
Characterization of Green and Yellow Papaya (Carica papaya) for Anti-Diabetic Activity in Liver and Myoblast Cells and Wound-Healing Activity in Fibroblast Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Papaya Samples’ Preparation
2.3. Determination of Total Polyphenols
2.4. Determination of Total Flavonoids
2.5. Antioxidant Assays
2.5.1. DPPH (2,2-Diphenyl-1-Picryhydrazyl) Assay
2.5.2. FRAP (Ferric-Reducing Antioxidant Potential (FRAP) Assay
2.6. Cell Culture
2.7. Glucose Uptake Assays Using 2-NBDG
2.8. Glut-2 Expression in HepG2 Cells
2.9. Glut 4 Expression in C2 C12 Myoblast Cells, USA
2.10. Triglyceride Assays
2.11. Wound-Healing Scratch Assay
2.12. Collagen Sircol Dye Binding Assay
3. Results
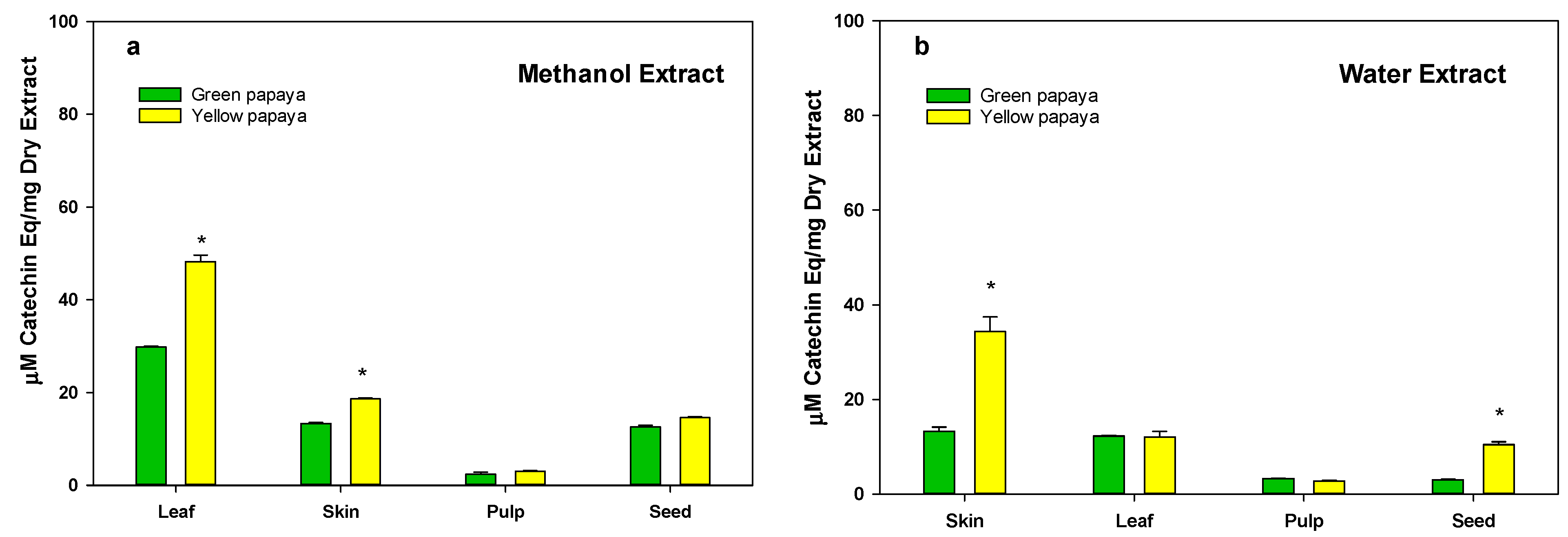
3.1. Polyphenolic Contents in Green and Yellow Papaya
3.2. Total Flavonoid Content in Green and Yellow Papaya
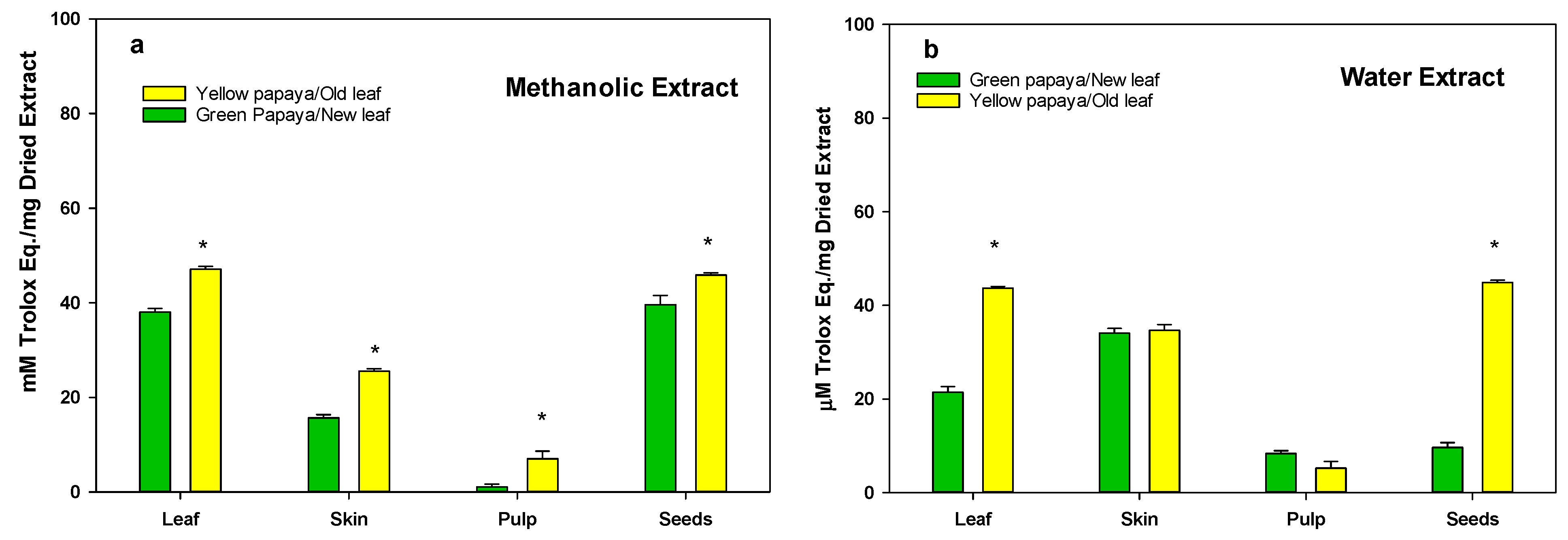
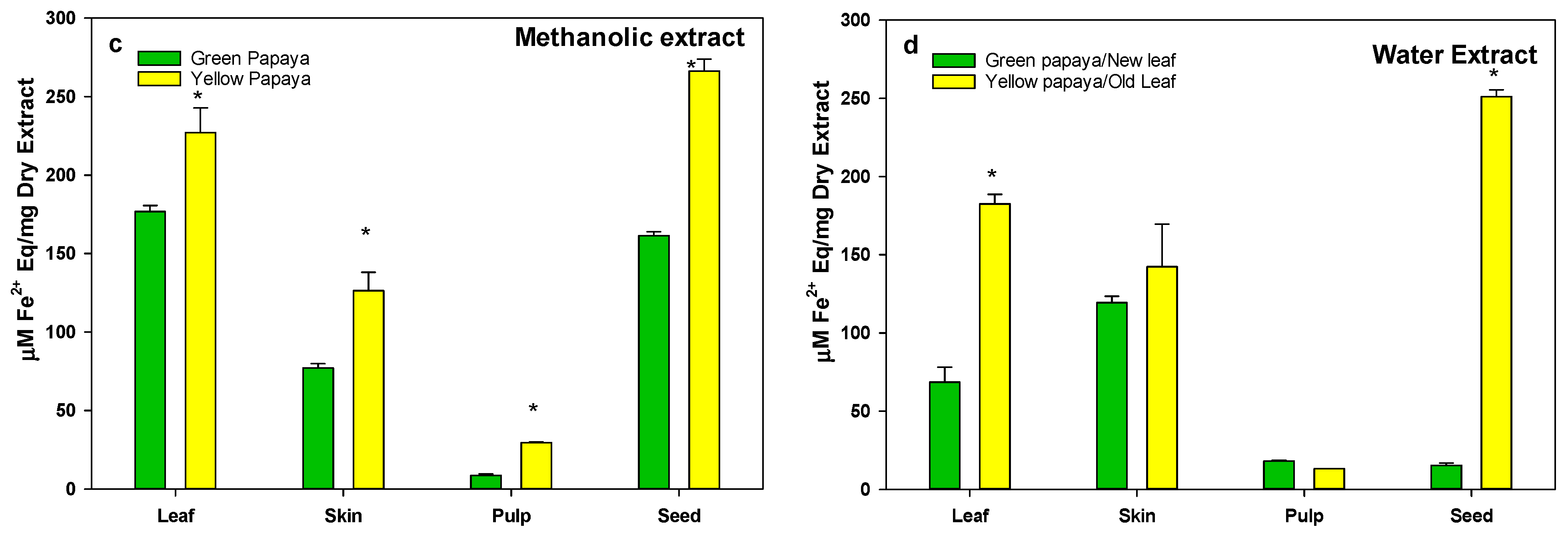
3.3. Anti-Oxidation Activity in Green and Yellow Papaya Fractions
3.4. Glucose Uptake Stimulatory Activity
3.5. Glucose Transporter, Glut-2 Expression
3.6. Glucose Uptake Activity in Myoblast Cells
3.7. Triglyceride Lowering Effects
3.8. Wound-Healing Properties of Papaya Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. National Diabetes Statistics Report Website. Available online: https://www.cdc.gov/diabetes/data/statistics-report/index.html (accessed on 2 February 2023).
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2009, 32, S62–S67. [Google Scholar] [CrossRef] [PubMed]
- IDF Diabetes Atlas. 2021. Available online: https://diabetesatlas.org/atlas/tenth-edition/ (accessed on 10 February 2023).
- Geiss, L.S.; Li, Y.; Hora, I.; Albright, A.; Rolka, D.; Gregg, E.W. Resurgence of Diabetes-Related Nontraumatic Lower-Extremity Amputation in the Young and Middle-Aged Adult U.S. Population. Diabetes Care 2019, 42, 50–54. [Google Scholar] [CrossRef]
- Petersen, M. Economic Costs of Diabetes in the U.S. in 2017. Diabetes Care 2018, 41, 917–928. [Google Scholar]
