
Amelioration of Sensorineural Hearing Loss through Regulation of Trpv1, Cacna1h, and Ngf Gene Expression by a Combination of Cuscutae Semen and Rehmanniae Radix Preparata
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of TS
2.2. House Ear Institute-Organ of Corti 1 (HEI-OC1) Cells
2.3. MTT Assay
2.4. Zebrafish
2.5. Neomycin-Induced Ototoxicity in Zebrafish
2.6. Scanning Electron Microscopy (SEM)
2.7. Mouse
2.8. Auditory Brainstem Response (ABR)
2.9. Extraction of Total RNA
2.10. Quantitative RT-PCR
2.11. Statistical Analysis
3. Results
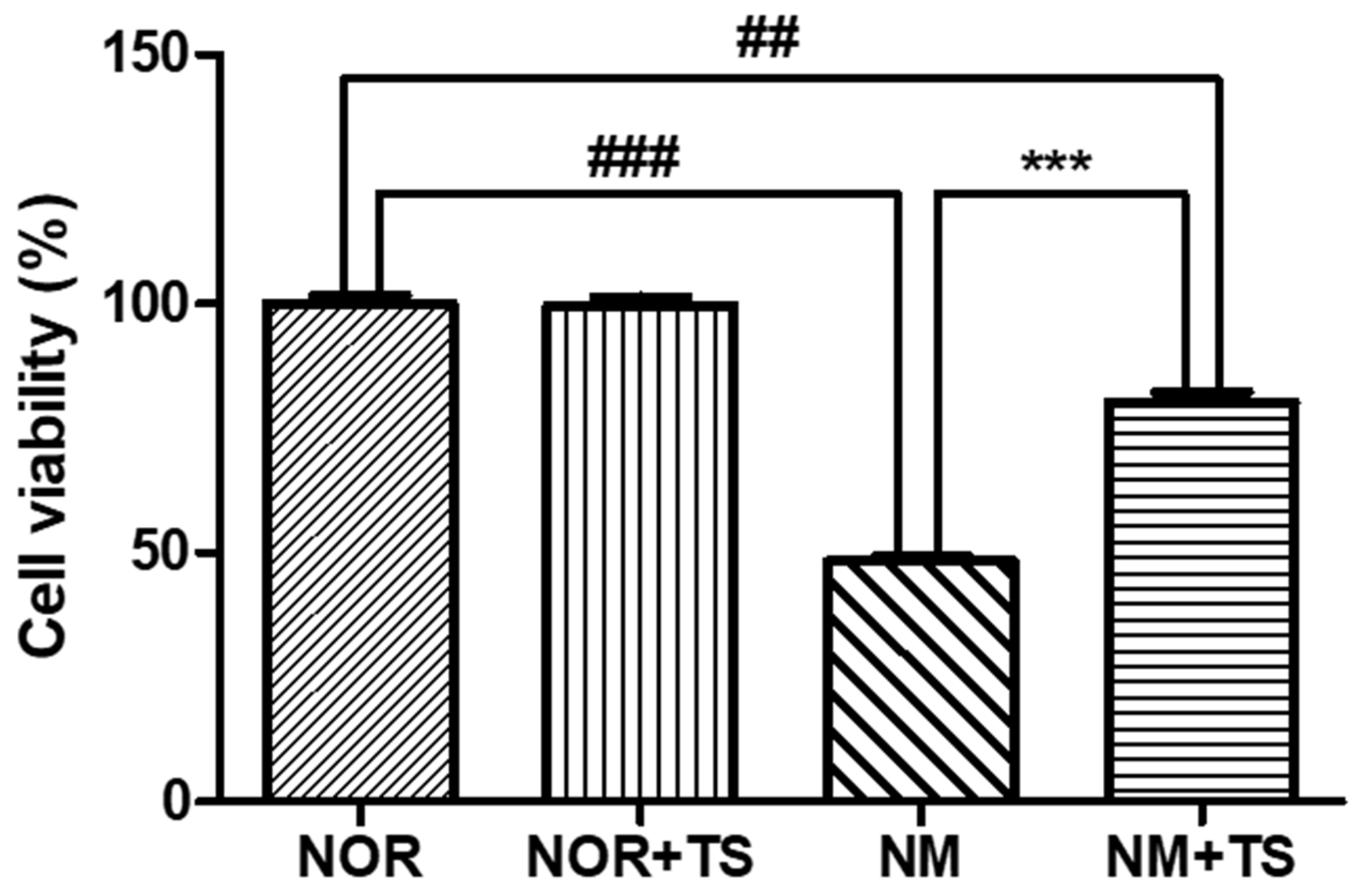
3.1. Effect of TS on Cell Viability in Neomycin-Treated HEI-OC1 Cells
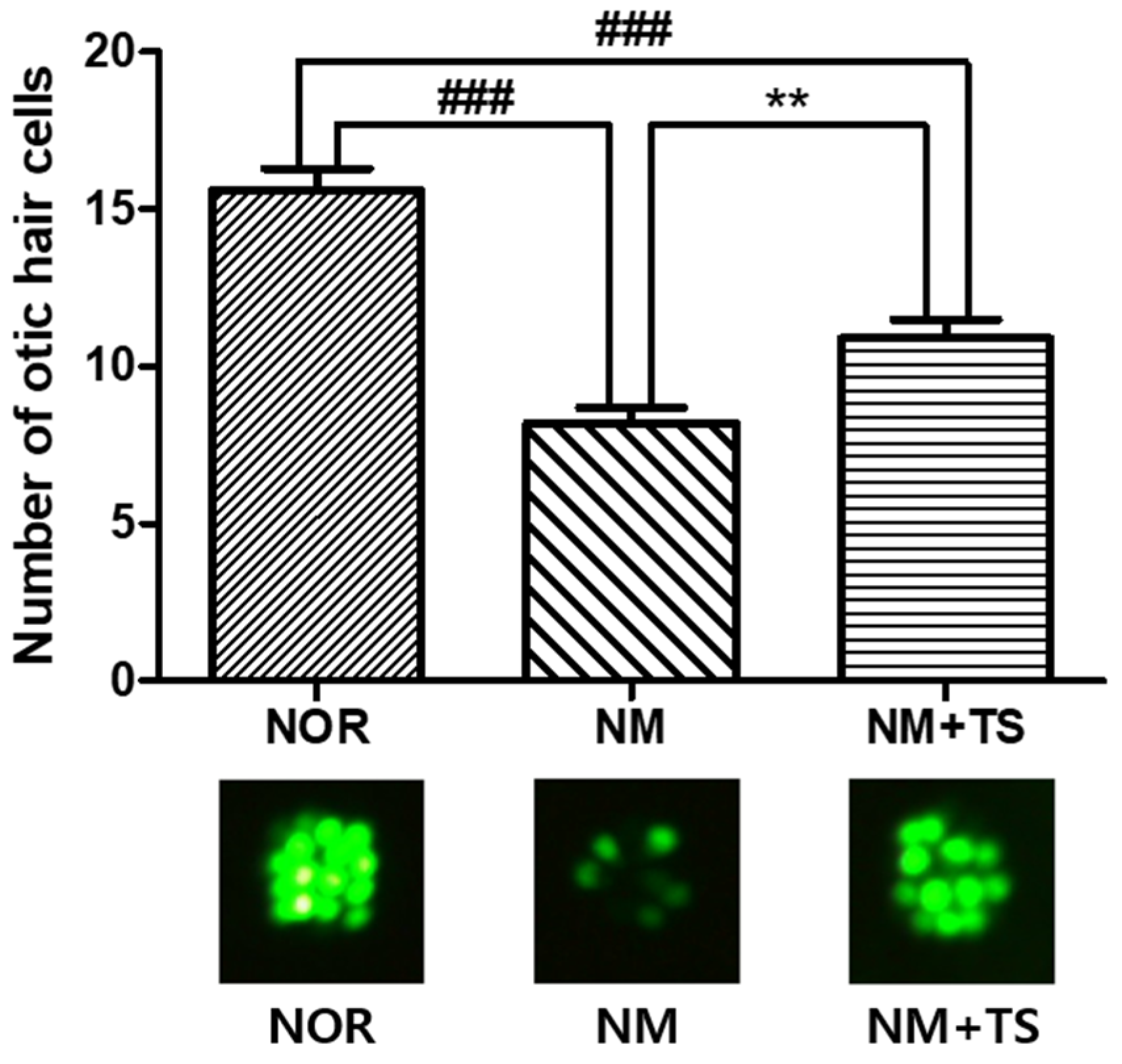
3.2. Effect of TS on Otic Hair Cells in Neomycin-Treated Zebrafish
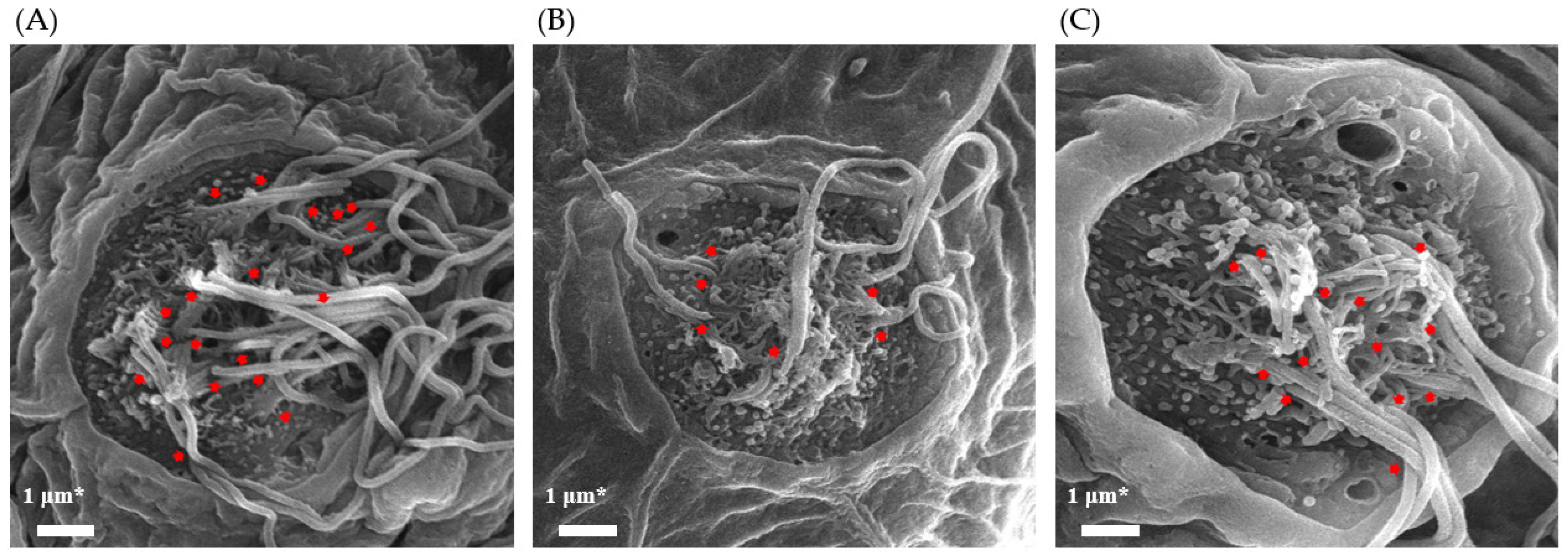
3.3. Effect of TS on Cilia Bundle in Neomycin-Treated Zebrafish
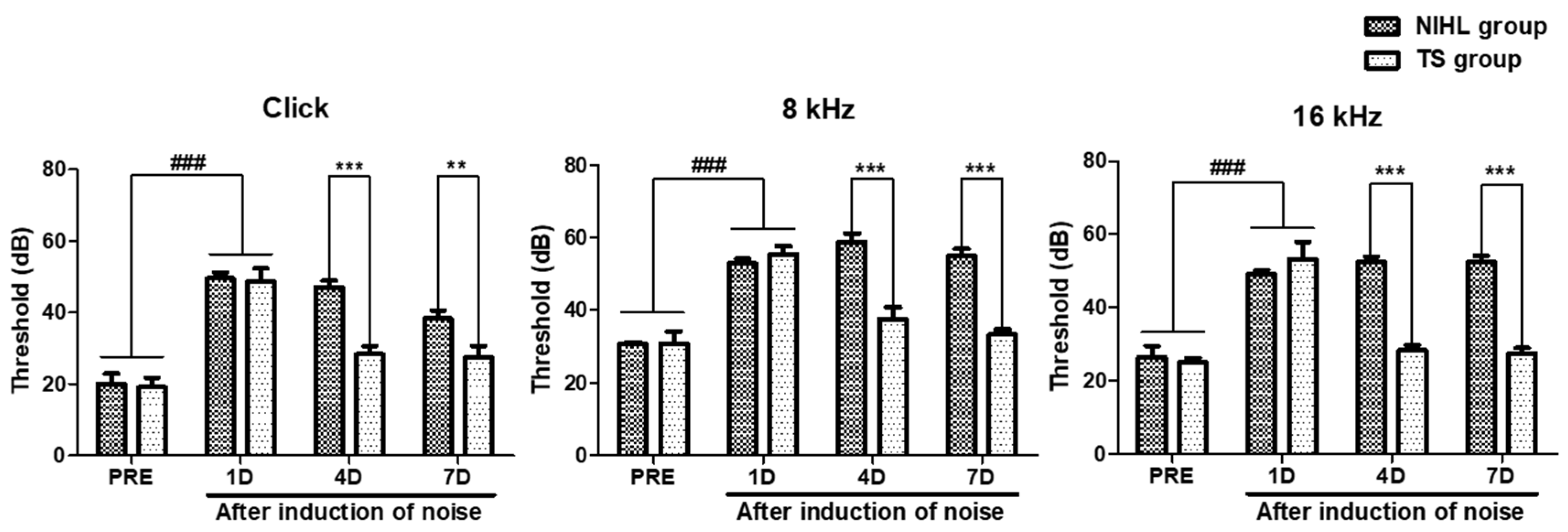
3.4. Effect of TS on Hearing Threshold in Mice with Noise-Induced Hearing Loss
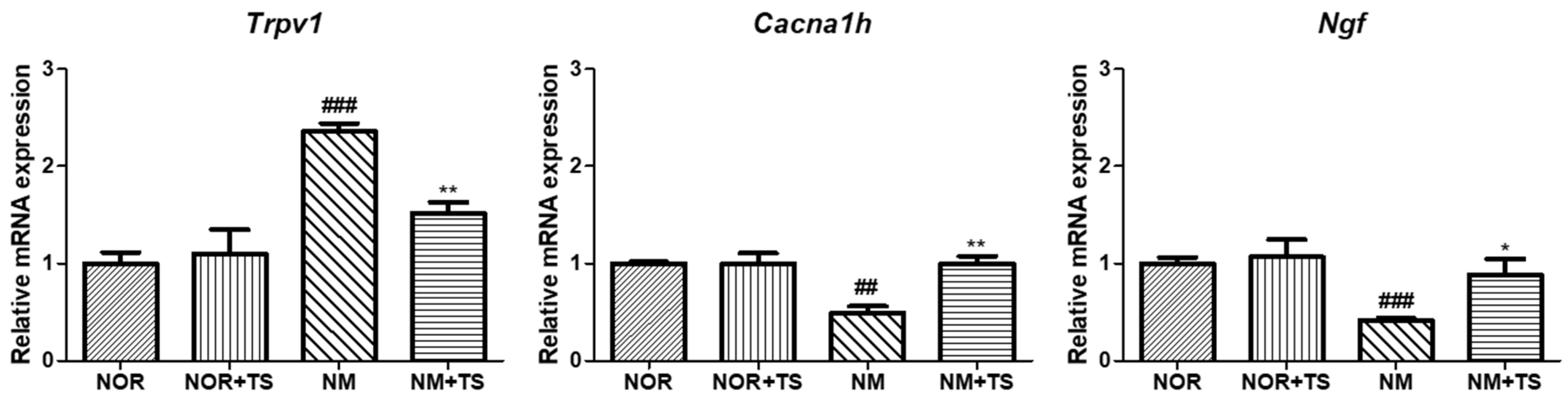
3.5. Effect of TS on Gene-Expression-Related Hearing Loss in Neomycin-Treated HEI-OC1 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuhn, M.; Heman-Ackah, S.E.; Shaikh, J.A.; Roehm, P.C. Sudden sensorineural hearing loss: A review of diagnosis, treatment, and prognosis. Trends Amplif. 2011, 15, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Choi, H.G.; Jang, J.H.; Sim, S.; Hong, S.K.; Lee, H.-J.; Park, B.; Kim, H.-J. Analysis of Predisposing Factors for Hearing Loss in Adults. J. Korean Med. Sci. 2015, 30, 1175–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umashankar, A.; Prakash, P.; Prabhu, P. Sudden Sensorineural Hearing Loss Post Coronavirus Disease: A Systematic Review of Case Reports. Indian J. Otolaryngol. Head Neck Surg. 2022, 74, 3028–3035. [Google Scholar] [CrossRef] [PubMed]
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Hearing Loss, Tinnitus, and Dizziness in COVID-19: A Systematic Review and Meta-Analysis. Can. J. Neurol. Sci. 2022, 49, 184–195. [Google Scholar] [CrossRef]
