
Glycation of Whey Proteins Increases the Ex Vivo Immune Response of Lymphocytes Sensitized to β-Lactoglobulin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
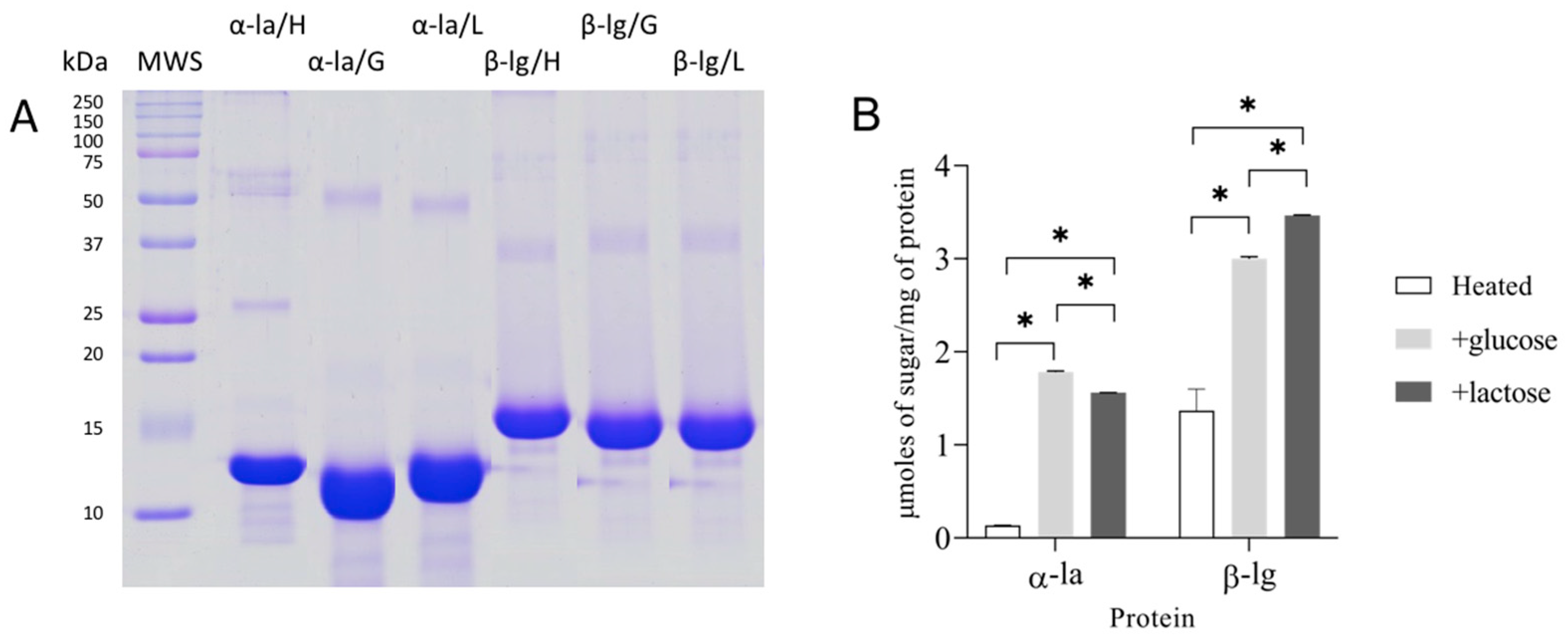
2.2. Glycation Protocol and Product Characterization
SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.3. In Vivo Experiment
2.3.1. Serum and Fecal Sample Preparation
2.3.2. Lymphocyte Isolation
- Complete Media (CM)—RPMI-1640 (G6784, Sigma-Aldrich) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 1 mM of nonessential amino acids, 1 mM of HEPES, 1 mM of sodium pyruvate, and 10 units/mL of penicillin–streptomycin solution.
- Incomplete Media (IM)—RPMI-1640 supplemented with 1 mM of HEPES, 1 mM of sodium pyruvate, and 10 units/mL of penicillin–streptomycin solution.
