

Improved Strength Recovery and Reduced Fatigue with Suppressed Plasma Myostatin Following Supplementation of a Vicia faba Hydrolysate, in a Healthy Male Population

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Trial Design
2.3. Ingredient Production and Supplementation
2.4. Strength Measurements
2.5. Exhaustive Exercise Test (EET)
2.6. Fatigue Index (FI)
2.7. Myokine Array
2.8. Data Analysis and Statistics
3. Results
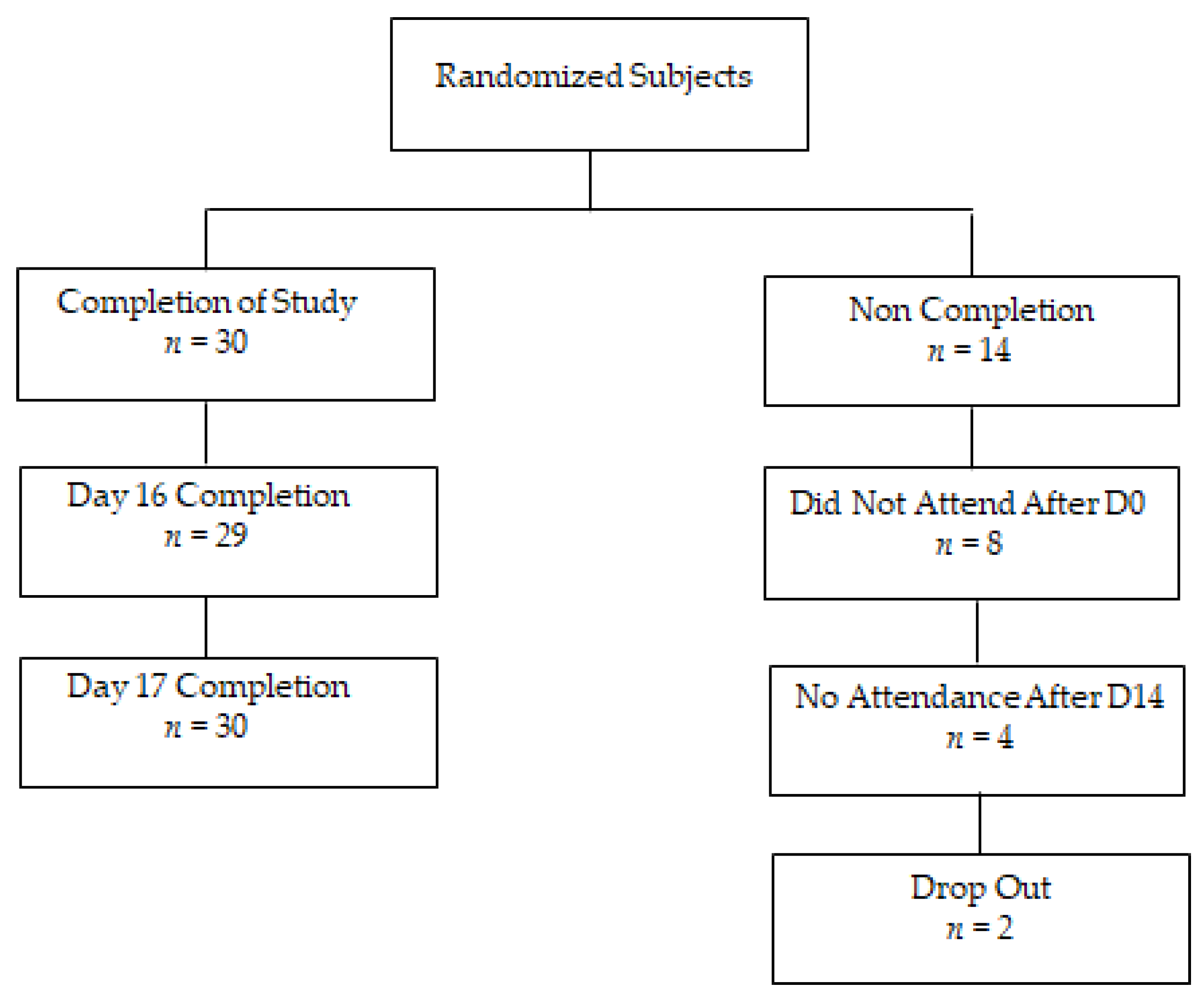
3.1. Trial Design
3.2. Strength Recovery
3.3. Fatigue Index
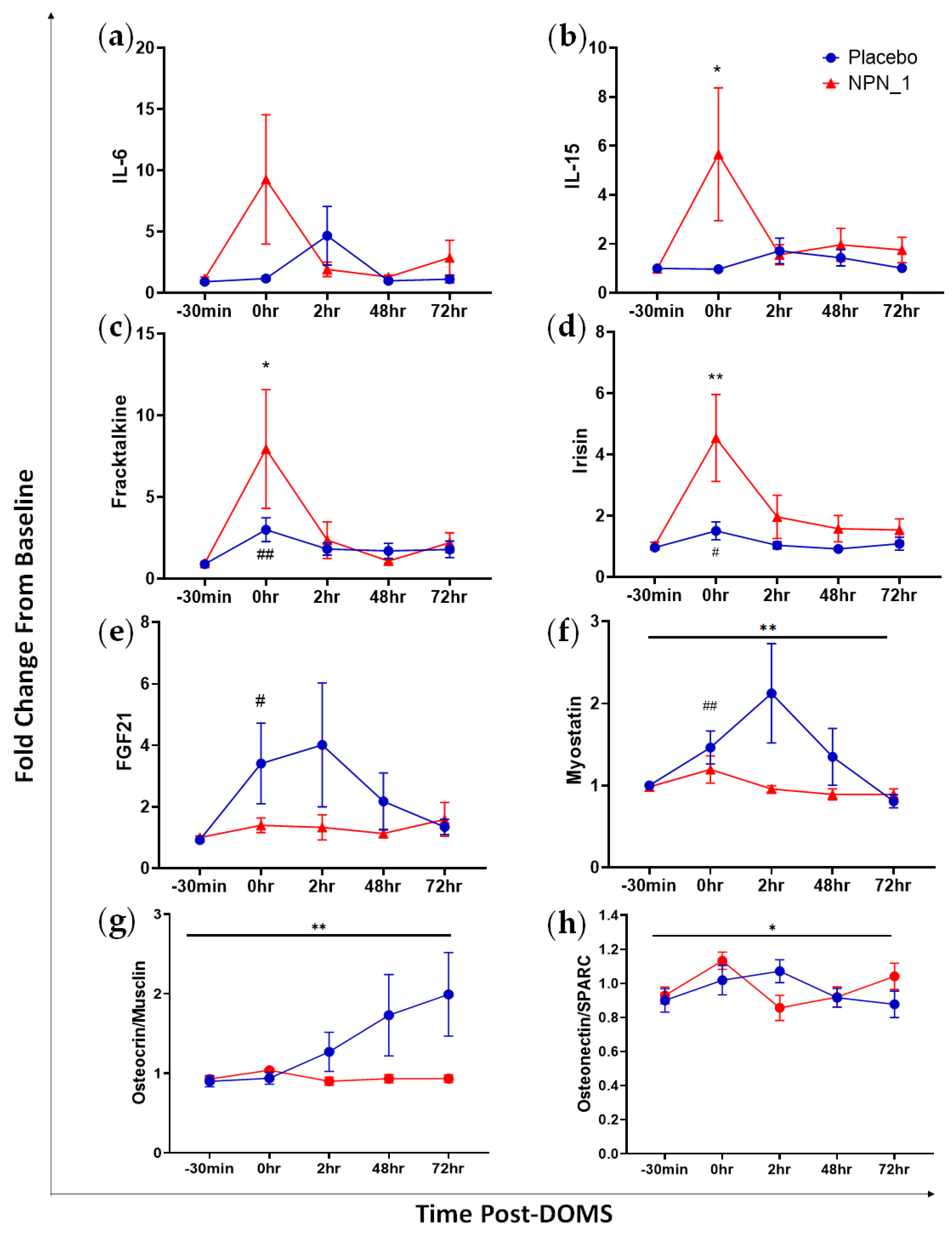
3.4. Molecular Markers of Muscle Recovery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Twist, C.; Eston, R. The effets of exercise-induced muscle damage on maximal intensity intermittent exercise performance. Eur. J. Appl. Physiol. 2005, 94, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.; Hume, P.A.; Maxwell, L. Delayed Onset Muscle Soreness. Sport. Med. 2003, 33, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, P.M.; Hubal, M.J. Exercise-induced muscle damage in humans. Am. J. Phys. Med. Rehabil. 2002, 81, S52–S69. [Google Scholar] [CrossRef] [PubMed]