- Vlajkovic, S.M.; Thorne, P.R. Molecular Mechanisms of Sensorineural Hearing Loss and Development of Inner Ear Therapeutics. Int. J. Mol. Sci. 2021, 22, 5647. [Google Scholar] [CrossRef]
- Koeberle, A.; Werz, O. Multi-target approach for natural products in inflammation. Drug Discov. Today 2014, 19, 1871–1882. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [Green Version]
- Park, I.; Yang, S.; Choi, G.; Moon, B.C.; Song, J.H. An Integrated Approach for Efficient and Accurate Medicinal Cuscutae Semen Identification. Plants 2020, 9, 1410. [Google Scholar] [CrossRef]
- Ye, M.; Li, Y.; Yan, Y.; Liu, H.; Ji, X. Determination of flavonoids in Semen Cuscutae by RP-HPLC. J. Pharm. Biomed. Anal. 2002, 28, 621–628. [Google Scholar] [CrossRef]
- Hong, S.P.; Kim, Y.C.; Kim, K.H.; Park, J.H.; Park, M.K. Characteristic component of Rehmanniae Radix Preparata compared to Rehmanniae Radix and Rehmanniae Radix Crudus. Anal. Sci. Technol. 1993, 6, 401–404. [Google Scholar]
- Youn, U.J.; Gu, B.S.; Kim, K.H.; Ha, C.; Jung, I.C. Variation of main components according to the number of steaming and drying of Rehmanniae radix preparata. J. Pharmacopunct. 2018, 21, 112–119. [Google Scholar] [CrossRef]
- Yu, H.H.; Kim, Y.H.; Jung, S.Y.; Shin, M.K.; Park, R.K.; So, H.S.; Kim, K.Y.; Lee, D.H.; You, Y.O. Rehmannia glutinosa activates intracellular antioxidant enzyme systems in mouse auditory cells. Am. J. Chin. Med. 2006, 34, 1083–1093. [Google Scholar] [CrossRef]
- Yu, H.H.; Seo, S.J.; Kim, Y.H. Protective effect of Rehmannia glutinosa on the cisplatin-induced damage of HEI-OC1 auditory cells through scavenging free radicals. J. Ethnopharmacol. 2006, 107, 383–388. [Google Scholar] [CrossRef]
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Axelsson, A.; Ringdahl, A. Tinnitus—A study of its prevalence and characteristics. Br. J. Audiol. 1989, 23, 53–62. [Google Scholar] [CrossRef]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Myrdal, S.E.; Steyger, P.S. TRPV1 regulators mediate gentamicin penetration of cultured kidney cells. Hear. Res. 2005, 204, 170–182. [Google Scholar] [CrossRef] [Green Version]
