
The Effects of Cellular Membrane Damage on the Long-Term Storage and Adhesion of Probiotic Bacteria in Caco-2 Cell Line
 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum Preparation
2.2. Bacterial Cultures
2.3. Fluid Bed Drying
2.4. Plate Count Method
2.5. Intestinal Epithelial Cell Culture
2.6. Adhesion Assay
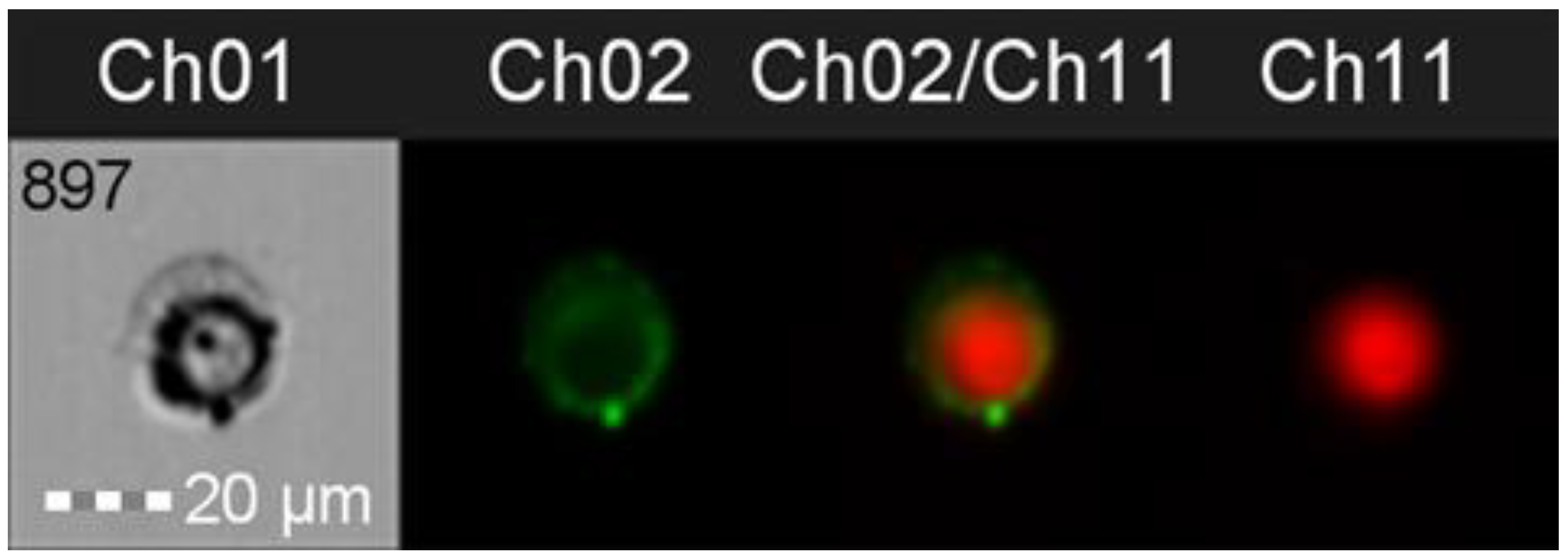
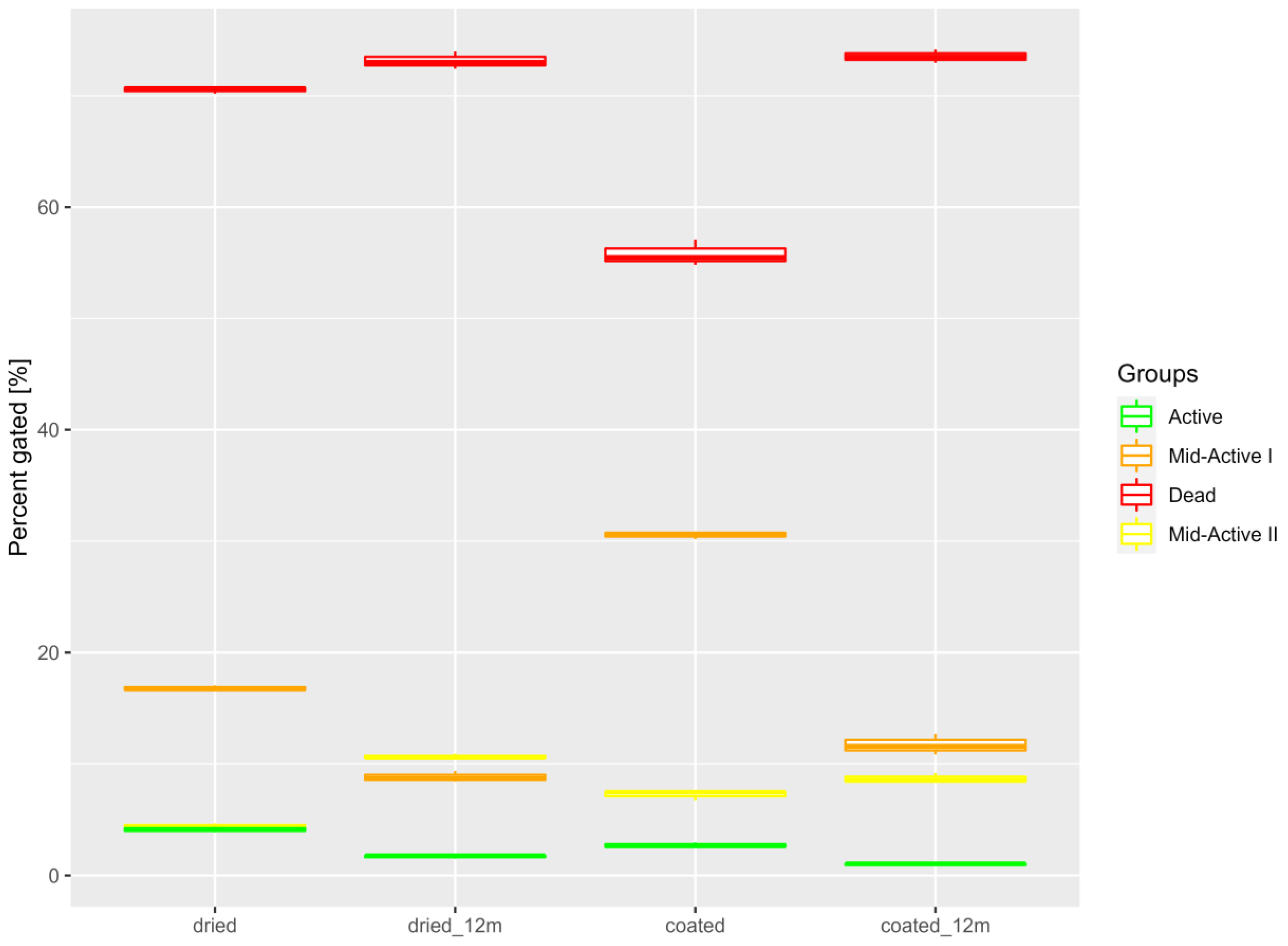
2.7. Imaging Flow Cytometry
2.8. Scanning Electron Microscopy (SEM)
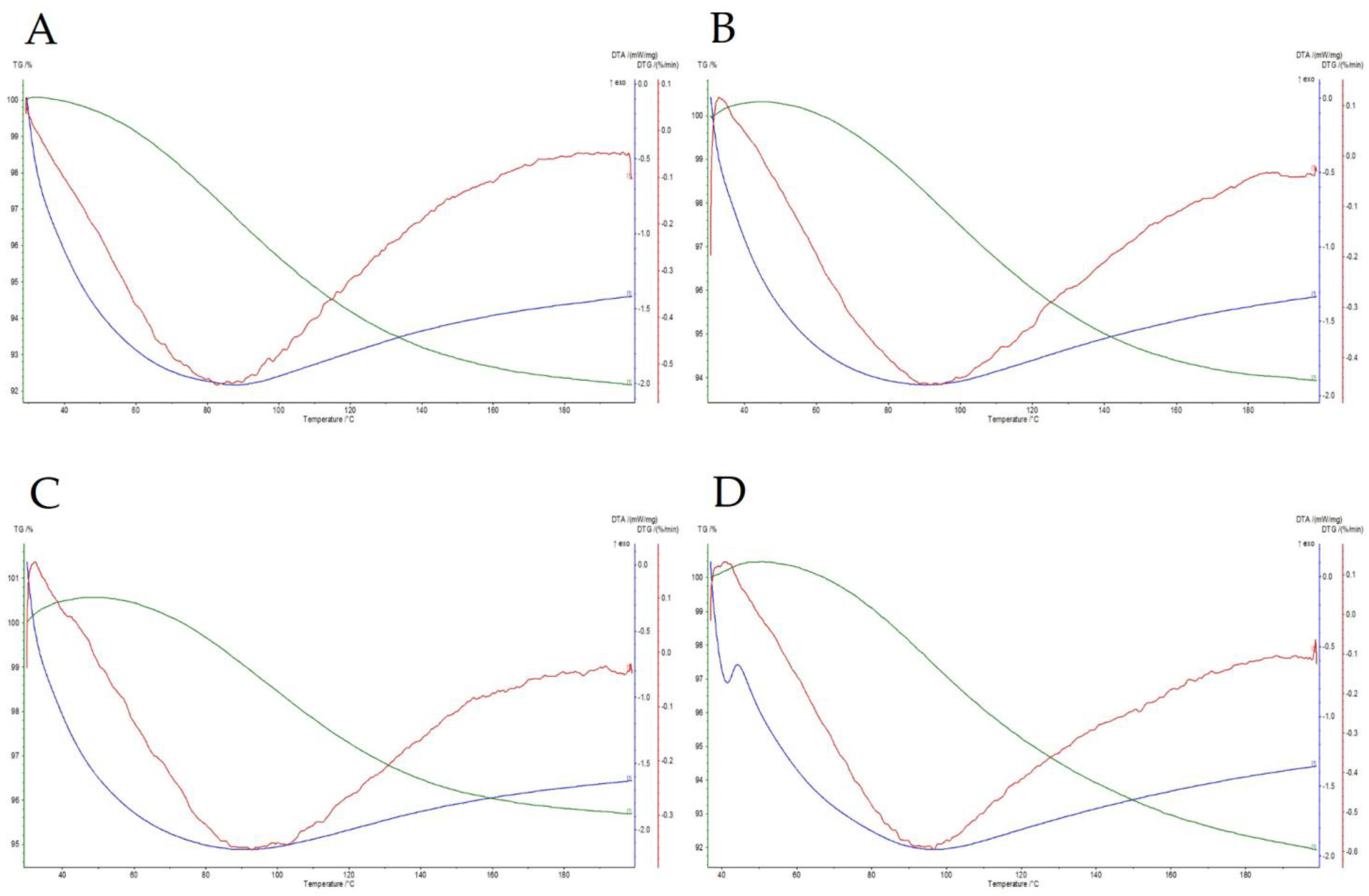
2.9. Thermogravimetry–Differential Thermal Analysis (TG/DTA)
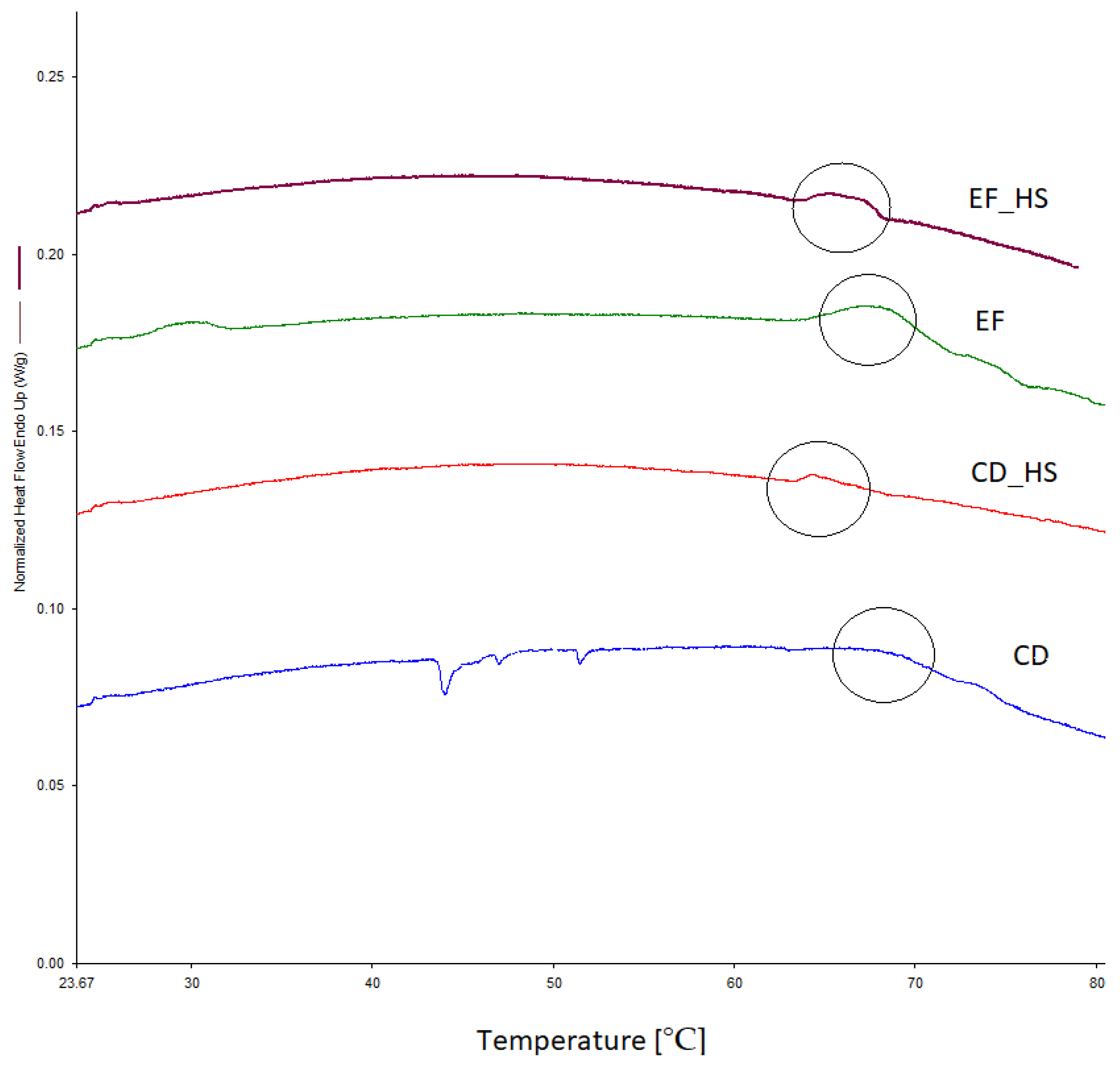
2.10. Differential Scanning Calorimetry (DSC)
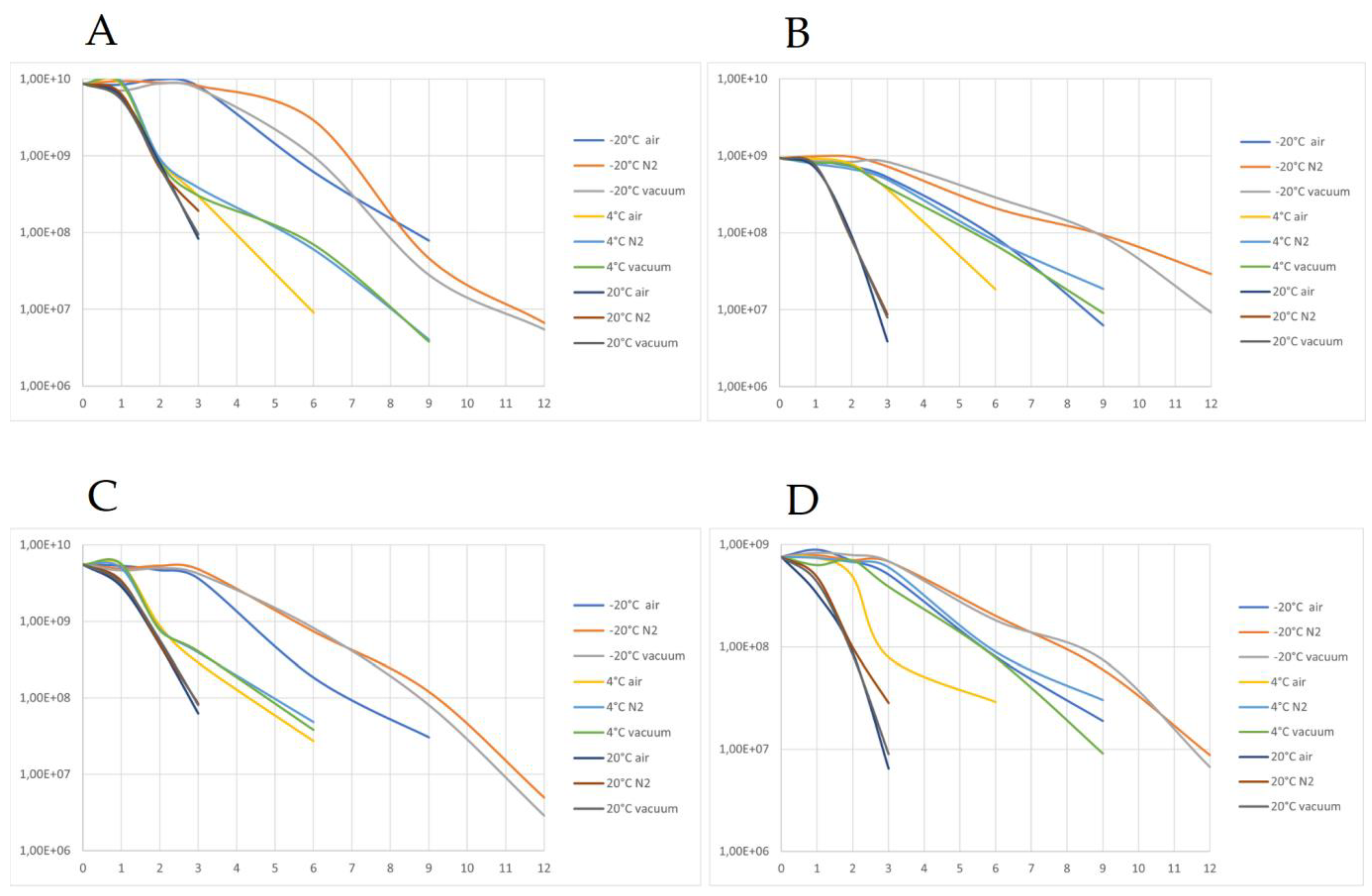
2.11. Storage and Shelf-Life Tests
3. Results and Discussion
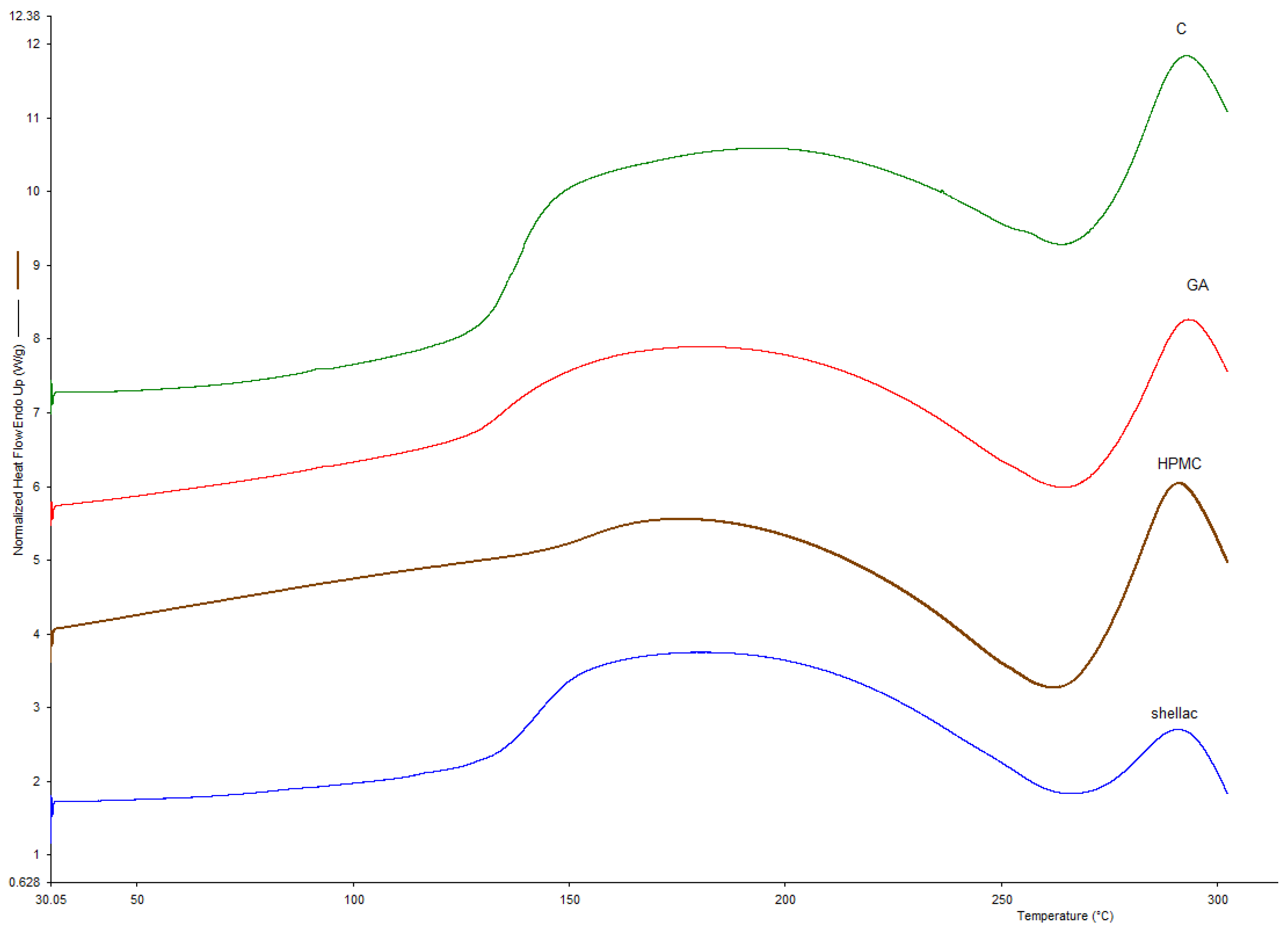
3.1. Glass Transition Temperature
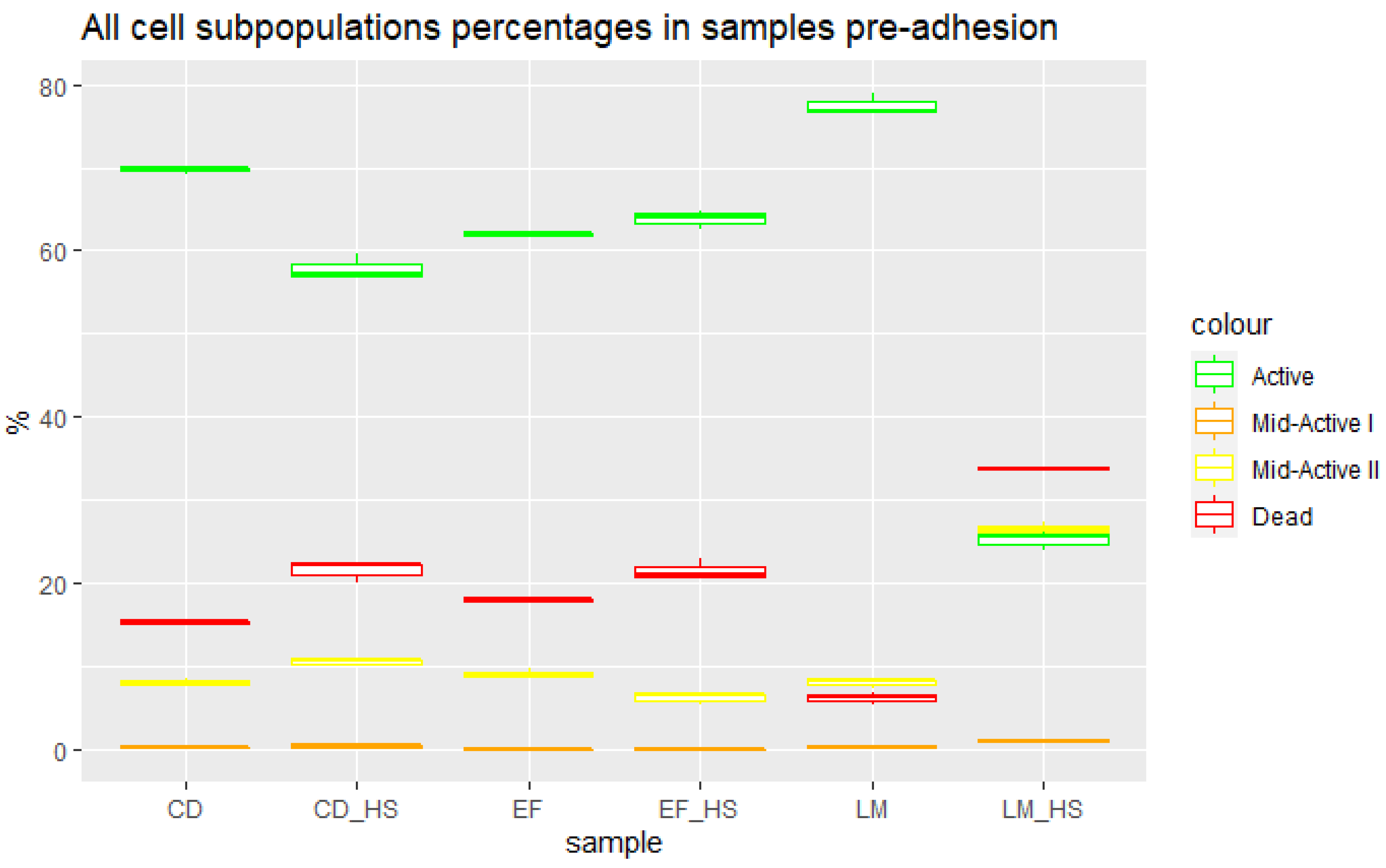
3.2. Cells Pre Adhesion
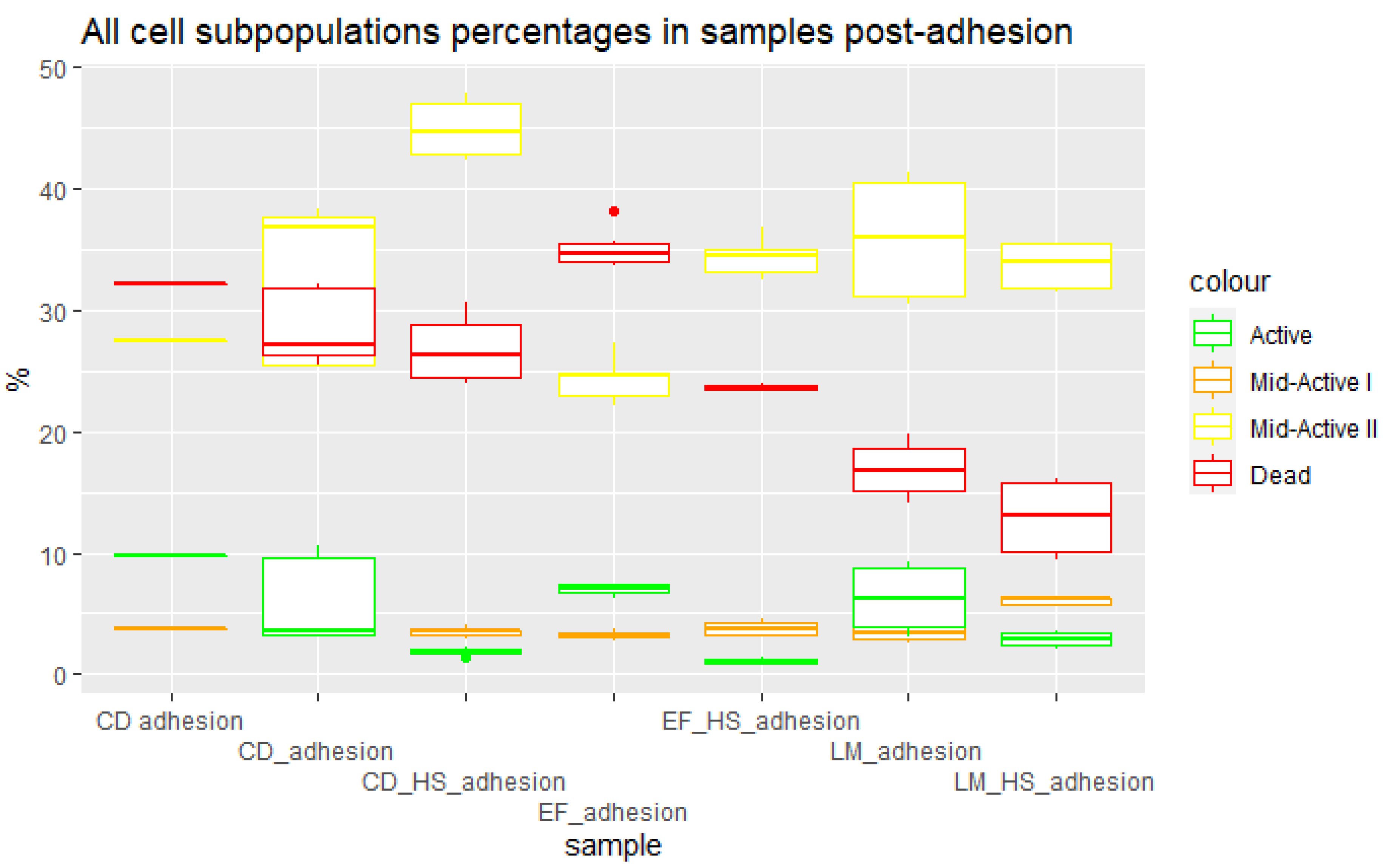
3.3. Cells Post Adhesion
3.4. Adherence Patterns and Membrane Staining with DRAQ-5
3.5. The Thermal Resistance of Bacteria
3.6. Analysis of Samples after Storage
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kiepś, J.; Dembczyński, R. Current Trends in the Production of Probiotic Formulations. Foods 2022, 11, 2330. [Google Scholar] [CrossRef]