- Ranchordas, M.K.; Rogerson, D.; Soltani, H.; Costello, J.T. Antioxidants for preventing and reducing muscle soreness after exercise: A Cochrane systematic review. Br. J. Sport. Med. 2020, 54, 74–78. [Google Scholar] [CrossRef]
- Cleak, M.; Eston, R. Stiffness and Strength Loss After Intense Eccentric Exercise. Br. J. Sport. Med. 1992, 26, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Bongiovanni, T.; Genovesi, F.; Nemmer, M.; Carling, C.; Alberti, G.; Howatson, G. Nutritional interventions for reducing the signs and symptoms of exercise-induced muscle damage and accelerate recovery in athletes: Current knowledge, practical application and future perspectives. Eur. J. Appl. Physiol. 2020, 120, 1965–1996. [Google Scholar] [CrossRef]
- Nabuco, H.C.G.; Tomeleri, C.M.; Fernandes, R.R.; Sugihara Junior, P., Jr.; Cavalcante, E.F.; Cunha, P.M.; Antunes, M.; Nunes, J.P.; Venturini, D.; Barbosa, D.S.; et al. Effect of whey protein supplementation combined with resistance training on body composition, muscular strength, functional capacity, and plasma-metabolism biomarkers in older women with sarcopenic obesity: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. ESPEN 2019, 32, 88–95. [Google Scholar] [CrossRef]