- Mukherjea, D.; Jajoo, S.; Sheehan, K.; Kaur, T.; Sheth, S.; Bunch, J.; Perro, C.; Rybak, L.P.; Ramkumar, V. NOX3 NADPH oxidase couples transient receptor potential vanilloid 1 to signal transducer and activator of transcription 1-mediated inflammation and hearing loss. Antioxid. Redox Signal. 2011, 14, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Dai, C.; Steyger, P.S.; Kim, Y.; Vass, Z.; Ren, T.; Nuttall, A.L. Vanilloid receptors in hearing: Altered cochlear sensitivity by vanilloids and expression of TRPV1 in the organ of corti. J. Neurophysiol. 2003, 90, 444–455. [Google Scholar] [CrossRef]
- Jiang, M.; Li, H.; Johnson, A.; Karasawa, T.; Zhang, Y.; Meier, W.B.; Taghizadeh, F.; Kachelmeier, A.; Steyger, P.S. Inflammation up-regulates cochlear expression of TRPV1 to potentiate drug-induced hearing loss. Sci. Adv. 2019, 5, eaaw1836. [Google Scholar] [CrossRef] [Green Version]
- Storozhuk, M.V.; Moroz, O.F.; Zholos, A.V. Multifunctional TRPV1 Ion Channels in Physiology and Pathology with Focus on the Brain, Vasculature, and Some Visceral Systems. BioMed Res. Int. 2019, 2019, 5806321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramkumar, V.; Sheth, S.; Dhukhwa, A.; Al Aameri, R.; Rybak, L.; Mukherjea, D. Transient Receptor Potential Channels and Auditory Functions. Antioxid. Redox Signal. 2022, 36, 1158–1170. [Google Scholar] [CrossRef] [PubMed]
- Fridberger, A.; Flock, A.; Ulfendahl, M.; Flock, B. Acoustic overstimulation increases outer hair cell Ca2+ concentrations and causes dynamic contractions of the hearing organ. Proc. Natl. Acad. Sci. USA 1998, 95, 7127–7132. [Google Scholar] [CrossRef] [Green Version]
- Mann, W.; Pilgramm, M.; Löhle, E.; Beck, C. Calcium antagonists and damage to the organ of Corti in acoustic trauma. HNO 1987, 35, 203–207. [Google Scholar] [PubMed]
