
Osteocytic Sclerostin Expression as an Indicator of Altered Bone Turnover
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parathyroidectomized (PTX) Rat Model
2.2. Patient Cohort
2.3. Serum Biochemistry
2.4. Bone Histomorphometric Analysis
2.5. Osteocyte Quantification
2.6. Quantification of Sclerostin-Positive Osteocytes
2.7. Statistical Analysis
3. Results
3.1. PTH Exerts a Direct Inhibitory Effect on Bone Sclerostin Expression in PTX Rats
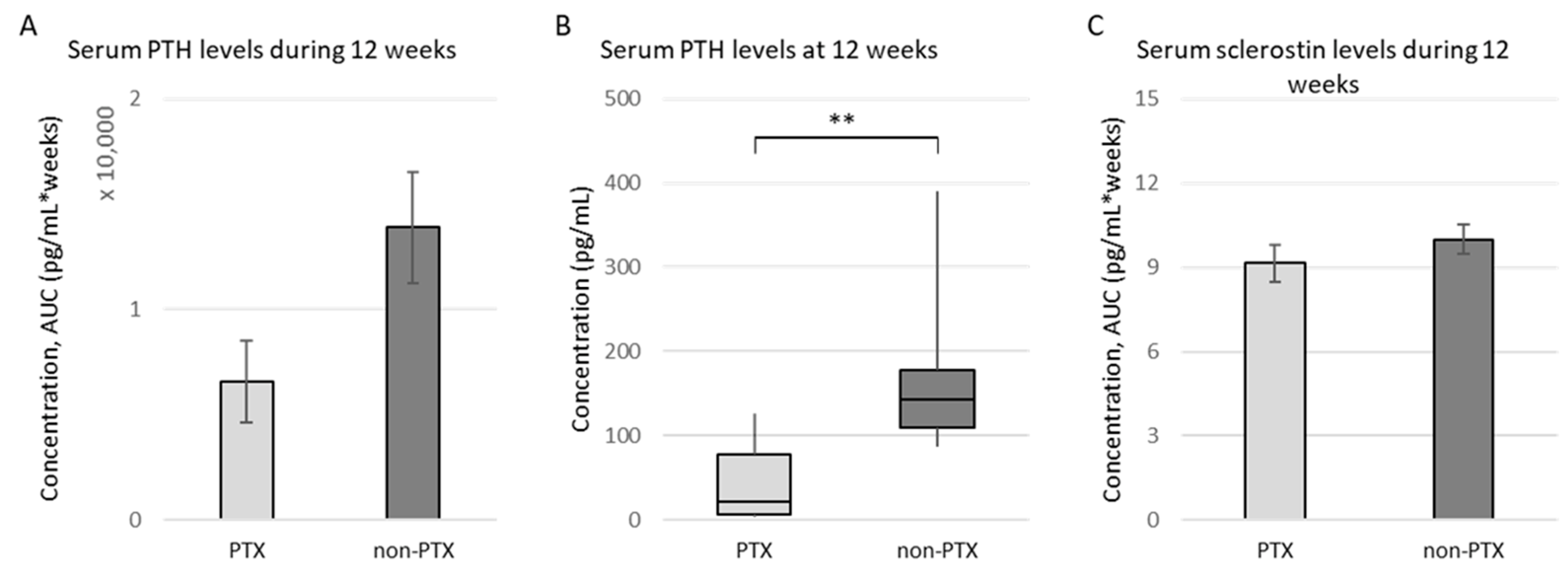
3.1.1. Characteristics of (Non-)PTX Rats
3.1.2. Quantification of (Sclerostin-Positive) Osteocytes in Bone of (Non-)PTX Rats
3.1.3. Serum iPTH Is Inversely Correlated with Sclerostin Expression in Trabecular Bone
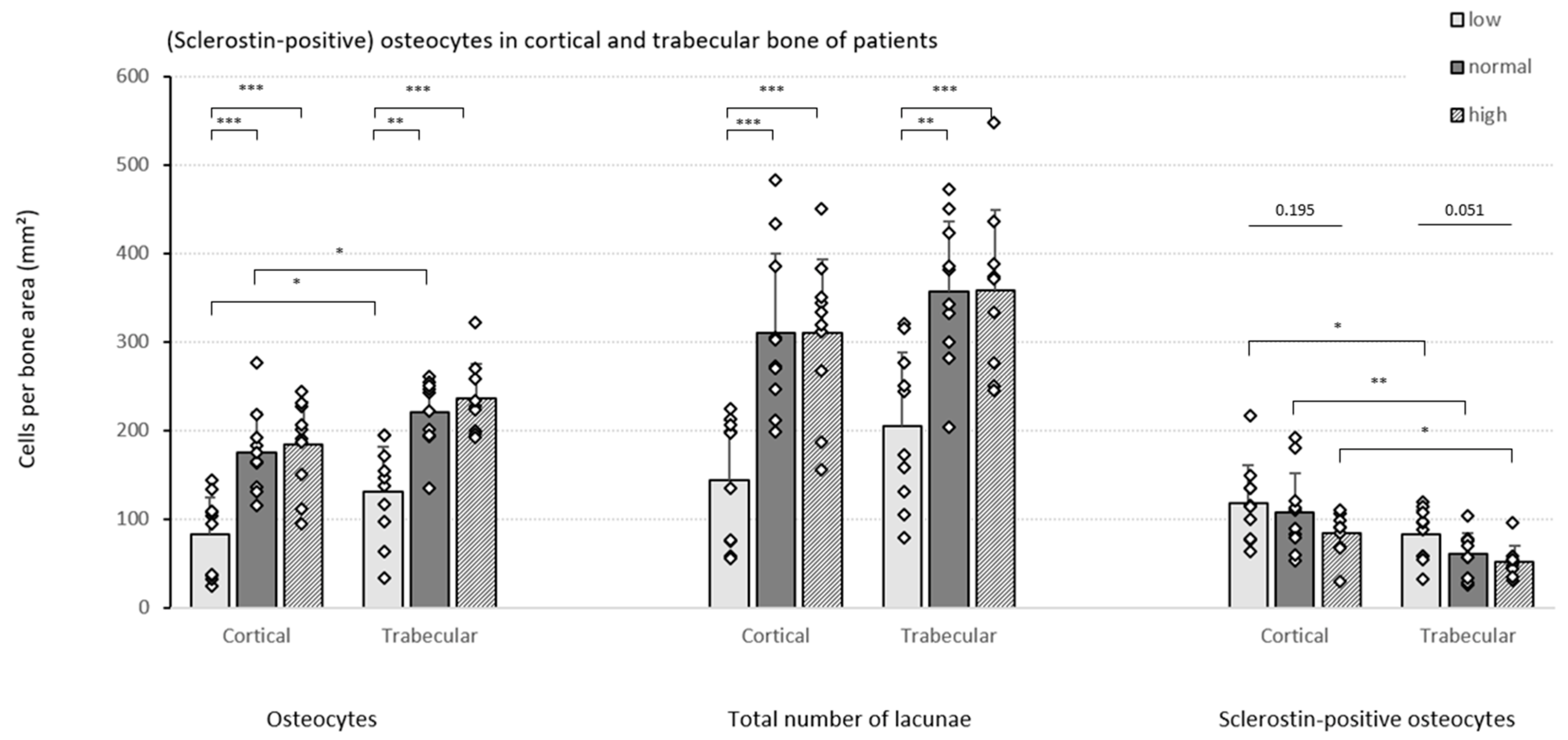
3.2. Sclerostin Expression, Number of Osteocytes and Bone Histomorphometry in Patients with Different Degrees of Bone Turnover
3.2.1. Demographics and Specific Characteristics of the Cohort
3.2.2. Quantification of (Sclerostin-Positive) Osteocytes in Patients with Varying Degrees of Bone Turnover
3.2.3. Specific Bone Turnover Parameters Reflect Osteocytic Sclerostin Expression
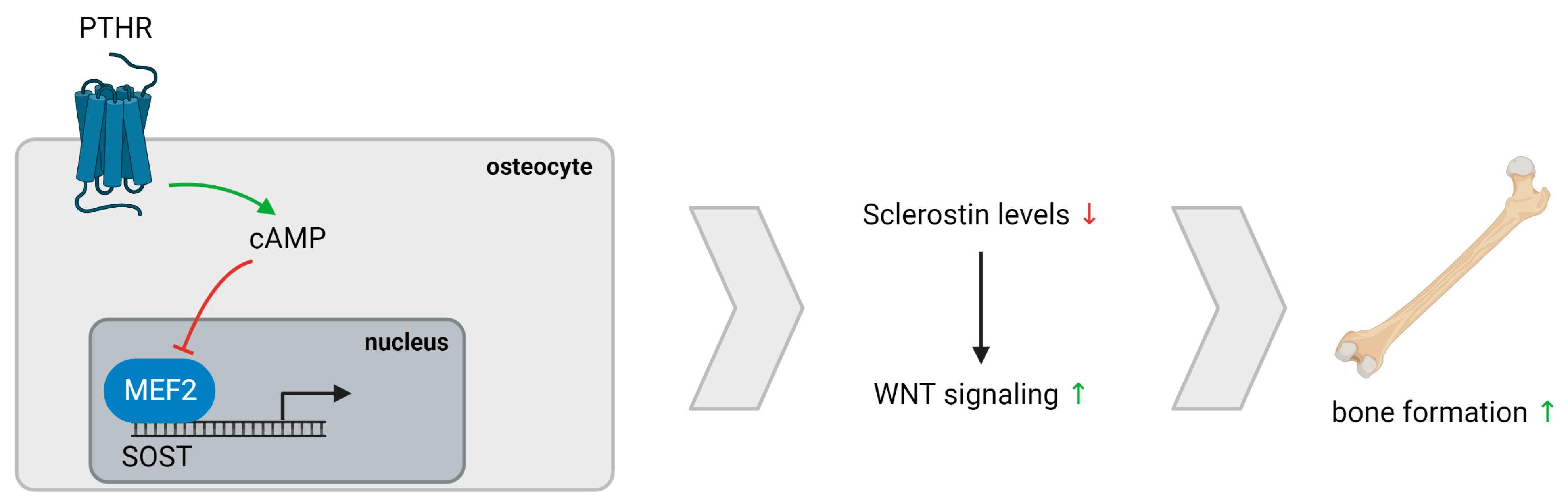
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular mechanosensors in osteocytes. Bone Res. 2020, 8, 23. [Google Scholar] [CrossRef]
- Bonewald, L.F.; Johnson, M.L. Osteocytes, mechanosensing and Wnt signaling. Bone 2008, 42, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Bonewald, L.F. The Role of the Osteocyte in Bone and Nonbone Disease. Endocrinol. Metab. Clin. N. Am. 2017, 46, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huybrechts, Y.; Mortier, G.; Boudin, E.; Van Hul, W. WNT Signaling and Bone: Lessons From Skeletal Dysplasias and Disorders. Front. Endocrinol. 2020, 11, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin binds to LRP5/6 and antagonizes canonical Wnt signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, F.; Mawad, H.; El-Husseini, A.A.; Davenport, D.L.; Malluche, H.H. Serum bone markers in ROD patients across the spectrum of decreases in GFR: Activin A increases before all other markers . Clin. Nephrol. 2019, 91, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Cejka, D.; Marculescu, R.; Kozakowski, N.; Plischke, M.; Reiter, T.; Gessl, A.; Haas, M. Renal elimination of sclerostin increases with declining kidney function. J. Clin. Endocrinol. Metab. 