- Islam, S.U. Clinical Uses of Probiotics. Medicine 2016, 95, e2658. [Google Scholar] [CrossRef]
- Prakoeswa, C.R.S.; Bonita, L.; Karim, A.; Herwanto, N.; Umborowati, M.A.; Setyaningrum, T.; Hidayati, A.N.; Surono, I.S. Beneficial Effect of Lactobacillus Plantarum IS-10506 Supplementation in Adults with Atopic Dermatitis: A Randomized Controlled Trial. J. Dermatol. Treat. 2022, 33, 1491–1498. [Google Scholar] [CrossRef]
- Sáez-Lara, M.J.; Robles-Sanchez, C.; Ruiz-Ojeda, F.J.; Plaza-Diaz, J.; Gil, A. Effects of Probiotics and Synbiotics on Obesity, Insulin Resistance Syndrome, Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease: A Review of Human Clinical Trials. Int. J. Mol. Sci. 2016, 17, 928. [Google Scholar] [CrossRef] [Green Version]
- Broeckx, G.; Vandenheuvel, D.; Claes, I.J.J.; Lebeer, S.; Kiekens, F. Drying Techniques of Probiotic Bacteria as an Important Step towards the Development of Novel Pharmabiotics. Int. J. Pharm. 2016, 505, 303–318. [Google Scholar] [CrossRef]
- Alp, D.; Kuleaşan, H. Adhesion Mechanisms of Lactic Acid Bacteria: Conventional and Novel Approaches for Testing. World J. Microbiol. Biotechnol. 2019, 35, 156. [Google Scholar] [CrossRef]