- Esterberg, R.; Hailey, D.W.; Coffin, A.B.; Raible, D.W.; Rubel, E.W. Disruption of intracellular calcium regulation is integral to aminoglycoside-induced hair cell death. J. Neurosci. 2013, 33, 7513–7525. [Google Scholar] [CrossRef] [Green Version]
- Orrenius, S.; McCabe, M.J., Jr.; Nicotera, P. Ca2+-dependent mechanisms of cytotoxicity and programmed cell death. Toxicol. Lett. 1992, 64–65, 357–364. [Google Scholar] [CrossRef]
- Triggle, D.J. The pharmacology of ion channels: With particular reference to voltage-gated Ca2+ channels. Eur. J. Pharmacol. 1999, 375, 311–325. [Google Scholar] [CrossRef]
- Lundt, A.; Seidel, R.; Soós, J.; Henseler, C.; Müller, R.; Bakki, M.; Arshaad, M.I.; Ehninger, D.; Hescheler, J.; Sachinidis, A.; et al. Cav3.2 T-Type Calcium Channels Are Physiologically Mandatory for the Auditory System. Neuroscience 2019, 409, 81–100. [Google Scholar] [CrossRef]
- Comunanza, V.; Carbone, E.; Marcantoni, A.; Sher, E.; Ursu, D. Calcium-dependent inhibition of T-type calcium channels by TRPV1 activation in rat sensory neurons. Pflugers Arch. 2011, 462, 709–722. [Google Scholar] [CrossRef]
- Reijntjes, D.O.J.; Pyott, S.J. The afferent signaling complex: Regulation of type I spiral ganglion neuron responses in the auditory periphery. Hear. Res. 2016, 336, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bernd, P.; Represa, J. Characterization and localization of nerve growth factor receptors in the embryonic otic vesicle and cochleovestibular ganglion. Dev. Biol. 1989, 134, 11–20. [Google Scholar] [CrossRef]
- Lewin, G.R.; Barde, Y.A. Physiology of the neurotrophins. Annu. Rev. Neurosci. 1996, 19, 289–317. [Google Scholar] [CrossRef]
- Wong, A.C.; Ryan, A.F. Mechanisms of sensorineural cell damage, death and survival in the cochlea. Front. Aging Neurosci. 2015, 7, 58. [Google Scholar] [CrossRef]
- Salvinelli, F.; Casale, M.; Greco, F.; Trivelli, M.; Di Peco, V.; Amendola, T.; Antonelli, A.; Stampachiacchiere, B.; Aloe, L. Nerve growth factor serum level is reduced in patients with sensorineural hearing impairment: Possible clinical implications. J. Biol. Regul. Homeost. Agents 2002, 16, 176–180. [Google Scholar]