2014, 99, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Hruska, K.A.; Sugatani, T.; Agapova, O.; Fang, Y. The chronic kidney disease—Mineral bone disorder (CKD-MBD): Advances in pathophysiology. Bone 2017, 100, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Ott, S.M. Renal Osteodystrophy-Time for Common Nomenclature. Curr. Osteoporos. Rep. 2017, 15, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Moe, S.; Drüeke, T.; Cunningham, J.; Goodman, W.; Martin, K.; Olgaard, K.; Ott, S.; Sprague, S.; Lameire, N.; Eknoyan, G. Definition, evaluation, and classification of renal osteodystrophy: A position statement from Kidney Disease: Improving Global Outcomes (KDIGO). Kidney Int. 2006, 69, 1945–1953. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.C.; Cohen-Solal, M.; D’Haese, P.C.; Ferreira, A. The Role of Bone Biopsy in the Management of CKD-MBD. Calcif. Tissue Int. 2021, 108, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P.; D’Haese, P.; Bacchetta, J.; Cannata-Andia, J.; Ferreira, A.; Haarhaus, M.; Mazzaferro, S.; Lafage Proust, M.H.; Salam, S.; Spasovski, G.; et al. Bone biopsy practice patterns across Europe: The European renal osteodystrophy initiative-a position paper. Nephrol. Dial. Transplant. 2017, 32, 1608–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalle Carbonare, L.; Valenti, M.T.; Giannini, S.; Gallieni, M.; Stefani, F.; Ciresa, R.; Politi, C.; Fusaro, M.J.J.o.C.M. Bone Biopsy for Histomorphometry in Chronic Kidney Disease (CKD): State-of-the-Art and New Perspectives. J. Clin. Med. 2021, 10, 4617. [Google Scholar] [CrossRef] [PubMed]
- Sprague, S.M.; Bellorin-Font, E.; Jorgetti, V.; Carvalho, A.B.; Malluche, H.H.; Ferreira, A.; D’Haese, P.C.; Drüeke, T.B.; Du, H.; Manley, T.; et al. Diagnostic Accuracy of Bone Turnover Markers and Bone Histology in Patients With CKD Treated by Dialysis. Am. J. Kidney Dis. 2016, 67, 559–566. [Google Scholar] [CrossRef]
- Zeng, S.; Querfeld, U.; Feger, M.; Haffner, D.; Hasan, A.A.; Chu, C.; Slowinski, T.; Bernd Dschietzig, T.; Schäfer, F.; Xiong, Y.; et al. Relationship between GFR, intact PTH, oxidized PTH, non-oxidized PTH as well as FGF23 in patients with CKD. FASEB J. 2020, 34, 15269–15281. [Google Scholar] [CrossRef] [PubMed]
- Bellorin-Font, E.; Voinescu, A.; Martin, K.J. Chapter 23—Calcium, phosphate, PTH, vitamin D, and FGF-23 in CKD-mineral and bone disorder. In Nutritional Management of Renal Disease, 4th ed.; Kopple, J.D., Massry, S.G., Kalantar-Zadeh, K., Fouque, D., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 353–381. [Google Scholar] [CrossRef]