- Wang, P.Y.; Fang, J.C.; Gao, Z.H.; Zhang, C.; Xie, S.Y. Higher intake of fruits, vegetables or their fiber reduces the risk of type 2 diabetes: A meta-analysis. J. Diabetes Investig. 2016, 7, 56–69. [Google Scholar] [CrossRef]
- Derosa, G.; Limas, C.P.; Macías, P.C.; Estrella, A.; Maffioli, P. Dietary and nutraceutical approach to type 2 diabetes. Arch. Med. Sci. 2014, 10, 336–344. [Google Scholar] [CrossRef]
- Yadava, U.; Burris, A.; McCrary, D. Papaya: A Potential Annual Crop under Middle Georgia Conditions; Timber Press: Portland, OR, USA, 1990. [Google Scholar]
- Crane, J. Papaya Growing in the Florida Home Landscape; University of Florida: Gainesville, FL, USA, 2005; Volume HS11, pp. 1–7. [Google Scholar]
- Krishna, K.; Paridhavi, M.; Patel, J. Review on nutritional, medicinal and pharmacological properties of Papaya (Cirica papaya Linn.). Nat. Prod. Radiance 2008, 7, 364–373. [Google Scholar]
- Checker, R.; Patwardhan, R.S.; Sharma, D.; Menon, J.; Thoh, M.; Sandur, S.K.; Sainis, K.B.; Poduval, T.B. Plumbagin, a Vitamin K3 Analogue, abrogates Lipopolysaccharide-Induced Oxidative Stress, Inflammation and Endotoxic Shock via NF-kappa B Suppression. Inflammation 2014, 37, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Maniyar, Y.; Bhixavatimath, P. Antihyperglycemic and hypolipidemic activities of aqueous extract of Carica papaya Linn. leaves in alloxan-induced diabetic rats. J. Ayurveda Integr. Med. 2012, 3, 70–74. [Google Scholar] [CrossRef]
- Juárez-Rojop, I.E.; Díaz-Zagoya, J.C.; Ble-Castillo, J.L.; Miranda-Osorio, P.H.; Castell-Rodríguez, A.E.; Tovilla-Zárate, C.A.; Rodríguez-Hernández, A.; Aguilar-Mariscal, H.; Ramón-Frías, T.; Bermúdez-Ocaña, D.Y. Hypoglycemic effect of Carica papaya leaves in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2012, 12, 236. [Google Scholar] [CrossRef]
- Nayak, B.S.; Ramdeen, R.; Adogwa, A.; Ramsubhag, A.; Marshall, J.R. Wound-healing potential of an ethanol extract of Carica papaya (Caricaceae) seeds. Int. Wound J. 2012, 9, 650–655. [Google Scholar] [CrossRef]
- Singh, O.; Ali, M. Phytochemical and antifungal profiles of the seeds of Carica papaya L. Indian J. Pharm. Sci. 2011, 73, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Maisarah, A.M.; Nurul Amira, B.; Asmah, R.; Fauziah, O. Antioxidant analysis of different parts of Carica papaya. Int. Food Res. J. 2013, 20, 1043–1048. [Google Scholar]