- Han, Z.; Wang, C.P.; Cong, N.; Gu, Y.Y.; Ma, R.; Chi, F.L. Therapeutic value of nerve growth factor in promoting neural stem cell survival and differentiation and protecting against neuronal hearing loss. Mol. Cell. Biochem. 2017, 428, 149–159. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, H.; Zheng, T.; Wang, W.; Zhang, X.; Wang, A.; Li, B.; Wang, Y.; Zheng, Q. Otoprotective effects of mouse nerve growth factor in DBA/2J mice with early-onset progressive hearing loss. J. Neurosci. Res. 2017, 95, 1937–1950. [Google Scholar] [CrossRef] [Green Version]
- Kalinec, G.M.; Park, C.; Thein, P.; Kalinec, F. Working with Auditory HEI-OC1 Cells. J. Vis. Exp. 2016, 54425. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Nam, Y.H.; Rodriguez, I.; Jeong, S.Y.; Pham, T.N.M.; Nuankaew, W.; Kim, Y.H.; Castañeda, R.; Jeong, S.Y.; Park, M.S.; Lee, K.W.; et al. Avocado Oil Extract Modulates Auditory Hair Cell Function through the Regulation of Amino Acid Biosynthesis Genes. Nutrients 2019, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Fettiplace, R. Hair Cell Transduction, Tuning, and Synaptic Transmission in the Mammalian Cochlea. Compr. Physiol. 2017, 7, 1197–1227. [Google Scholar]
- White, H.J.; Helwany, M.; Biknevicius, A.R.; Peterson, D.C. Anatomy, Head and Neck, Ear Organ of Corti; StatPearls: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Appler, J.M.; Goodrich, L.V. Connecting the ear to the brain: Molecular mechanisms of auditory circuit assembly. Prog. Neurobiol. 2011, 93, 488–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzi, C.; Gilbert, G.; Carn, H.; Garnier, S.; Moore, B.C. Speech perception problems of the hearing impaired reflect inability to use temporal fine structure. Proc. Natl. Acad. Sci. USA 2006, 103, 18866–18869. [Google Scholar] [CrossRef] [Green Version]