2.3.3. Isolation of Peripheral Blood Mononuclear Cells (PMBC)
2.3.4. Isolation of Lymphocytes from Tissues
2.4. Lymphocyte Culture
2.5. Lymphocyte Proliferation Assay
2.6. Lymphocyte Phenotyping
2.7. Determination of Specific Immunoglobulin Classes G and A in Serum Samples and Fecal Extracts
2.8. Cytokine Profile
2.9. Statistical Analysis
3. Results and Discussion
3.1. Glycated Proteins
3.2. Humoral Response against β–lg
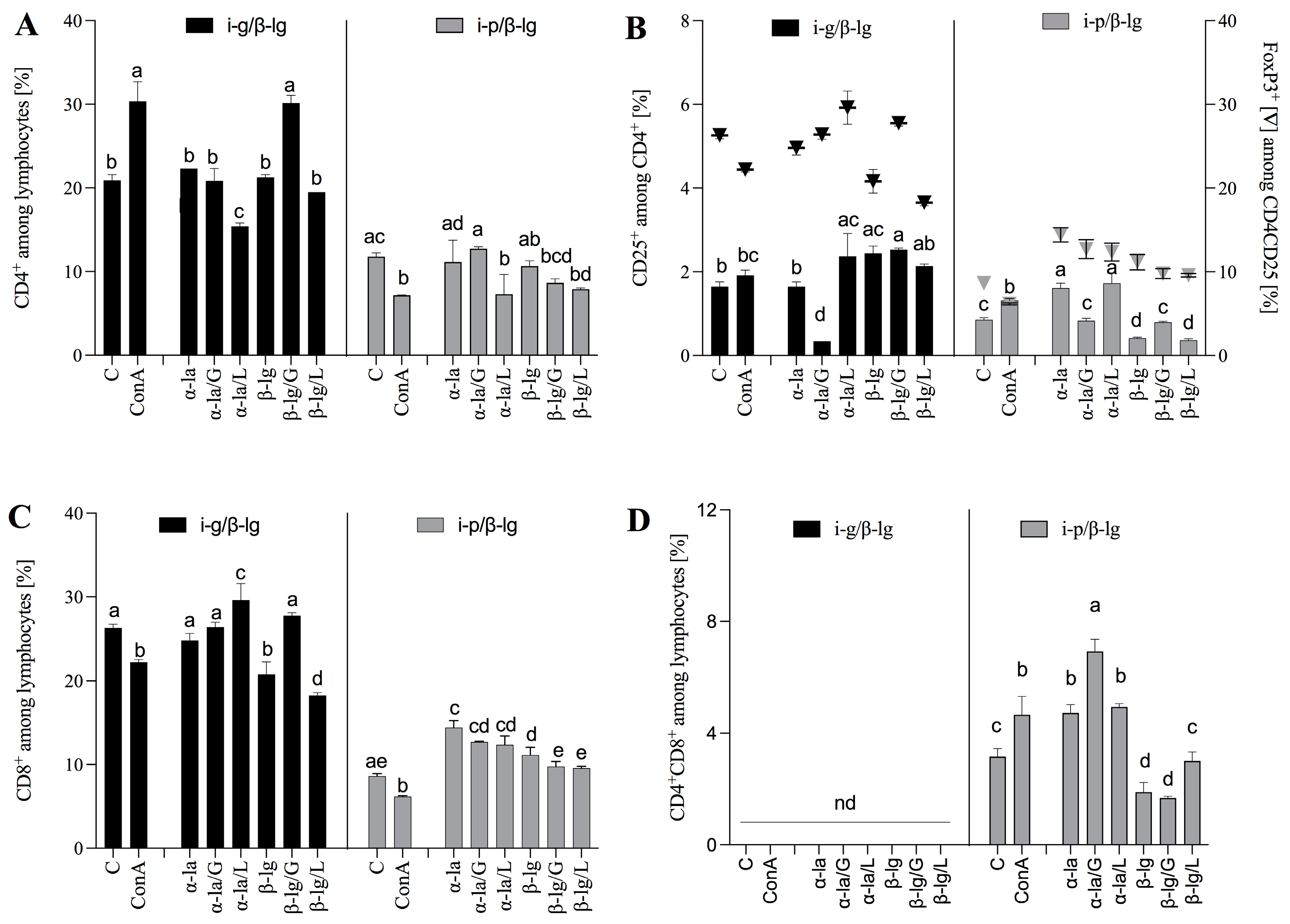
3.3. The Route of Antigen Exposure Modulates the Cellular Immune Response
3.4. Ex Vivo Lymphocyte Responses to α-la and β-lg

3.5. Cytokine Profiling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Złotkowska, D.; Stachurska, E.; Fuc, E.; Wróblewska, B.; Mikołajczyk, A.; Wasilewska, E. Differences in regulatory mechanisms induced by β-lactoglobulin and κ-casein in cow’s milk allergy mouse model-in vivo and ex vivo studies. Nutrients 2021, 13, 349. [Google Scholar] [CrossRef] [PubMed]
- Fuc, E.; Złotkowska, D.; Stachurska, E.; Wróblewska, B. Immunoreactive properties of alfa-casein and kappa-casein: Ex vivo and in vivo studies. J. Dairy Sci. 2018, 101, 10703–10713. [Google Scholar] [CrossRef] [Green Version]
- Fotschki, J.; Szyc, A.M.; Laparra, J.M.; Markiewicz, L.H.; Wróblewska, B. Immune-modulating properties of horse milk administered to mice sensitized to cow milk. J. Dairy Sci. 2016, 99, 9395–9404. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Ahmed, I.; Lv, L.; Li, Z.; Li, Z.; Lin, H.; Lin, H.; Zhao, J.; Tian, S.; Ma, J. Impacts of glycation and transglutaminase-catalyzed glycosylation with glucosamine on the conformational structure and allergenicity of bovine β-lactoglobulin. Food Funct. 2018, 9, 3944–3955. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Li, L.; Xu, D.; Sheng, B.; Chen, J.; Li, B.; Zhang, X. Heat-induced amyloid-like aggregation of β-lactoglobulin regulated by glycation: A comparison of five kinds of reducing saccharides. Int. J. Biol. Macromol. 2018, 120, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Milkovska-Stamenova, S.; Hoffmann, R. Influence of storage and heating on protein glycation levels of processed lactose-free and regular bovine milk products. Food Chem. 2017, 221, 489–495. [Google Scholar] [CrossRef]
- Chudzik-Kozłowska, J.; Wasilewska, E.; Złotkowska, D. Evaluation of Immunoreactivity of Pea (Pisum sativum) Albumins in BALB/c and C57BL/6 Mice. J. Agric. Food Chem. 2020, 68, 3891–3902. [Google Scholar] [CrossRef] [PubMed]