- Byakika, S.; Mukisa, I.M.; Byaruhanga, Y.B.; Muyanja, C. A Review of Criteria and Methods for Evaluating the Probiotic Potential of Microorganisms. Food Rev. Int. 2019, 35, 427–466. [Google Scholar] [CrossRef]
- Sarkar, S. Whether Viable and Dead Probiotic Are Equally Efficacious? NFS 2018, 48, 285–300. [Google Scholar] [CrossRef]
- Kulkarni, S.; Haq, S.F.; Samant, S.; Sukumaran, S. Adaptation of Lactobacillus Acidophilus to Thermal Stress Yields a Thermotolerant Variant Which Also Exhibits Improved Survival at PH 2. Probiotics Antimicro. Prot. 2018, 10, 717–727. [Google Scholar] [CrossRef]
- Bommasamudram, J.; Muthu, A.; Devappa, S. Effect of Sub-Lethal Heat Stress on Viability of Lacticaseibacillus Casei N in Spray-Dried Powders. LWT 2022, 155, 112904. [Google Scholar] [CrossRef]
- Wang, L.-Q.; Zhao, F.; Liu, F.; Meng, X.-C. Live/Dead State Is Not the Factor Influencing Adhesion Ability of Bifidobacterium Animalis KLDS2.0603. J. Microbiol. 2013, 51, 584–589. [Google Scholar] [CrossRef]
- Kiepś, J.; Juzwa, W.; Dembczyński, R. Imaging Flow Cytometry Demonstrates Physiological and Morphological Diversity within Treated Probiotic Bacteria Groups. Int. J. Mol. Sci. 2023, 24, 6841. [Google Scholar] [CrossRef]
- Valeriano, V.D.; Bagon, B.B.; Balolong, M.P.; Kang, D.-K. Carbohydrate-Binding Specificities of Potential Probiotic Lactobacillus Strains in Porcine Jejunal (IPEC-J2) Cells and Porcine Mucin. J. Microbiol. 2016, 54, 510–519. [Google Scholar] [CrossRef]