- Kalinec, G.; Thein, P.; Park, C.; Kalinec, F. HEI-OC1 cells as a model for investigating drug cytotoxicity. Hear. Res. 2016, 335, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Kalinec, G.M.; Webster, P.; Lim, D.J.; Kalinec, F. A cochlear cell line as an in vitro system for drug ototoxicity screening. Audiol. Neurootol. 2003, 8, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Nadol, J.B., Jr. Hearing loss. N. Engl. J. Med. 1993, 329, 1092–1102. [Google Scholar] [CrossRef]
- Leis, J.A.; Rutka, J.A.; Gold, W.L. Aminoglycoside-induced ototoxicity. Can. Med. Assoc. J. 2015, 187, E52. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Chen, B.; Chen, J.; Li, Y. Protective Effects of Vitamin C against Neomycin-Induced Apoptosis in HEI-OC1 Auditory Cell. Neural Plast. 2022, 1298692. [Google Scholar] [CrossRef]
- Dooley, K.; Zon, L.I. Zebrafish: A model system for the study of human disease. Curr. Opin. Genet. Dev. 2000, 10, 252–256. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Fritzsch, B.; Beisel, K.W. Evolution and development of the vertebrate ear. Brain Res. Bull. 2001, 55, 711–721. [Google Scholar] [CrossRef]
- Lush, M.E.; Piotrowski, T. Sensory hair cell regeneration in the zebrafish lateral line. Dev. Dyn. 2014, 243, 1187–1202. [Google Scholar] [CrossRef] [Green Version]
- Duncan, J.S.; Fritzsch, B. Evolution of sound and balance perception: Innovations that aggregate single hair cells into the ear and transform a gravistatic sensor into the organ of corti. Anat. Rec. 2012, 295, 1760–1774. [Google Scholar] [CrossRef]
- Whitfield, T.T. Zebrafish as a model for hearing and deafness. J. Neurobiol. 2002, 53, 157–171. [Google Scholar] [CrossRef]
- Murakami, S.L.; Cunningham, L.L.; Werner, L.A.; Bauer, E.; Pujol, R.; Raible, D.W.; Rubel, E.W. Developmental differences in susceptibility to neomycin-induced hair cell death in the lateral line neuromasts of zebrafish (Danio rerio). Hear. Res. 2003, 186, 47–56. [Google Scholar] [CrossRef]
- Williams, J.A.; Holder, N. Cell turnover in neuromasts of zebrafish larvae. Hear. Res. 2000, 143, 171–181. [Google Scholar] [CrossRef]
- Matsui, J.I.; Cotanche, D.A. Sensory hair cell death and regeneration: Two halves of the same equation. Curr. Opin. Otolaryngol. Head Neck Surg. 2004, 12, 418–425. [Google Scholar] [CrossRef]