- Otsuki, N.; Dang, N.H.; Kumagai, E.; Kondo, A.; Iwata, S.; Morimoto, C. Aqueous extract of Carica papaya leaves exhibits anti-tumor activity and immunomodulatory effects. J. Ethnopharmacol. 2010, 127, 760–767. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Parat, M.O.; Hodson, M.P.; Pan, J.; Shaw, P.N.; Hewavitharana, A.K. Chemical Characterization and in Vitro Cytotoxicity on Squamous Cell Carcinoma Cells of Carica papaya Leaf Extracts. Toxins 2015, 8, 7. [Google Scholar] [CrossRef]
- Runnie, I.; Salleh, M.N.; Mohamed, S.; Head, R.J.; Abeywardena, M.Y. Vasorelaxation induced by common edible tropical plant extracts in isolated rat aorta and mesenteric vascular bed. J. Ethnopharmacol. 2004, 92, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Azarkan, M.; El Moussaoui, A.; van Wuytswinkel, D.; Dehon, G.; Looze, Y. Fractionation and purification of the enzymes stored in the latex of Carica papaya. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 790, 229–238. [Google Scholar] [CrossRef]
- Sapaat, A.; Satrija, F.; Mahsol, H.H.; Ahmad, A.H. Anthelmintic activity of papaya seeds on Hymenolepis diminuta infections in rats. Trop. Biomed. 2012, 29, 508–512. [Google Scholar]
- Satrija, F.; Nansen, P.; Murtini, S.; He, S. Anthelmintic activity of papaya latex against patent Heligmosomoides polygyrus infections in mice. J. Ethnopharmacol. 1995, 48, 161–164. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Shaw, P.N.; Parat, M.O.; Hewavitharana, A.K. Anticancer activity of Carica papaya: A review. Mol. Nutr. Food Res. 2013, 57, 153–164. [Google Scholar] [CrossRef]
- Aruoma, O.I.; Somanah, J.; Bourdon, E.; Rondeau, P.; Bahorun, T. Diabetes as a risk factor to cancer: Functional role of fermented papaya preparation as phytonutraceutical adjunct in the treatment of diabetes and cancer. Mutat. Res. 2014, 768, 60–68. [Google Scholar] [CrossRef]
- Tang, C.; Syed, M.; Hamilton, R. Benzyl isothiocyanate in the Caricaceae. Mol. Nutr. Food Res. 1972, 11, 2531–2533. [Google Scholar] [CrossRef]