- Ramos-Campo, D.J.; Ávila-Gandía, V.; López-Román, F.J.; Miñarro, J.; Contreras, C.; Soto-Méndez, F.; Pedrol, J.C.D.; Luque-Rubia, A.J. Supplementation of Re-Esterified Docosahexaenoic and Eicosapentaenoic Acids Reduce Inflammatory and Muscle Damage Markers after Exercise in Endurance Athletes: A Randomized, Controlled Crossover Trial. Nutrients 2020, 12, 719. [Google Scholar] [CrossRef] [Green Version]
- Buchwald-Werner, S.; Naka, I.; Wilhelm, M.; Schütz, E.; Schoen, C.; Reule, C. Effects of lemon verbena extract (Recoverben®) supplementation on muscle strength and recovery after exhaustive exercise: A randomized, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2018, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bell, P.G.; Walshe, I.H.; Davison, G.W.; Stevenson, E.J.; Howatson, G. Recovery facilitation with montmorency cherries following high-intensity, metabolically challenging exercise. Appl. Physiol. Nutr. Metab. 2015, 40, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Veggi, K.F.T.; Machado, M.; Koch, A.J.; Santana, S.C.; Oliveira, S.S.; Stec, M.J. Oral Creatine Supplementation Augments the Repeated Bout Effect. 2013. Available online: www.IJSNEM-Journal.com (accessed on 23 January 2013).