- Raupbach, J.; Ott, C.; Koenig, J.; Grune, T. Proteasomal degradation of glycated proteins depends on substrate unfolding: Preferred degradation of moderately modified myoglobin. Free Radic. Biol. Med. 2020, 152, 516–524. [Google Scholar] [CrossRef]
- Prasad, C.; Davis, K.E.; Imrhan, V.; Juma, S.; Vijayagopal, P. Advanced Glycation End Products and Risks for Chronic Diseases: Intervening Through Lifestyle Modification. Am. J. Lifestyle Med. 2019, 13, 384–404. [Google Scholar] [CrossRef]
- Zhang, M.; Zheng, J.; Ge, K.; Zhang, H.; Fang, B.; Jiang, L.; Guo, H.; Ding, Q.; Ren, F. Glycation of α-lactalbumin with different size saccharides: Effect on protein structure and antigenicity. Int. Dairy J. 2014, 34, 220–228. [Google Scholar] [CrossRef]
- Kobayashi, K.; Yoshida, T.; Takahashi, K.; Hattori, M. Modulation of the T cell response to β-lactoglobulin by conjugation with carboxymethyl dextran. Bioconjug. Chem. 2003, 14, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Iskandar, S.; Yoshida, T.; Takahashi, K.; Hattori, M. Reduced Immunogenicity of β-Lactoglobulin by Conjugating with Chitosan. Biosci. Biotechnol. Biochem. 2006, 70, 2349–2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, Q.; Jiang, X.; Li, Y.; Samiwala, M.; Labuza, T.P. Can Glycation Reduce Food Allergenicity? J. Agric. Food Chem. 2018, 66, 4295–4299. [Google Scholar] [CrossRef] [PubMed]
- Taheri-Kafrani, A.; Gaudin, J.C.; Rabesona, H.; Nioi, C.; Agarwal, D.; Drouet, M.; Chobert, J.M.; Bordbar, A.K.; Haertle, T. Effects of Heating and Glycation of β-Lactoglobulin on Its Recognition by IgE of Sera from Cow Milk Allergy Patients. J. Agric. Food Chem. 2009, 57, 4974–4982. [Google Scholar] [CrossRef]
- Xu, L.; Gong, Y.; Gern, J.E.; Ikeda, S.; Lucey, J.A. Glycation of whey protein with dextrans of different molar mass: Effect on immunoglobulin E-binding capacity with blood sera obtained from patients with cow milk protein allergy. J. Dairy Sci. 2018, 101, 6823–6834. [Google Scholar] [CrossRef] [Green Version]
- Enomoto, H.; Hayashi, Y.; Li, C.P.; Ohki, S.; Ohtomo, H.; Shiokawa, M.; Aoki, T. Glycation and phosphorylation of α-lactalbumin by dry heating: Effect on protein structure and physiological functions. J. Dairy Sci. 2009, 92, 3057–3068. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Tu, Z.C.; Liu, G.X.; Niu, C.D.; Yao, H.L.; Wang, H.; Sha, X.M.; Shao, Y.H.; Kaltashov, I.A. Ultrasonic Pretreatment Combined with Dry-State Glycation Reduced the Immunoglobulin E/Immunoglobulin G-Binding Ability of α-Lactalbumin Revealed by High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2018, 66, 5691–5698. [Google Scholar] [CrossRef]