- Emeruwa, A.C. Antibacterial substance from Carica papaya fruit extract. J. Nat. Prod. 1982, 45, 123–127. [Google Scholar] [CrossRef]
- Nayak, S.B.; Pinto Pereira, L.; Maharaj, D. Wound healing activity of Carica papaya L. in experimentally induced diabetic rats. Indian J. Exp. Biol. 2007, 45, 739–743. [Google Scholar]
- Li, H.; Rafie, R.; Xu, Z.; Siddiqui, R.A. Phytochemical profile and anti-oxidation activity changes during ginger (Zingiber officinale) harvest: Baby ginger attenuates lipid accumulation and ameliorates glucose uptake in HepG2 cells. Food Sci. Nutr. 2022, 10, 133–144. [Google Scholar] [CrossRef]
- Al-Shaya, H.M.; Li, H.; Beg, O.U.; Hamama, A.A.; Witiak, S.M.; Kaseloo, P.; Siddiqui, R.A. Phytochemical profile and antioxidation activity of annona fruit and its effect on lymphoma cell proliferation. Food Sci. Nutr. 2020, 8, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E. Scratch Wound Healing Assay. Methods Mol. Biol. 2020, 2109, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Lareu, R.R.; Zeugolis, D.I.; Abu-Rub, M.; Pandit, A.; Raghunath, M. Essential modification of the Sircol Collagen Assay for the accurate quantification of collagen content in complex protein solutions. Acta Biomater. 2010, 6, 3146–3151. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Pastrana, D.M.; Yahia, E.M.; González-Aguilar, G.A. Phenolic and carotenoid profiles of papaya fruit (Carica papaya L.) and their contents under low temperature storage. J. Sci. Food Agric. 2010, 90, 2358–2365. [Google Scholar] [CrossRef]
- Abdel-Halim, S.; Ibrahim, M.; Abdel Mohsen, M.; Abou-Setta, L.; Sleem, A.; El-Missiry, M. The influence of the extraction method on polyphenols, flavonoids composition and anti-hyperlipidemic properties of papaya leaves (Carica papaya Linn.). Bull. Natl. Res. Cent. 2021, 45, 85. [Google Scholar] [CrossRef]