- Machin, D.R.; Christmas, K.M.; Chou, T.-H.; Hill, S.C.; Van Pelt, D.W.; Trombold, J.R.; Coyle, E.F. Effects of Differing Dosages of Pomegranate Juice Supplementation after Eccentric Exercise. Physiol. J. 2014, 2014, 271959. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Forough, J.; Wu, J.; Jahandideh, F.; Wu, J. Food-derived bioactive peptides on inflammation and oxidative stress. Biomed. Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothman, S. How is the balance between protein synthesis and degradation achieved? Theor. Biol. Med. Model. 2010, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, A.; Wall, A.; Khaldi, N.; Kussmann, M. Artificial Intelligence in Functional Food Ingredient Discovery and Characterisation: A Focus on Bioactive Plant and Food Peptides. Front. Genet. 2021, 12, 768979. [Google Scholar] [CrossRef]
- Kennedy, K.; Keogh, B.; Lopez, C.; Adelfio, A.; Molloy, B.; Kerr, A.; Wall, A.M.; Jalowicki, G.; Holton, T.A.; Khaldi, N. An Artificial Intelligence Characterised Functional Ingredient, Derived from Rice, Inhibits TNF-α and Significantly Improves Physical Strength in an Inflammaging Population. Foods 2020, 9, 1147. [Google Scholar] [CrossRef]
- Chauhan, S.; Kerr, A.; Keogh, B.; Nolan, S.; Casey, R.; Adelfio, A.; Murphy, N.; Doherty, A.; Davis, H.; Wall, A.; et al. An Artificial-Intelligence-Discovered Functional Ingredient, NRT_N0G5IJ, Derived from Pisum sativum, Decreases HbA1c in a Prediabetic Population. Nutrients 2021, 13, 1635. [Google Scholar] [CrossRef]
- Corrochano, A.R.; Cal, R.; Kennedy, K.; Wall, A.; Murphy, N.; Trajkovic, S.; O’Callaghan, S.; Adelfio, A.; Khaldi, N. Characterising the efficacy and bioavailability of bioactive peptides identified for attenuating muscle atrophy within a Vicia faba-derived functional ingredient. Curr. Res. Food Sci. 2021, 4, 224–232. [Google Scholar] [CrossRef]
- Cal, R.; Davis, H.; Kerr, A.; Wall, A.; Molloy, B.; Chauhan, S.; Trajkovic, S.; Holyer, I.; Adelfio, A.; Khaldi, N. Preclinical Evaluation of a Food-Derived Functional Ingredient to Address Skeletal Muscle Atrophy. Nutrients 2020, 12, 2274. [Google Scholar] [CrossRef]
- Feiring, D.C.; Ellenbecker, T.S.; Derscheid, G.L. Test-retest reliability of the biodex isokinetic dynamometer. J. Orthop. Sports Phys. Ther. 1990, 11, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Drouin, J.M.; Valovich-mcLeod, T.C.; Shultz, S.J.; Gansneder, B.M.; Perrin, D.H. Reliability and validity of the Biodex system 3 pro isokinetic dynamometer velocity, torque and position measurements. Eur. J. Appl. Physiol. 2004, 91, 22–29. [Google Scholar] [CrossRef]
- Baumgart, C.; Welling, W.; Hoppe, M.W.; Freiwald, J.; Gokeler, A. Angle-specific analysis of isokinetic quadriceps and hamstring torques and ratios in patients after ACL-reconstruction. BMC Sports Sci. Med. Rehabil. 2018, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Malley, E.; Richter, C.; King, E.; Strike, S.; Moran, K.; Franklyn-Miller, A.; Moran, R. Countermovement Jump and Isokinetic Dynamometry as Measures of Rehabilitation Status After Anterior Cruciate Ligament Reconstruction. J. Athl. Train. 