- Nishiyama, K.; Sugiyama, M.; Mukai, T. Adhesion Properties of Lactic Acid Bacteria on Intestinal Mucin. Microorganisms 2016, 4, 34. [Google Scholar] [CrossRef]
- Gorreja, F.; Walker, W.A. The Potential Role of Adherence Factors in Probiotic Function in the Gastrointestinal Tract of Adults and Pediatrics: A Narrative Review of Experimental and Human Studies. Gut Microbes 2022, 14, 2149214. [Google Scholar] [CrossRef]
- Jessie Lau, L.Y.; Chye, F.Y. Antagonistic Effects of Lactobacillus Plantarum 0612 on the Adhesion of Selected Foodborne Enteropathogens in Various Colonic Environments. Food Control 2018, 91, 237–247. [Google Scholar] [CrossRef]
- Walsham, A.D.S.; MacKenzie, D.A.; Cook, V.; Wemyss-Holden, S.; Hews, C.L.; Juge, N.; Schüller, S. Lactobacillus Reuteri Inhibition of Enteropathogenic Escherichia Coli Adherence to Human Intestinal Epithelium. Front. Microbiol. 2016, 7, 244. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Teng, K.; Liu, G.; Liu, Y.; Zhang, J.; Zhang, X.; Zhang, M.; Tao, Y.; Zhong, J. Lactobacillus Reuteri HCM2 Protects Mice against Enterotoxigenic Escherichia Coli through Modulation of Gut Microbiota. Sci. Rep. 2018, 8, 17485. [Google Scholar] [CrossRef] [Green Version]
