-W.; Hou, C.; Zhang, T.; Li, Y. Walnut (Juglans regia L.) Oligopeptides Alleviate Alcohol-Induced Acute Liver Injury through the Inhibition of Inflammation and Oxidative Stress in Rats. Nutrients 2023, 15, 2210. https://doi.org/10.3390/nu15092210
Liu R, Hao Y-T, Zhu N, Liu X-R, Mao R-X, Kang J-W, Hou C, Zhang T, Li Y. Walnut (Juglans regia L.) Oligopeptides Alleviate Alcohol-Induced Acute Liver Injury through the Inhibition of Inflammation and Oxidative Stress in Rats. Nutrients. 2023; 15(9):2210. https://doi.org/10.3390/nu15092210
Chicago/Turabian StyleLiu, Rui, Yun-Tao Hao, Na Zhu, Xin-Ran Liu, Rui-Xue Mao, Jia-Wei Kang, Chao Hou, Ting Zhang, and Yong Li. 2023. "Walnut (Juglans regia L.) Oligopeptides Alleviate Alcohol-Induced Acute Liver Injury through the Inhibition of Inflammation and Oxidative Stress in Rats" Nutrients 15, no. 9: 2210. https://doi.org/10.3390/nu15092210