- Monroe, J.D.; Rajadinakaran, G.; Smith, M.E. Sensory hair cell death and regeneration in fishes. Front. Cell. Neurosci. 2015, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Jeong, S.Y.; Nam, Y.H.; Park, J.H.; Rodriguez, I.; Shim, J.H.; Yasmin, T.; Kwak, H.J.; Oh, Y.; Oh, M.; et al. Fatty Acid Derivatives Isolated from the Oil of Persea americana (Avocado) Protects against Neomycin-Induced Hair Cell Damage. Plants 2021, 10, 171. [Google Scholar] [CrossRef]
- Kim, H.G.; Oh, S.M.; Kim, N.W.; Shim, J.H.; Nam, Y.H.; Nguyen, T.N.; Lee, M.H.; Lee, D.Y.; Kang, T.H.; Baek, N.I. Three New Phthalide Glycosides from the Rhizomes of Cnidium officinale and Their Recovery Effect on Damaged Otic Hair Cells in Zebrafish. Molecules 2021, 26, 7034. [Google Scholar] [CrossRef]
- Harris, J.A.; Cheng, A.G.; Cunningham, L.L.; MacDonald, G.; Raible, D.W.; Rubel, E.W. Neomycin-induced hair cell death and rapid regeneration in the lateral line of zebrafish (Danio rerio). J. Assoc. Res. Otolaryngol. 2003, 4, 219–234. [Google Scholar] [CrossRef]
- Wang, D.; Zhou, J. The Kinocilia of Cochlear Hair Cells: Structures, Functions, and Diseases. Front. Cell Dev. Biol. 2021, 9, 715037. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, J.J. Auditory brainstem response. Handb. Clin. Neurol. 2019, 160, 451–464. [Google Scholar] [PubMed]
- Heffner, H.E.; Heffner, R.S. Hearing ranges of laboratory animals. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 20–22. [Google Scholar] [PubMed]
- Girotto, G.; Vuckovic, D.; Buniello, A.; Lorente-Cánovas, B.; Lewis, M.; Gasparini, P.; Steel, K.P. Expression and replication studies to identify new candidate genes involved in normal hearing function. PLoS ONE 2014, 9, e85352. [Google Scholar] [CrossRef]
- Ohlemiller, K.K. Mouse methods and models for studies in hearing. J. Acoust. Soc. Am. 2019, 146, 3668. [Google Scholar] [CrossRef] [Green Version]