- Torres, P.A.; De Brauwere, D.P. Three feedback loops precisely regulating serum phosphate concentration. Kidney Int. 2011, 80, 443–445. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Jose, A.; Sharma, S. Physiology, Parathyroid Hormone. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Chiang, C. The use of bone turnover markers in chronic kidney disease-mineral and bone disorders. Nephrology 2017, 22 (Suppl. S2), 11–13. [Google Scholar] [CrossRef] [Green Version]
- Vervloet, M.G.; Brandenburg, V.M. Circulating markers of bone turnover. J. Nephrol. 2017, 30, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, H.S.; Behets, G.; Viaene, L.; Bammens, B.; Claes, K.; Meijers, B.; Naesens, M.; Sprangers, B.; Kuypers, D.; Cavalier, E.; et al. Diagnostic Accuracy of Noninvasive Bone Turnover Markers in Renal Osteodystrophy. Am. J. Kidney Dis. 2022, 79, 667–676.e1. [Google Scholar] [CrossRef] [PubMed]
- Smout, D.; Jørgensen, H.S.; Cavalier, E.; Evenepoel, P. Clinical utility of bone turnover markers in patients with chronic kidney disease. Curr. Opin. Nephrol. Hypertens. 2022, 31, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Cavalier, E.; Souberbielle, J.C.; Delanaye, P. PTH determination in hemodialyzed patients—A laboratory perspective. Semin. Dial. 2019, 32, 490–492. [Google Scholar] [CrossRef]
- Bover, J.; Ureña-Torres, P.; Cozzolino, M.; Rodríguez-García, M.; Gómez-Alonso, C. The Non-invasive Diagnosis of Bone Disorders in CKD. Calcif. Tissue Int. 2021, 108, 512–527. [Google Scholar] [CrossRef]
- Behets, G.J.; Viaene, L.; Meijers, B.; Blocki, F.; Brandenburg, V.M.; Verhulst, A.; D’Haese, P.C.; Evenepoel, P. Circulating levels of sclerostin but not DKK1 associate with laboratory parameters of CKD-MBD. PLoS ONE 2017, 12, e0176411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.; Magalhães, J.; Mendonça, L.; Neto, R.; Santos, J.; Carvalho, C.G.; Oliveira, A.; Beco, A.; Frazão, J. Evaluation of Renal Osteodystrophy and Serum Bone-Related Biomarkers in a Peritoneal Dialysis Population. J. Bone Miner. Res. 2022, 37, 1689–1699. [Google Scholar] [CrossRef]
- Salam, S.; Gallagher, O.; Gossiel, F.; Paggiosi, M.; Khwaja, A.; Eastell, R. Diagnostic accuracy of biomarkers and imaging for bone turnover in renal osteodystrophy. J. Am. Soc. Nephrol. 2018, 29, 1557–1565. [Google Scholar] [CrossRef] [Green Version]