- Yang, W.; Tu, Z.; Wang, H.; Zhang, L.; Xu, S.; Niu, C.; Yao, H.; Kaltashov, I.A. Mechanism of Reduction in IgG and IgE Binding of β-Lactoglobulin Induced by Ultrasound Pretreatment Combined with Dry-State Glycation: A Study Using Conventional Spectrometry and High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 8018–8027. [Google Scholar] [CrossRef]
- Deng, Y.; Govers, C.; Bastiaan-Net, S.; van der Hulst, N.; Hettinga, K.; Wichers, H.J. Hydrophobicity and aggregation, but not glycation, are key determinants for uptake of thermally processed β-lactoglobulin by THP-1 macrophages. Food Res. Int. 2019, 120, 102–113. [Google Scholar] [CrossRef]
- Li, J.L.; Liu, J.; Ye, Y.H.; Yang, P.; Tu, Z. cai Reduced IgE/IgG binding capacities of bovine α-Lactalbumin by glycation after dynamic high-pressure microfluidization pretreatment evaluated by high resolution mass spectrometry. Food Chem. 2019, 299, 125166. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Jansen, K.; Głobińska, A.; van de Veen, W.; Akdis, M. Regulatory Immune Mechanisms in Tolerance to Food Allergy. Front. Immunol. 2018, 9, 2939. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Furth, A.J. A microassay for protein glycation based on the periodate method. Anal. Biochem. 1991, 192, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Engholm-Keller, K.; Poojary, M.M.; Boelt, S.G.; Rogowska-Wrzesinska, A.; Skibsted, L.H.; Davies, M.J.; Lund, M.N. Generation of Aggregates of α-Lactalbumin by UV-B Light Exposure. J. Agric. Food Chem. 2020, 68, 6701–6714. [Google Scholar] [CrossRef]
- Li, R.; Lund, P.; Nielsen, S.B.; Lund, M.N. Formation of whey protein aggregates by partial hydrolysis and reduced thermal treatment. Food Hydrocoll. 2022, 124, 107206. [Google Scholar] [CrossRef]
- Gazi, I.; Franc, V.; Tamara, S.; van Gool, M.P.; Huppertz, T.; Heck, A.J.R. Identifying glycation hot-spots in bovine milk proteins during production and storage of skim milk powder. Int. Dairy J. 2022, 129, 105340. [Google Scholar] [CrossRef]
- Ledesma-Osuna, A.I.; Ramos-Clamont, G.; Vázquez-Moreno, L. Characterization of bovine serum albumin glycated with glucose, galactose and lactose. Acta Biochim. Polonica 2008, 55, 491–497. [Google Scholar] [CrossRef]