- Hong, B.N.; Park, T.G.; Hong, H.N.; Kang, T.H. A Validation Study of Auditory Brainstem Response (ABR) in ICR Mouse. Audiol. Speech Res. 2008, 4, 58–63. [Google Scholar] [CrossRef]
- Henderson, D.; Hamernik, R.P. Biologic bases of noise-induced hearing loss. Occup. Med. 1995, 10, 513–534. [Google Scholar]
- Park, S.N.; Back, S.A.; Park, K.H.; Seo, J.H.; Noh, H.I.; Akil, O.; Lustig, L.R.; Yeo, S.W. Comparison of functional and morphologic characteristics of mice models of noise-induced hearing loss. Auris Nasus Larynx 2013, 40, 11–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′ to 3′) | NCBI Sequence |
|---|---|---|---|
| Trpv1 | Forward | GGAAGACAGATAGCCTGAAG | NM_001001445.2 |
| Reverse | GAGAATGTAGGCCAAGACC | ||
| Cacna1h | Forward | GCTCTACTTCATCTCCTTCC | NM_021415.4 |
| Reverse | CTGTGGCCATCTTCAGTAG | ||
| Ngf | Forward | TGAAGCCCACTGGACTAA | NM_001112698.2 |
| Reverse | GTCTATCCGGATGAACCTC | ||
| β-actin | Forward | GAAGAGCTATGAGCTGCCTGA | NM_007393.5 |
| Reverse | TGATCCACATCTGCTGGAAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, B.N.; Shin, S.W.; Nam, Y.H.; Shim, J.H.; Kim, N.W.; Kim, M.C.; Nuankaew, W.; Kwak, J.H.; Kang, T.H. Amelioration of Sensorineural Hearing Loss through Regulation of Trpv1, Cacna1h, and Ngf Gene Expression by a Combination of Cuscutae Semen and Rehmanniae Radix Preparata. Nutrients 2023, 15, 1773. https://doi.org/10.3390/nu15071773
Hong BN, Shin SW, Nam YH, Shim JH, Kim NW, Kim MC, Nuankaew W, Kwak JH, Kang TH. Amelioration of Sensorineural Hearing Loss through Regulation of Trpv1, Cacna1h, and Ngf Gene Expression by a Combination of Cuscutae Semen and Rehmanniae Radix Preparata. Nutrients. 2023; 15(7):1773. https://doi.org/10.3390/nu15071773
Chicago/Turabian StyleHong, Bin Na, Sung Woo Shin, Youn Hee Nam, Ji Heon Shim, Na Woo Kim, Min Cheol Kim, Wanlapa Nuankaew, Jong Hwan Kwak, and Tong Ho Kang. 2023. "Amelioration of Sensorineural Hearing Loss through Regulation of Trpv1, Cacna1h, and Ngf Gene Expression by a Combination of Cuscutae Semen and Rehmanniae Radix Preparata" Nutrients 15, no. 7: 1773. https://doi.org/10.3390/nu15071773