- Opdebeeck, B.; Maudsley, S.; Azmi, A.; De Maré, A.; De Leger, W.; Meijers, B.; Verhulst, A.; Evenepoel, P.; D’Haese, P.C.; Neven, E. Indoxyl Sulfate and p-Cresyl Sulfate Promote Vascular Calcification and Associate with Glucose Intolerance. J. Am. Soc. Nephrol. JASN 2019, 30, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Murton, M.; Goff-Leggett, D.; Bobrowska, A.; Garcia Sanchez, J.J.; James, G.; Wittbrodt, E.; Nolan, S.; Sörstadius, E.; Pecoits-Filho, R.; Tuttle, K.J.A.i.t. Burden of chronic kidney disease by KDIGO categories of glomerular filtration rate and albuminuria: A systematic review. Adv. Ther. 2021, 38, 180–200. [Google Scholar] [CrossRef]
- Behets, G.J.; Spasovski, G.; Sterling, L.R.; Goodman, W.G.; Spiegel, D.M.; De Broe, M.E.; D’Haese, P.C. Bone histomorphometry before and after long-term treatment with cinacalcet in dialysis patients with secondary hyperparathyroidism. Kidney Int. 2015, 87, 846–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouillon, R.; Coopmans, W.; Degroote, D.E.; Radoux, D.; Eliard, P.H. Immunoradiometric assay of parathyrin with polyclonal and monoclonal region-specific antibodies. Clin. Chem. 1990, 36, 271–276. [Google Scholar] [CrossRef]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Atkins, G.J.; Rowe, P.S.; Lim, H.P.; Welldon, K.J.; Ormsby, R.; Wijenayaka, A.R.; Zelenchuk, L.; Evdokiou, A.; Findlay, D.M. Sclerostin is a locally acting regulator of late-osteoblast/preosteocyte differentiation and regulates mineralization through a MEPE-ASARM-dependent mechanism. J. Bone Miner. Res. 2011, 26, 1425–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellido, T.; Saini, V.; Pajevic, P.D. Effects of PTH on osteocyte function. Bone 2013, 54, 250–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Calle, J.; Sato, A.Y.; Bellido, T. Role and mechanism of action of sclerostin in bone. Bone 2017, 96, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Leupin, O.; Kramer, I.; Collette, N.M.; Loots, G.G.; Natt, F.; Kneissel, M.; Keller, H. Control of the SOST bone enhancer by PTH using MEF2 transcription factors. J. Bone Miner. Res. 2007, 22, 1957–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenepoel, P.; Claes, K.; Viaene, L.; Bammens, B.; Meijers, B.; Naesens, M.; Sprangers, B.; Kuypers, D. Decreased Circulating Sclerostin Levels in Renal Transplant Recipients With Persistent Hyperparathyroidism. Transplantation 2016, 100, 2188–2193. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T.; Ali, A.A.; Gubrij, I.; Plotkin, L.I.; Fu, Q.; O’Brien, C.A.; Manolagas, S.C.; Jilka, R.L. Chronic elevation of parathyroid hormone in mice reduces expression of sclerostin by osteocytes: A novel mechanism for hormonal control of osteoblastogenesis. Endocrinology 2005, 146, 4577–4583. [Google Scholar] [CrossRef]