- Bron, P.A.; Kleerebezem, M.; Brummer, R.-J.; Cani, P.D.; Mercenier, A.; MacDonald, T.T.; Garcia-Ródenas, C.L.; Wells, J.M. Can Probiotics Modulate Human Disease by Impacting Intestinal Barrier Function? Br. J. Nutr. 2017, 117, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion Mechanisms Mediated by Probiotics and Prebiotics and Their Potential Impact on Human Health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.H.; Jeong, C.H.; Cheng, W.N.; Choi, Y.; Shin, D.M.; Lee, S.; Han, S.G. Quality Characteristics of Yogurts Fermented with Short-Chain Fatty Acid-Producing Probiotics and Their Effects on Mucin Production and Probiotic Adhesion onto Human Colon Epithelial Cells. J. Dairy Sci. 2021, 104, 7415–7425. [Google Scholar] [CrossRef]
- Mohanty, D.; Panda, S.; Kumar, S.; Ray, P. In Vitro Evaluation of Adherence and Anti-Infective Property of Probiotic Lactobacillus Plantarum DM 69 against Salmonella Enterica. Microb. Pathog. 2019, 126, 212–217. [Google Scholar] [CrossRef]
- Xiong, R.; Pan, D.; Wu, Z.; Guo, Y.; Zeng, X.; Lian, L. Structure and Immunomodulatory Activity of a Recombinant Mucus-Binding Protein of Lactobacillus Acidophilus. Future Microbiol. 2018, 13, 1731–1743. [Google Scholar] [CrossRef]