2018, 53, 687–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccone, A.B.; Deckert, J.A.; Herda, T.J.; Gallagher, P.M.; Weir, J.P. Methodological Differences in the Interpretation of Fatigue Data from Repeated Maximal Effort Knee Extensions. Open Sports Sci. J. 2017, 10, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Klingel, S.L.; Metherel, A.H.; Irfan, M.; Rajna, A.; Chabowski, A.; Bazinet, R.P.; Mutch, D.M. EPA and DHA have divergent effects on serum triglycerides and lipogenesis, but similar effects on lipoprotein lipase activity: A randomized controlled trial. Am. J. Clin. Nutr. 2019, 110, 1502–1509. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, O.; Douzi, W.; Theurot, D.; Bosquet, L.; Dugué, B. An evidence-based approach for choosing post-exercise recovery techniques to reduce markers of muscle damage, Soreness, fatigue, and inflammation: A systematic review with meta-analysis. Front. Physiol. 2018, 9, 403. [Google Scholar] [CrossRef]
- Baumert, P.; Temple, S.; Stanley, J.M.; Cocks, M.; Strauss, J.A.; Shepherd, S.O.; Drust, B.; Lake, M.J.; Stewart, C.E.; Erskine, R.M. Neuromuscular fatigue and recovery after strenuous exercise depends on skeletal muscle size and stem cell characteristics. Sci. Rep. 2021, 11, 7733. [Google Scholar] [CrossRef] [PubMed]
- Beals, K.; Allison, K.F.; Darnell, M.; Lovalekar, M.; Baker, R.; Nieman, D.C.; Vodovotz, Y.; Lephart, S.M. The effects of a tart cherry beverage on reducing exercise-induced muscle soreness. Isokinet. Exerc. Sci. 2017, 25, 53–63. [Google Scholar] [CrossRef]
- Bell, P.G.; Stevenson, E.; Davison, G.W.; Howatson, G. The effects of montmorency tart cherry concentrate supplementation on recovery following prolonged, intermittent exercise. Nutrients 2016, 8, 441. [Google Scholar] [CrossRef] [Green Version]
- VanDusseldorp, T.A.; Escobar, K.A.; Johnson, K.E.; Stratton, M.T.; Moriarty, T.; Cole, N.; McCormick, J.J.; Kerksick, C.M.; Vaughan, R.A.; Dokladny, K.; et al. Effect of branched-chain amino acid supplementation on recovery following acute eccentric exercise. Nutrients 2018, 10, 1389. [Google Scholar] [CrossRef] [Green Version]
- McGlory, C.; Devries, M.C.; Phillips, S.M. Skeletal muscle and resistance exercise training; the role of protein synthesis in recovery and remodeling. J. Appl. Physiol. 2017, 122, 541–548. [Google Scholar] [CrossRef]
- Carraro, F.; Stuart, C.A.; Hartl, W.H.; Rosenblatt, J.; Wolfe, R.R. Effect of exercise and recovery on muscle protein synthesis in human subjects. Am. J. Physiol. Endocrinol. Metab. 1990, 259, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Weijzen, M.E.; Holwerda, A.M.; Jetten, G.H.; Houben, L.H.; Kerr, A.; Davis, H.; Keogh, B.; Khaldi, N.; Verdijk, L.B.; van Loon, L.J. Vicia faba Peptide Network Supplementation Does Not Differ From Milk Protein in Modulating Changes in Muscle Size During Short-Term Immobilization and Subsequent Remobilization but Increases Muscle Protein Synthesis Rates During Remobilization in Healthy Young Men. J. Nutr. 2023; in press. [Google Scholar] [CrossRef]
- Wall, B.T.; Dirks, M.L.; Snijders, T.; van Dijk, J.-W.; Fritisch, M.; Verdijk, L.B.; van Loon, L.J.C. Short-term muscle disuse lowers myofibrillar protein synthesis rates and induces anabolic resistance to protein ingestion. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E137–E147. [Google Scholar] [CrossRef] [PubMed]