- Pires, G.O.; Vieira, I.O.; Hernandes, F.R.; Teixeira, A.L.; Oliveira, I.B.; Dominguez, W.V.; dos Reis, L.M.; Montenegro, F.M.; Moysés, R.M.; Carvalho, A.B.; et al. Effects of parathyroidectomy on the biology of bone tissue in patients with chronic kidney disease and secondary hyperparathyroidism. Bone 2019, 121, 277–283. [Google Scholar] [CrossRef]
- Neto, R.; Pereira, L.; Magalhães, J.; Quelhas-Santos, J.; Martins, S.; Carvalho, C.; Frazão, J.M. Sclerostin and DKK1 circulating levels associate with low bone turnover in patients with chronic kidney disease Stages 3 and 4. Clin. Kidney J. 2021, 14, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- van Lierop, A.H.; Hamdy, N.A.; Hamersma, H.; van Bezooijen, R.L.; Power, J.; Loveridge, N.; Papapoulos, S.E. Patients with sclerosteosis and disease carriers: Human models of the effect of sclerostin on bone turnover. J. Bone Miner. Res. 2011, 26, 2804–2811. [Google Scholar] [CrossRef]
- Li, J.; Bao, Q.; Chen, S.; Liu, H.; Feng, J.; Qin, H.; Li, A.; Liu, D.; Shen, Y.; Zhao, Y.; et al. Different bone remodeling levels of trabecular and cortical bone in response to changes in Wnt/β-catenin signaling in mice. J. Orthop. Res. 2017, 35, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Ott, S.M. Cortical or Trabecular Bone: What’s the Difference? Am. J. Nephrol. 2018, 47, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Boltenstål, H.; Qureshi, A.R.; Behets, G.J.; Lindholm, B.; Stenvinkel, P.; D’Haese, P.C.; Haarhaus, M. Association of Serum Sclerostin with Bone Sclerostin in Chronic Kidney Disease is Lost in Glucocorticoid Treated Patients. Calcif. Tissue Int. 2019, 104, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Forster, C.M.; White, C.A.; Turner, M.E.; Norman, P.A.; Ward, E.C.; Hopman, W.M.; Adams, M.A.; Holden, R.M. Circulating Levels of Dickkopf-Related Protein 1 Decrease as Measured GFR Declines and Are Associated with PTH Levels. Am. J. Nephrol. 2020, 51, 871–880. [Google Scholar] [CrossRef]
- Kuroda, T.; Shiraki, M.; Nakamura, T.; Suzuki, H.; Hiraishi, K.; Sugimoto, T.; Tanaka, S. Relationship Between Changes in Serum Levels of Intact Parathyroid Hormone and Sclerostin After a Single Dose of Zoledronic Acid: Results of a Phase 1 Pharmacokinetic Study. Calcif. Tissue Int. 2022, 110, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Pura, L.; Bandiara, R.; Gondodiputro, R.S. Hypertension. Sclerostin serum concentration in patients with predialysis CKD stage 3-5. Indones. J. Kidney Hypertens. 2018, 1, 1. [Google Scholar] [CrossRef]