- Turkmen, N.; Velioglu, Y.S.; Sari, F.; Polat, G. Effect of extraction conditions on measured total polyphenol contents and antioxidant and antibacterial activities of black tea. Molecules 2007, 12, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, S.; Sumathi, V.; Jegathambigai, N.R.; Latha, L.Y. Antihyperglycaemic effects of ethanol extracts of Carica papaya and Pandanus amaryfollius leaf in streptozotocin-induced diabetic mice. Nat. Prod. Res. 2011, 25, 1982–1987. [Google Scholar] [CrossRef]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [PubMed]
- Nesto, R.W.; Bell, D.; Bonow, R.O.; Fonseca, V.; Grundy, S.M.; Horton, E.S.; Le Winter, M.; Porte, D.; Semenkovich, C.F.; Smith, S.; et al. Thiazolidinedione use, fluid retention, and congestive heart failure: A consensus statement from the American Heart Association and American Diabetes Association. Circulation 2003, 108, 2941–2948. [Google Scholar] [CrossRef]
- Onakpa, M.M.; Ajugbonna, O.P. Antidiabetic Potentials Of Cassia occidentalis Leaf Extract on Alloxan Induced Diabetic Albino Mice. Int. J. PharmTech Res. 2012, 4, 1766–1769. [Google Scholar]
- Zhao, R.Y.; He, X.W.; Shan, Y.M.; Zhu, L.L.; Zhou, Q. A stewardship intervention program for safe medication management and use of antidiabetic drugs. Clin. Interv. Aging 2015, 10, 1201–1212. [Google Scholar] [CrossRef]
- Sola, D.; Rossi, L.; Schianca, G.P.; Maffioli, P.; Bigliocca, M.; Mella, R.; Corlianò, F.; Fra, G.P.; Bartoli, E.; Derosa, G. Sulfonylureas and their use in clinical practice. Arch. Med. Sci. 2015, 11, 840–848. [Google Scholar] [CrossRef]
- Aquilante, C.L. Sulfonylurea pharmacogenomics in Type 2 diabetes: The influence of drug target and diabetes risk polymorphisms. Expert Rev. Cardiovasc. Ther. 2010, 8, 359–372. [Google Scholar] [CrossRef]
- Jayaraman, S.; Roy, A.; Vengadassalapathy, S.; Sekar, R.; Veeraraghavan, V.P.; Rajagopal, P.; Rengasamy, G.; Mukherjee, R.; Sekar, D.; Manjunathan, R. An Overview on the Therapeutic Function of Foods Enriched with Plant Sterols in Diabetes Management. Antioxidants 2021, 10, 1903. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Aspects Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Caruso, C.; Baldwin, S.A.; Panico, M.; Blench, I.; Morris, H.R.; Allard, W.J.; Lienhard, G.E.; Lodish, H.F. Sequence and structure of a human glucose transporter. Science 1985, 229, 941–945. [Google Scholar] [CrossRef] [PubMed]