- Yadav, A.K.; Tyagi, A.; Kumar, A.; Saklani, A.C.; Grover, S.; Batish, V.K. Adhesion of Indigenous Lactobacillusplantarum to Gut Extracellular Matrix and Its Physicochemical Characterization. Arch. Microbiol. 2015, 197, 155–164. [Google Scholar] [CrossRef]
- Najafi, S.; Sotoodehnejadnematalahi, F.; Amiri, M.M.; Pourshafie, M.R.; Rohani, M. Decreased Mucosal Adhesion of Lactobacillus Species in Patients with Inflammatory Bowel Disease. Casp. J. Intern. Med. 2022, 13, 713–720. [Google Scholar] [CrossRef]
- Krausova, G.; Hynstova, I.; Svejstil, R.; Mrvikova, I.; Kadlec, R. Identification of Synbiotics Conducive to Probiotics Adherence to Intestinal Mucosa Using an In Vitro Caco-2 and HT29-MTX Cell Model. Processes 2021, 9, 569. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Zavala, L.; Carasi, P.; Trejo, S.A.; Bronsoms, S.; de los Ángeles Serradell, M.; Garrote, G.L.; Abraham, A.G. Simulated Gastrointestinal Conditions Increase Adhesion Ability of Lactobacillus Paracasei Strains Isolated from Kefir to Caco-2 Cells and Mucin. Food Res. Int. 2018, 103, 462–467. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Llamas, M.G.; Iraporda, C.; Dueñas, M.T.; Abraham, A.G.; Garrote, G.L. Impact of Growth Temperature on Exopolysaccharide Production and Probiotic Properties of Lactobacillus Paracasei Strains Isolated from Kefir Grains. Food Microbiol. 2018, 69, 212–218. [Google Scholar] [CrossRef]