- Sabbagh, Y.; Graciolli, F.G.; O’Brien, S.; Tang, W.; dos Reis, L.M.; Ryan, S.; Phillips, L.; Boulanger, J.; Song, W.; Bracken, C.; et al. Repression of osteocyte Wnt/β-catenin signaling is an early event in the progression of renal osteodystrophy. J. Bone Miner. Res. 2012, 27, 1757–1772. [Google Scholar] [CrossRef] [PubMed]
- Graciolli, F.G.; Neves, K.R.; Barreto, F.; Barreto, D.V.; Dos Reis, L.M.; Canziani, M.E.; Sabbagh, Y.; Carvalho, A.B.; Jorgetti, V.; Elias, R.M.J.K.i. The complexity of chronic kidney disease–mineral and bone disorder across stages of chronic kidney disease. Kidney Int. 2017, 91, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, M.B.; Tsai, J.N.; Wein, M.N. Bone Turnover Markers in the Diagnosis and Monitoring of Metabolic Bone Disease. Clin. Chem. 2017, 63, 464–474. [Google Scholar] [CrossRef] [Green Version]
- Samelson, E.J.; Broe, K.E.; Xu, H.; Yang, L.; Boyd, S.; Biver, E.; Szulc, P.; Adachi, J.; Amin, S.; Atkinson, E.; et al. Cortical and trabecular bone microarchitecture as an independent predictor of incident fracture risk in older women and men in the Bone Microarchitecture International Consortium (BoMIC): A prospective study. Lancet. Diabetes Endocrinol. 2019, 7, 34–43. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter (Unit) | Osteocytes per Bone Area | Sclerostin-Positive Osteocytes per Bone Area | ||
|---|---|---|---|---|
| PTX, n = 10 | Non-PTX, n = 9 | PTX, n = 10 | Non-PTX, n = 9 | |
| BFR (µm2/mm2/day) | −0.050 (0.891) | 0.048 (0.911) | −0.506 (0.135) | −0.238 (0.570) |
| B.Ar (%) | −0.248 (0.489) | −0.786 (0.021) | 0.333 (0.347) | 0.429 (0.289) |
| O.Ar (%) | −0.134 (0.713) | −0.143 (0.736) | −0.456 (0.185) | 0.262 (0.531) |
| O.Pm (%) | −0.146 (0.688) | −0.262 (0.531) | −0.468 (0.172) | 0.333 (0.420) |
| O.Wi (µm) | 0.061 (0.868) | −0.071 (0.867) | −0.334 (0.345) | 0.024 (0.955) |
| Ob.Pm.O (%) | 0.055 (0.880) | −0.838 (0.009) | −0.178 (0.623) | 0.659 (0.076) |
| Ob.Pm.T (%) | 0.006 (0.987) | −0.647 (0.083) | −0.276 (0.440) | 0.443 (0.272) |
| E.Pm (%) | 0.418 (0.229) | 0.048 (0.911) | 0.382 (0.276) | −0.643 (0.086) |
| Oc.Pm.E (%) | 0.042 (0.907) | −0.143 (0.736) | 0.309 (0.385) | −0.095 (0.823) |
| Oc.Pm.T (%) | 0.370 (0.293) | −0.119 (0.779) | 0.430 (0.214) | −0.548 (0.160) |
| MAR (µm/day) | −0.263 (0.464) | −0.405 (0.320) | −0.506 (0.135) | 0.071 (0.867) |
| Aj.AR (µm/day) | −0.163 (0.654) | 0.238 (0.570) | −0.719 (0.019) | −0.619 (0.102) |
| Mlt (days) | 0.188 (0.604) | −0.048 (0.911) | 0.757 (0.011) | 0.381 (0.352) |
| Omt (days) | 0.388 (0.569) | 0.238 (0.570) | 0.569 (0.086) | 0.167 (0.693) |
| Compartment | PTX, n = 10 | Non-PTX, n = 9 |