- Kanagaratham, C.; El Ansari, Y.S.; Lewis, O.L.; Oettgen, H.C. IgE and IgG Antibodies as Regulators of Mast Cell and Basophil Functions in Food Allergy. Front. Immunol. 2020, 11, 603050. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Torres, F.I.; Cabrera-Chávez, F.; Arvizu-Flores, A.A.; Flores-Mendoza, L.K.; Lopez-Teros, V.; Astiazaran-Garcia, H.; Gracia-Valenzuela, M.H.; Figueroa-Salcido, O.G.; Arámburo-Gálvez, J.G.; Ontiveros, N. Assessment of the Route of Exposure to Ovalbumin and Cow’s Milk Proteins on the Induction of IgE Responses in BALB/c Mice. Biology 2022, 11, 542. [Google Scholar] [CrossRef]
- Boyaka, P.N.; Fujihashi, K. Host Defenses at Mucosal Surfaces, 5th ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; ISBN 9780702068966. [Google Scholar]
- Littman, D.R.; Rudensky, A.Y. Th17 and Regulatory T Cells in Mediating and Restraining Inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Taniuchi, I. The CD4/CD8 lineages: Central decisions and peripheral modifications for T lymphocytes. Curr. Top. Microbiol. Immunol. 2014, 373, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mancera, M.S.; Bolaños, N.I.; Salamanca, M.; Orjuela, G.A.; Rodriguez, A.N.; Gonzalez, J.M. Percentages of cd4+cd8+ double-positive t lymphocytes in the peripheral blood of adults from a blood bank in bogotá, colombia. Turk. J. Hematol. 2020, 37, 36–41. [Google Scholar] [CrossRef]
- Pitmon, E.; Meehan, E.V.; Ahmadi, E.; Adler, A.J.; Wang, K. High glucose promotes regulatory T cell differ-entiation. PLoS ONE 2023, 18, e0280916. [Google Scholar] [CrossRef]
- Frischmeyer-Guerrerio, P.A.; Guerrerio, A.L.; Chichester, K.L.; Bieneman, A.P.; Hamilton, R.A.; Wood, R.A.; Schroeder, J.T. Dendritic cell and T cell responses in children with food allergy. Clin. Exp. Allergy 2011, 41, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, S.; Rincón, M. The two faces of IL-6 on Th1/Th2 differentiation. Mol. Immunol. 2002, 39, 531–536. [Google Scholar] [CrossRef]
- Packi, K.; Matysiak, J.; Klimczak, S.; Matuszewska, E.; Bręborowicz, A.; Pietkiewicz, D.; Matysiak, J. Analysis of the Serum Profile of Cytokines Involved in the T-Helper Cell Type 17 Immune Response Pathway in Atopic Children with Food Allergy. Int. J. Environ. Res. Public Health 2022, 19, 7877. [Google Scholar] [CrossRef]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The Biological Functions of T Helper 17 Cell Effector Cytokines in Inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dardalhon, V.; Korn, T.; Kuchroo, V.K.; Anderson, A.C. Role of Th1 and Th17 cells in organ-specific autoimmunity. J. Autoimmun. 2008, 31, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.Y.; Flavell, R.A. How Diverse—CD4 Effector T Cells and their Functions. J. Mol. Cell Biol. 2009, 2, 20–36. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | Group | Cytokine Concentration [pg/mL] in Media after Stimulation of Cells with | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | ConA | α-la | α-la/G | α-la/L | β-lg | β-lg/G | β-lg/L | ||