- James, D.E.; Brown, R.; Navarro, J.; Pilch, P.F. Insulin-regulatable tissues express a unique insulin-sensitive glucose transport protein. Nature 1988, 333, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Kayano, T.; Fukumoto, H.; Eddy, R.L.; Fan, Y.S.; Byers, M.G.; Shows, T.B.; Bell, G.I. Evidence for a family of human glucose transporter-like proteins. Sequence and gene localization of a protein expressed in fetal skeletal muscle and other tissues. J. Biol. Chem. 1988, 263, 15245–15248. [Google Scholar] [CrossRef]
- Uldry, M.; Ibberson, M.; Hosokawa, M.; Thorens, B. GLUT2 is a high affinity glucosamine transporter. FEBS Lett. 2002, 524, 199–203. [Google Scholar] [CrossRef]
- Tirone, T.A.; Brunicardi, F.C. Overview of glucose regulation. World J. Surg. 2001, 25, 461–467. [Google Scholar] [CrossRef]
- Krentz, A.J.; Bailey, C.J. Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs 2005, 65, 385–411. [Google Scholar] [CrossRef]
- Karim, S.; Adams, D.H.; Lalor, P.F. Hepatic expression and cellular distribution of the glucose transporter family. World J. Gastroenterol. 2012, 18, 6771–6781. [Google Scholar] [CrossRef]
- Roy, J.R.; Janaki, C.S.; Jayaraman, S.; Periyasamy, V.; Balaji, T.; Vijayamalathi, M.; Veeraraghavan, V.P. Carica papaya Reduces Muscle Insulin Resistance via IR/GLUT4 Mediated Signaling Mechanisms in High Fat Diet and Streptozotocin-Induced Type-2 Diabetic Rats. Antioxidants 2022, 11, 2081. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Hsieh, C.; Chiang, W.C. Effects of guava leaf extract on glucose and lipid homeostasis in diet-induced insulin-resistant C57BL/6J mice. J. Food Sci. Technol. 2018, 3, 305–317. [Google Scholar]
- Wulansari, D.; Basori, A.; Suhartati, S. Effect of Papaya Seed Extract (Carica papaya Linn.) on Glucose Transporter 4 (GLUT 4) Expression of Skeletal Muscle Tissue in Diabetic Mice Induced by High Fructose Diet. Tradit. Med. J. 2017, 22, 131–137. [Google Scholar] [CrossRef]
- WHO. WHO Obesity and Overweight. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 February 2023).