|---|---|---|
| Cortical | −0.095 (0.823) | 0.071 (0.867) |
| Trabecular | −0.714 (0.047) | −0.429 (0.289) |
| Parameter (Unit) | All Patients | Low Turnover | Normal Turnover | High Turnover | p-Value |
|---|---|---|---|---|---|
| Demographics | |||||
| Patients (male) | 30 (19) | 10 (8) | 10 (8) | 10 (3) | 0.041 |
| Age (years) | 52.1 (15.7) | 58.0 (12.6) | 51.2 (6.6) | 47.9 (15.0) | 0.236 |
| Status (Tx/CKD5) | 20/10 | 10/0 | 7/3 | 3/7 | 0.0039 |
| Histomorphometric parameters | |||||
| BFR (µm2/mm2/day) | 379.92 (551.3) | 54.07 (41.80) | 379.92 (151.66) | 827.61 (776.26) | 0.000 abc |
| B.Ar (%) | 19.61 (7.78) | 15.50 (3.79) | 21.16 (3.68) | 23.51 (8.25) | 0.039 a |
| O.Ar (%) | 4.76 (4.68) | 1.44 (3.56) | 4.56 (3.16) | 7.31 (5.51) | 0.034 c |
| O.Pm (%) | 37.04 (27.23) | 16.26 (21.16) | 41.55 (49.06) | 46.32 (18.11) | 0.010 ac |
| O.Wi (µm) | 8.29 (4.25) | 6.89 (1.47) | 8.25 (3.89) | 10.38 (2.29) | 0.016 c |
| Ob.Pm.O (%) | 25.65 (36.68) | 23.41 (29.34) | 34.94 (29.00) | 28.38 (27.92) | 0.440 |
| Ob.Pm.T (%) | 7.01 (16.81) | 2.45 (4.52) | 9.74 (17.88) | 12.32 (17.51) | 0.029 c |
| E.Pm (%) | 3.14 (3.17) | 2.15 (2.21) | 3.95 (3.87) | 3.96 (6.71) | 0.076 |
| Oc.Pm.E (%) | 31.91 (22.98) | 26.93 (32.86) | 32.95 (11.44) | 25.78 (21.53) | 0.715 |
| Oc.Pm.T (%) | 0.92 (1.03) | 0.48 (0.89) | 1.13 (1.08) | 0.84 (2.68) | 0.120 |
| MAR (µm/day) | 0.89 (0.45) | 0.64 (0.08) | 0.87 (0.15) | 1.20 (0.30) | 0.000 abc |
| Aj.AR (µm/day) | 0.35 (0.36) | 0.06 (0.18) | 0.39 (0.22) | 0.63 (0.17) | 0.000 abc |
| Mlt (days) | 26.39 (31.47) | 94.36 (143.87) | 27.68 (13.12) | 14.73 (8.97) | 0.001 ac |
| Omt (days) | 9.71 (3.58) | 10.70 (1.87) | 9.89 (4.42) | 8.90 (2.80) | 0.246 |
| Serum factors | |||||
| Creatinine (mg/dL) | 1.71 (4.52) | 1.47 (0.37) | 1.81 (1.05) | 6.44 (4.84) | 0.053 |
| eGFR | 41 (42) | 49 (17) | 42 (22) | 7 (17) | 0.012 c |
| (mL/min/1.73 m2) | |||||
| Calcium (mmol/L) | 2.43 (0.20) | 2.43 (0.22) | 2.42 (0.22) | 2.43 (0.26) | 0.820 |
| Phosphate (mmol/L) | 1.05 (0.55) | 0.97 (0.28) | 0.97 (0.31) | 1.48 (0.68) | 0.197 |
| 25-OH Vit D (µg/L) | 33.50 (14.63) | 35.05 (9.58) | 33.20 (8.70) | 31.00 (13.30) | 0.595 |
| iPTH (ng/dL) | 109.20 (365.50) | 66.35 (45.33) | 111.60 (54.10) | 427.90 (576.90) | 0.005 c |
| ALP (U/L) | 88.12 (73.45) | 74.5 (30.78) | 92.00 (37.00) | 174.00 (97.06) | 0.001 bc |
| BALP (µg/L) | 23.36 (16.53) | 15.61 (11.04) | 27.68 (12.50) | 47.52 (12.33) | 0.007 c |
| PINP (ng/mL) | 80.06 (83.89) | 46.68 (38.15) | 88.33 (70.13) | 142.28 (17.31) | 0.002 c |