| IL-10 | i-g/β-lg | 0.00 c | 175.69 ± 11.04 *,a | 54.72 ± 8.83 *,b | 0.00 c | 0.00 c | 413.68 ± 44.69 *,d | 0.00 c | 0.00 c |
| i-p/β-lg | 0.00 c | 72 ± 8.26 b | 265.28 ± 59.05 a | 0.00 c | 0.00 c | 583.76 ± 14.99 d | 0.00 c | 0.00 c | |
| IL-17A | i-g/β-lg | 237.73 ± 22.81 *,c,f,g | 694.3 ± 81.72 *,a,d | 490.52 ± 22.54 *,b | 284.14 ± 0.58 *,c,e,g | 753.53 ± 76.06 *,d | 196.48 ± 42.51 *,e,f,g | 282.26 ± 19.92 *,g | 182.97 ± 9.98 f |
| i-p/β-lg | 11.16 ± 1.48 b,c,d,e | 340.12 ± 28.11 a | 73.59 ± 9.64 b,d,e,f | 112.78 ± 6.75 d | 63.5 ± 1.54 b | 21.95 ± 1.73 e,f | 14.25 ± 2.15 e | 135.64 ± 4.25 d | |
| TNF-α | i-g/β-lg | 15 ± 3.46 c | 1150 ± 118.93 *,a | 582 ± 42.15 *,b | 172 ± 4.04 d,e | 96 ± 3.05 e | 193 ± 26.90 *,e,d | 142 ± 4.89 e | 109 ± 29.84 e |
| i-p/β-lg | 91 ± 0.88 c | 798 ± 29.39 a | 1705 ± 339.82 b | 144 ± 18.10 c | 146 ± 11.47 e | 1656 ± 240.31 b | 182 ± 6.27 c | 149 ± 1.15 c | |
| IFN-ɣ | i-g/β-lg | 0.00 b,c | 348.93 ± 24.14 *,a | 30.56 ± 0.82 *,b | 0.00 c | 0.00 c | 11.83 ± 0.63 *,c | 1.80 ± 0.36 c | 0.00 *,c |
| i-p/β-lg | 30.57 ± 0.29 c | 2972.76 ± 218.64 a | 610.89 ± 110.07 b | 8.18 ± 0.11 c | 10.85 ± 0.25 c | 672.13 ± 48.68 b | 16.85 ± 0.49 c | 14.05 ± 0.28 c | |
| IL-6 | i-g/β-lg | 12.38 ± 2.22 c | 2387.12 ± 245.45 *,a | 285.0 ± 36.96 *,b | 58.30 ± 0.39 d | 31.04 ± 6.57 d | 85.38 ± 29.06 *,d | 48.81 ± 3.14 d | 43.32 ± 7.59 d |
| i-p/β-lg | 7.21 ± 0.97 c | 3331.01 ± 192.48 a | 904.66 ± 177.38 b | 9.59 ± 1.91 c | 9.50 ± 2.03 c | 1052.54 ± 181.81 b | 21.53 ± 0.98 c | 11.95 ± 2.15 c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Złotkowska, D.; Kuczyński, M.; Fuc, E.; Fotschki, J.; Wróblewska, B. Glycation of Whey Proteins Increases the Ex Vivo Immune Response of Lymphocytes Sensitized to β-Lactoglobulin. Nutrients 2023, 15, 3110. https://doi.org/10.3390/nu15143110
Złotkowska D, Kuczyński M, Fuc E, Fotschki J, Wróblewska B. Glycation of Whey Proteins Increases the Ex Vivo Immune Response of Lymphocytes Sensitized to β-Lactoglobulin. Nutrients. 2023; 15(14):3110. https://doi.org/10.3390/nu15143110
Chicago/Turabian StyleZłotkowska, Dagmara, Mateusz Kuczyński, Ewa Fuc, Joanna Fotschki, and Barbara Wróblewska. 2023. "Glycation of Whey Proteins Increases the Ex Vivo Immune Response of Lymphocytes Sensitized to β-Lactoglobulin" Nutrients 15, no. 14: 3110. https://doi.org/10.3390/nu15143110