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Od-Ek, P.; Deenin, W.; Malakul, W.; Phoungpetchara, I.; Tunsophon, S. Anti-obesity effect of carica papaya in high-fat diet fed rats. Biomed. Rep. 2020, 13, 30. [Google Scholar] [CrossRef] [PubMed]
- Zimmet, P.Z. Diabetes and its drivers: The largest epidemic in human history? Clin. Diabetes Endocrinol. 2017, 3, 1. [Google Scholar] [CrossRef]
- Wells, J.C.K. The diabesity epidemic in the light of evolution: Insights from the capacity-load model. Diabetologia 2019, 62, 1740–1750. [Google Scholar] [CrossRef]
- Al-Goblan, A.S.; Al-Alfi, M.A.; Khan, M.Z. Mechanism linking diabetes mellitus and obesity. Diabetes Metab. Syndr. Obes. 2014, 7, 587–591. [Google Scholar] [CrossRef]
- Liang, W.; Ye, D.D. The potential of adipokines as biomarkers and therapeutic agents for vascular complications in type 2 diabetes mellitus. Cytokine Growth Factor Rev. 2019, 48, 32–39. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, Y. Therapy Insight: Adipocytokines in metabolic syndrome and related cardiovascular disease. Nat. Clin. Pract. Cardiovasc. Med. 2006, 3, 35–42. [Google Scholar] [CrossRef]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes Metab. Syndr. Obes. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Osborn, O.; Olefsky, J.M. The cellular and signaling networks linking the immune system and metabolism in disease. Nat. Med. 2012, 18, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Lim, S. Ectopic fat assessment focusing on cardiometabolic and renal risk. Endocrinol. Metab. 2014, 29, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.E.; Kim, J.M.; Joung, K.H.; Lee, J.H.; You, B.R.; Choi, M.J.; Ryu, M.J.; Ko, Y.B.; Lee, M.A.; Lee, J.; et al. The Roles of Adipokines, Proinflammatory Cytokines, and Adipose Tissue Macrophages in Obesity-Associated Insulin Resistance in Modest Obesity and Early Metabolic Dysfunction. PLoS ONE 2016, 11, e0154003. [Google Scholar] [CrossRef] [PubMed]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2019, 10, 1638. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Jarisarapurin, W.; Sanrattana, W.; Chularojmontri, L.; Kunchana, K.; Wattanapitayakul, S.K. A ntioxidant Properties of Unripe Carica papaya Fruit Extract and Its Protective Effects against Endothelial Oxidative Stress. Evid Based Complement Alternat. Med. 2019, 2019, 4912631. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Frohnert, B.I.; Sinaiko, A.R.; Serrot, F.J.; Foncea, R.E.; Moran, A.; Ikramuddin, S.; Choudry, U.; Bernlohr, D.A. Increased adipose protein carbonylation in human obesity. Obesity 2011, 19, 1735–1741. [Google Scholar] [CrossRef]
- Asghar, N.; Naqvi, S.A.; Hussain, Z.; Rasool, N.; Khan, Z.A.; Shahzad, S.A.; Sherazi, T.A.; Janjua, M.R.; Nagra, S.A.; Zia-Ul-Haq, M.; et al. Compositional difference in antioxidant and antibacterial activity of all parts of the Carica papaya using different solvents. Chem. Cent. J. 2016, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Stanislas, G.; Douraguia, E.; Gonthier, M.P. Evaluation of nutritional and antioxidant properties of the tropical fruits banana, litchi, mango, papaya, passion fruit and pineapple cultivated in Réunion French Island. Food Chem. 2016, 212, 225–233. [Google Scholar] [CrossRef]