- Delon, L.C.; Guo, Z.; Oszmiana, A.; Chien, C.-C.; Gibson, R.; Prestidge, C.; Thierry, B. A Systematic Investigation of the Effect of the Fluid Shear Stress on Caco-2 cells towards the Optimization of Epithelial Organ-on-Chip Models. Biomaterials 2019, 225, 119521. [Google Scholar] [CrossRef] [PubMed]
- Mays, Z.J.S.; Chappell, T.C.; Nair, N.U. Quantifying and Engineering Mucus Adhesion of Probiotics. ACS Synth. Biol. 2020, 9, 356–367. [Google Scholar] [CrossRef]
- Saeed, A.; Yasmin, A.; Baig, M.; Ahmed, M.A.; Farooqi, Z.-R. Streptococcus Lactarius MB622 and Streptococcus Salivarius MB620 Isolated from Human Milk Reduce Chemokine IL-8 Production in Response to TNF-α in Caco-2 Cell Line, an Exploratory Study. Cytokine 2023, 168, 156232. [Google Scholar] [CrossRef]
- Lepock, J.R. Measurement of Protein Stability and Protein Denaturation in Cells Using Differential Scanning Calorimetry. Methods 2005, 35, 117–125. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tg Measured by DSC | Tg Measured by TG/DTA |
|---|---|---|
| uncoated | 145.3 °C | 141.3 °C |
| Gum arabic | 144.8 °C | 156.2 °C |
| HPMC | 152.6 °C | 156.1 °C |
| shellac | 147.6 °C | 140.3 °C |
| Sample | Number of Cells Pre-adhesion [cfu/mL] | Adhered Cells [cfu/mL] | Adhered Cells [%] |