| TRAP5b (U/L) | 4.30 (3.38) | 3.60 (1.56) | 3.99 (3.12) | 7.86 (1.83) | 0.005 c |
| Sclerostin (ng/mL) | 0.82 (0.36) | 0.82 (0.26) | 0.84 (0.41) | 0.86 (0.52) | 0.859 |
| Parameter (Unit) | Osteocytes per Bone Area (p-Value) | Sclerostin Expression per Bone Area (p-Value) |
|---|---|---|
| BFR (µm2/mm2/day) | 0.694 (0.000) | −0.339 (0.072) |
| B.Ar (%) | 0.237 (0.215) | −0.110 (0.569) |
| O.Ar (%) | 0.509 (0.005) | 0.009 (0.962) |
| O.Pm (%) | 0.542 (0.002) | −0.003 (0.988) |
| O.Wi (µm) | 0.397 (0.033) | 0.000 (0.998) |
| Ob.Pm.O (%) | −0.046 (0.814) | 0.100 (0.605) |
| Ob.Pm.T (%) | 0.223 (0.263) | −0.005 (0.980) |
| E.Pm (%) | 0.319 (0.091) | −0.251 (0.189) |
| Oc.Pm.E (%) | 0.218 (0.256) | −0.246 (0.199) |
| Oc.Pm.T (%) | 0.259 (0.192) | −0.325 (0.098) |
| MAR (µm/day) | 0.494 (0.006) | −0.326 (0.085) |
| Aj.AR (µm/day) | 0.590 (0.001) | −0.481 (0.008) |
| Mlt (days) | −0.469 (0.010) | 0.526 (0.003) |
| Omt (days) | −0.149 (0.440) | 0.287 (0.132) |
| Parameter (Unit) | Sclerostin Expression Cortical Bone (p-Value) | Sclerostin Expression Trabecular Bone (p-Value) |
|---|---|---|
| Age (years) | 0.242 (0.198) | 0.084 (0.664) |
| Creatinine (mg/dL) | −0.422 (0.025) | −0.346 (0.077) |
| eGFR (mL/min/1.73 m2) | 0.414 (0.028) | 0.383 (0.049) |
| Calcium (mmol/L) | 0.056 (0.775) | 0.054 (0.791) |
| Phosphate (mmol/L) | −0.272 (0.162) | −0.195 (0.329) |
| 25-OH Vit D (µg/L) | −0.235 (0.230) | −0.201 (0.316) |
| iPTH (ng/dL) | −0.561 (0.002) | −0.363 (0.057) |
| ALP (U/L) | −0.387 (0.042) | −0.340 (0.083) |
| BALP (µg/L) | −0.528 (0.012) | −0.293 (0.186) |
| PINP (ng/mL) | −0.541 (0.006) | −0.474 (0.019) |
| TRAP5b (U/L) | −0.562 (0.003) | −0.234 (0.261) |
| Sclerostin (ng/dL) | −0.250 (0.219) | −0.136 (0.508) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huybrechts, Y.; Evenepoel, P.; Haarhaus, M.; Cavalier, E.; Dams, G.; Van Hul, W.; D’Haese, P.C.; Verhulst, A. Osteocytic Sclerostin Expression as an Indicator of Altered Bone Turnover. Nutrients 2023, 15, 598. https://doi.org/10.3390/nu15030598
Huybrechts Y, Evenepoel P, Haarhaus M, Cavalier E, Dams G, Van Hul W, D’Haese PC, Verhulst A. Osteocytic Sclerostin Expression as an Indicator of Altered Bone Turnover. Nutrients. 2023; 15(3):598. https://doi.org/10.3390/nu15030598
Chicago/Turabian StyleHuybrechts, Yentl, Pieter Evenepoel, Mathias Haarhaus, Etienne Cavalier, Geert Dams, Wim Van Hul, Patrick C. D’Haese, and Anja Verhulst. 2023. "Osteocytic Sclerostin Expression as an Indicator of Altered Bone Turnover" Nutrients 15, no. 3: 598. https://doi.org/10.3390/nu15030598