- Udomkun, P.; Nagle, M.; Argyropoulos, D.; Mahayothee, B.; Latif, S.; Müller, J. Compositional and functional dynamics of dried papaya as affected by storage time and packaging material. Food Chem. 2016, 196, 712–719. [Google Scholar] [CrossRef]
- Awodele, O.; Yemitan, O.; Ise, P.U.; Ikumawoyi, V.O. Modulatory potentials of aqueous leaf and unripe fruit extracts of Carica papaya Linn. (Caricaceae) against carbon tetrachloride and acetaminophen-induced hepatotoxicity in rats. J. Intercult. Ethnopharmacol. 2016, 5, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Salla, S.; Sunkara, R.; Ogutu, S.; Walker, L.T.; Verghese, M. Antioxidant activity of papaya seed extracts against H2O2 induced oxidative stress in HepG2 cells. LWT Food Sci. Technol. 2016, 66, 293–297. [Google Scholar] [CrossRef]
- Spampinato, S.F.; Caruso, G.I.; De Pasquale, R.; Sortino, M.A.; Merlo, S. The Treatment of Impaired Wound Healing in Diabetes: Looking among Old Drugs. Pharmaceuticals 2020, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.; Elder, S.; Veves, A. Delayed wound healing in diabetes: Considering future treatments. Diabetes Manag. 2011, 1, 509–519. [Google Scholar] [CrossRef]
- Gurung, S.; Skalko-Basnet, N. Wound healing properties of Carica papaya latex: In vivo evaluation in mice burn model. J. Ethnopharmacol. 2009, 121, 338–341. [Google Scholar] [CrossRef]
- Nayak, B.S.; Pinto Pereira, L.M. Catharanthus roseus flower extract has wound-healing activity in Sprague Dawley rats. BMC Complement. Altern. Med. 2006, 6, 41. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fresh Papaya | |
|---|---|
| Green papaya (g) | 1171.48 ± 15.38 |
| Skin (%) | 3.58 ± 0.76 |
| Pulp (%) | 90.85 ± 2.12 |
| Seeds (%) | 5.57 ± 1.89 |
| Yellow papaya (g) | 1854.15 ± 24.53 |
| Skin (%) | 3.83 ± 0.50 |
| Pulp (%) | 91.12 ± 2.68 |
| Seeds (%) | 5.05 ± 2.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Beg, O.U.; Rafie, A.R.; Kanwal, S.; Ovalle-Cisneros, A.; Faison, M.O.; Siddiqui, R.A. Characterization of Green and Yellow Papaya (Carica papaya) for Anti-Diabetic Activity in Liver and Myoblast Cells and Wound-Healing Activity in Fibroblast Cells. Nutrients 2023, 15, 1929. https://doi.org/10.3390/nu15081929
Li H, Beg OU, Rafie AR, Kanwal S, Ovalle-Cisneros A, Faison MO, Siddiqui RA. Characterization of Green and Yellow Papaya (Carica papaya) for Anti-Diabetic Activity in Liver and Myoblast Cells and Wound-Healing Activity in Fibroblast Cells. Nutrients. 2023; 15(8):1929. https://doi.org/10.3390/nu15081929
Chicago/Turabian StyleLi, Haiwen, Obaid Ullah Beg, Ahmed Reza Rafie, Sadia Kanwal, Alexandra Ovalle-Cisneros, Milton Omar Faison, and Rafat Ali Siddiqui. 2023. "Characterization of Green and Yellow Papaya (Carica papaya) for Anti-Diabetic Activity in Liver and Myoblast Cells and Wound-Healing Activity in Fibroblast Cells" Nutrients 15, no. 8: 1929. https://doi.org/10.3390/nu15081929