|---|---|---|---|
| L. mesenteroides dried | 5.75 × 108 + SD | 4.68 × 107 | 8.14 |
| L. mesenteroides coated | 5.20 × 108 | 2.88 × 107 | 5.54 |
| E. faecium dried | 9.25 × 108 | 3.16 × 108 | 34.12 |
| E. faecium coated | 6.55 × 108 | 1.50 × 108 | 22.96 |
| C. divergens dried | 4.00 × 108 | 3.64 × 107 | 9.10 |
| C. divergens coated | 1.5 × 108 | 1.96 × 107 | 13.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiepś, J.; Juzwa, W.; Olejnik, A.; Sip, A.; Tomaszewska-Gras, J.; Dembczyński, R. The Effects of Cellular Membrane Damage on the Long-Term Storage and Adhesion of Probiotic Bacteria in Caco-2 Cell Line. Nutrients 2023, 15, 3484. https://doi.org/10.3390/nu15153484
Kiepś J, Juzwa W, Olejnik A, Sip A, Tomaszewska-Gras J, Dembczyński R. The Effects of Cellular Membrane Damage on the Long-Term Storage and Adhesion of Probiotic Bacteria in Caco-2 Cell Line. Nutrients. 2023; 15(15):3484. https://doi.org/10.3390/nu15153484
Chicago/Turabian StyleKiepś, Jakub, Wojciech Juzwa, Anna Olejnik, Anna Sip, Jolanta Tomaszewska-Gras, and Radosław Dembczyński. 2023. "The Effects of Cellular Membrane Damage on the Long-Term Storage and Adhesion of Probiotic Bacteria in Caco-2 Cell Line" Nutrients 15, no. 15: 3484. https://doi.org/10.3390/nu15153484