- Pinckaers, P.J.M.; Hendriks, F.K.; Hermans, W.J.; Goessens, J.P.; Senden, J.M.; Van Kranenburg, J.M.X.; Wodzig, W.K.H.W.; Snijders, T.; van Loon, L.J.C. Potato Protein Ingestion Increases Muscle Protein Synthesis Rates at Rest and during Recovery from Exercise in Humans. Med. Sci. Sports Exerc. 2022, 54, 1572–1581. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.W.; Kozior, M.; Lynch, A.E.; Bass, J.J.; Atherton, P.J.; Smith, K.; Jakeman, P.M. The Effect of Fava Bean (Vicia faba L.) Protein Ingestion on Myofibrillar Protein Synthesis at Rest and after Resistance Exercise in Healthy, Young Men and Women: A Randomised Control Trial. Nutrients 2022, 14, 3688. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.S.; McCormick, M.C.; Robergs, R.A. Interaction among skeletal muscle metabolic energy systems during intense exercise. J. Nutr. Metab. 2010, 2010, 905612. [Google Scholar] [CrossRef] [Green Version]
- Stupka, N.; Gregorevic, P.; Plant, D.R.; Lynch, G.S. The calcineurin signal transduction pathway is essential for successful muscle regeneration in mdx dystrophic mice. Acta Neuropathol. 2004, 107, 299–310. [Google Scholar] [CrossRef]
- Lee, J.H.; Jun, H.S. Role of myokines in regulating skeletal muscle mass and function. Front. Physiol. 2019, 10, 42. [Google Scholar] [CrossRef]
- Liu, T.-Y.; Shi, C.-X.; Gao, R.; Sun, H.-J.; Xiong, X.-Q.; Ding, L.; Chen, Q.; Li, Y.-H.; Wang, J.-J.; Kang, Y.-M.; et al. Irisin inhibits hepatic gluconeogenesis and increases glycogen synthesis via the PI3K/Akt pathway in type 2 diabetic mice and hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef]
- O’Leary, M.F.; Wallace, G.R.; Bennett, A.J.; Tsintzas, K.; Jones, S.W. IL-15 promotes human myogenesis and mitigates the detrimental effects of TNFα on myotube development. Sci. Rep. 2017, 7, 12997. [Google Scholar] [CrossRef] [Green Version]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Sun, H.; Sherrier, M.; Li, H. Skeletal Muscle and Bone—Emerging Targets of Fibroblast Growth Factor-21. Front. Physiol. 2021, 12, 269. [Google Scholar] [CrossRef]
- Sharma, M.; Langley, B.; Bass, J.; Kambadur, R. Myostatin in muscle growth and repair. Exerc. Sport Sci. Rev. 2001, 29, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Ortega, D.R.; López, A.M.; Amaya, H.M.; de La Rosa, F.J.B. Tart cherry and pomegranate supplementations enhance recovery from exercise-induced muscle damage: A systematic review. Biol. Sport 2021, 38, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Funes, L.; Carrera-Quintanar, L.; Cerdán-Calero, M.; Ferrer, M.D.; Drobnic, F.; Pons, A.; Roche, E.; Micol, V. Effect of lemon verbena supplementation on muscular damage markers, proinflammatory cytokines release and neutrophils’ oxidative stress in chronic exercise. Eur. J. Appl. Physiol. 2011, 111, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Howard, E.E.; Pasiakos, S.M.; Blesso, C.N.; Fussell, M.A.; Rodriguez, N.R. Divergent Roles of Inflammation in Skeletal Muscle Recovery From Injury. Front. Physiol. 2020, 11, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbotina, E.; Sierra, A.; Zhu, Z.; Gao, Z.; Koganti, S.R.K.; Reyes, S.; Stepniak, E.; Walsh, S.A.; Acevedo, M.R.; Perez-Terzic, C.M.; et al. Musclin is an activity-stimulated myokine that enhances physical endurance. Proc. Natl. Acad. Sci. USA 2015, 112, 16042–16047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Gatta, P.A.; Cameron-Smith, D.; Peake, J.M. Acute resistance exercise increases the expression of chemotactic factors within skeletal muscle. Eur. J. Appl. Physiol. 2014, 114, 2157–2167. [Google Scholar] [CrossRef] [Green Version]
- Gelabert-Rebato, M.; Martin-Rincon, M.; Galvan-Alvarez, V.; Gallego-Selles, A.; Martinez-Canton, M.; Vega-Morales, T.; Wiebe, J.C.; Castillo, C.F.-D.; Castilla-Hernandez, E.; Diaz-Tiberio, O.; et al. A single dose of the mango leaf extract zynamite® in combination with quercetin enhances peak power output during repeated sprint exercise in men and women. Nutrients 2019, 11, 2592. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
|
|
| Participant ID | Group | Issue | Adjudicated Result |
|---|---|---|---|
| 7 | Placebo | Exercise performed within 48 h of test day. Borg Scale ≤ 13 | Include in PP |
| 9 | Placebo | Exercise performed within 48 h of test day. Borg Scale > 13 | Exclude from PP |
| 12 | Placebo | Exercise performed within 48 h of test day. Borg Scale > 13 | Exclude from PP |
| 15 | Placebo | Exercise performed within 48 h of test day. Borg Scale > 13 | Exclude from PP |
| 38 | Placebo | Exercise performed within 48 h of test day. Borg Scale ≤ 13 | Include in PP |
| 40 | Placebo | Exercise performed within 48 h of test day. Borg Scale > 13 | Exclude from PP |
| 8 | NPN_1 | Exercise performed within 48 h of test day. Borg Scale ≤ 13 | Include in PP |
| 10 | NPN_1 | Exercise performed within 48 h of test day. Borg Scale ≤ 13 | Include in PP |
| 11 | NPN_1 | Exercise performed within 48 h of test day. Borg Scale ≤ 13 | Include in PP |
| 22 | NPN_1 | Exercise performed within 48 h of test day. Borg Scale > 13 | Exclude from PP |
| Placebo (n = 10) | NPN_1 (n = 14) | |
|---|---|---|
| Age (years) | 38 ± 4.8 | 37.1 ± 5.0 |
| Height (cm) | 180.8 ± 5.8 | 182.9 ± 7.1 |
| Body Mass (kg) | 86.3 ± 11.0 | 85.8 ± 9.9 |
| BMI (kg/m2) | 26.3 ± 2.3 | 25.6 ± 2.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerr, A.; Hart, L.; Davis, H.; Wall, A.; Lacey, S.; Franklyn-Miller, A.; Khaldi, N.; Keogh, B. Improved Strength Recovery and Reduced Fatigue with Suppressed Plasma Myostatin Following Supplementation of a Vicia faba Hydrolysate, in a Healthy Male Population. Nutrients 2023, 15, 986. https://doi.org/10.3390/nu15040986
Kerr A, Hart L, Davis H, Wall A, Lacey S, Franklyn-Miller A, Khaldi N, Keogh B. Improved Strength Recovery and Reduced Fatigue with Suppressed Plasma Myostatin Following Supplementation of a Vicia faba Hydrolysate, in a Healthy Male Population. Nutrients. 2023; 15(4):986. https://doi.org/10.3390/nu15040986
Chicago/Turabian StyleKerr, Alish, Luke Hart, Heidi Davis, Audrey Wall, Seán Lacey, Andrew Franklyn-Miller, Nora Khaldi, and Brian Keogh. 2023. "Improved Strength Recovery and Reduced Fatigue with Suppressed Plasma Myostatin Following Supplementation of a Vicia faba Hydrolysate, in a Healthy Male Population" Nutrients 15, no. 4: 986. https://doi.org/10.3390/nu15040986