
Metabolomic Profiling in Children with Celiac Disease: Beyond the Gluten-Free Diet
 , , , , , and
, , , , , and
Abstract
:1. Introduction
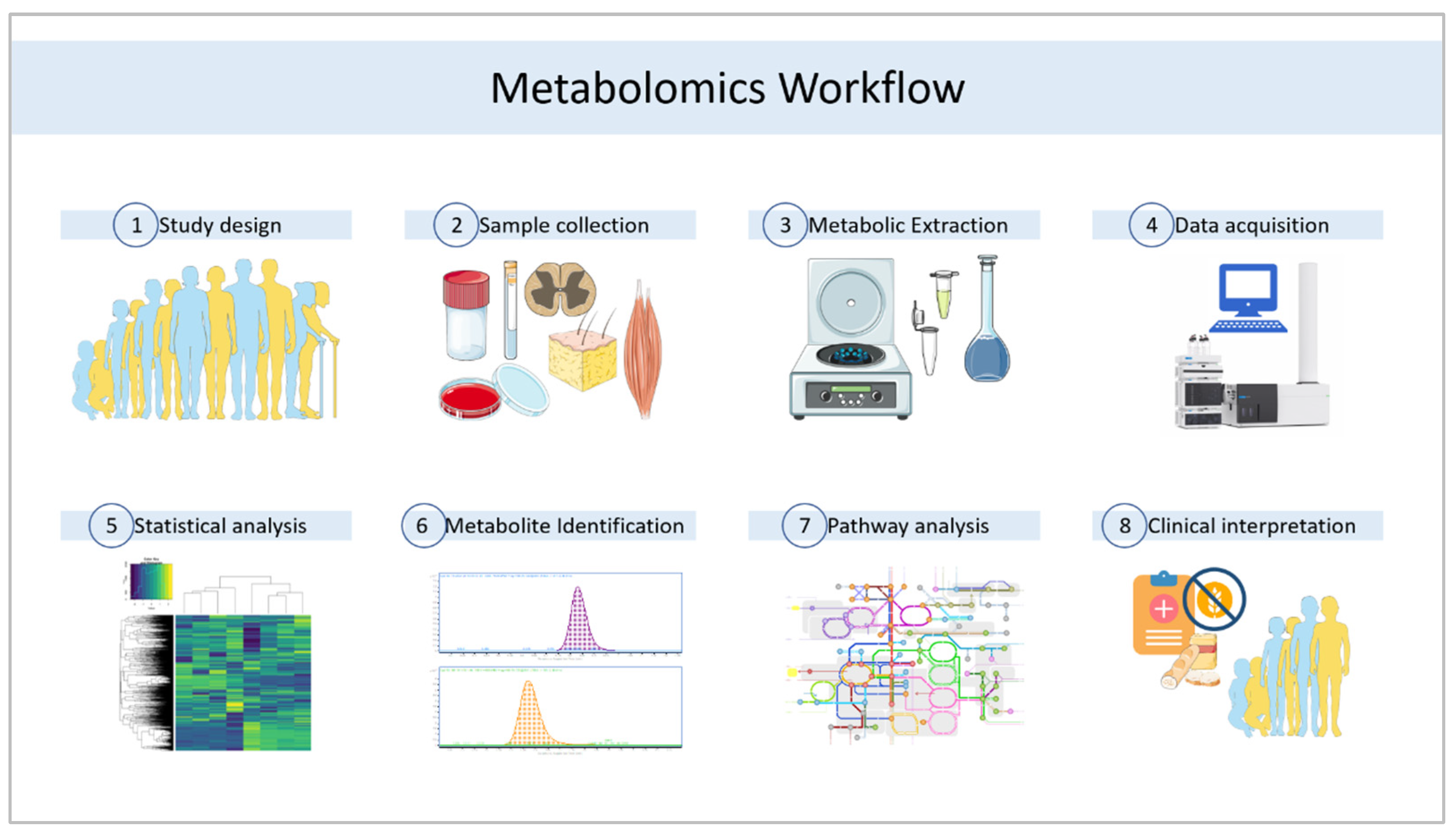
2. Materials and Methods
3. Metabolomics Platforms
4. Plasma Metabolomic Profile
4.1. CD’s Inherent Footprint and Role of the GFD
4.2. Genetic Influence (HLA)
5. Urine Metabolomic Profile
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martín-Masot, R.; Diaz-Castro, J.; Moreno-Fernandez, J.; Navas-López, V.M.; Nestares, T. The Role of Early Programming and Early Nutrition on the Development and Progression of Celiac Disease: A Review. Nutrients 2020, 12, 3427. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsson, J.F.; Bai, J.C.; Biagi, F.; Card, T.R.; Ciacci, C.; Ciclitira, P.J.; Green, P.H.R.; Hadjivassiliou, M.; Holdoway, A.; Van Heel, D.A.; et al. Diagnosis and Management of Adult Coeliac Disease: Guidelines from the British Society of Gastroenterology. Gut 2014, 63, 1210–1228. [Google Scholar] [CrossRef] [Green Version]
- Szajewska, H.; Shamir, R.; Mearin, L.; Ribes-Koninckx, C.; Catassi, C.; Domellof, M.; Fewtrell, M.S.; Husby, S.; Papadopoulou, A.; Vandenplas, Y.; et al. Gluten Introduction and the Risk of Coeliac Disease: A Position Paper by the European Society for Pediatric Gastroenterology, Hepatology, and Nutrition. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Lebwohl, B.; Sanders, D.S.; Green, P.H.R. Coeliac Disease. Lancet 2018, 391, 70–81. [Google Scholar] [CrossRef]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.A.; De Giorgio, R.; Catassi, C.; Fasano, A. Celiac Disease: A Comprehensive Current Review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Leonard, M.M.; Karathia, H.; Pujolassos, M.; Troisi, J.; Valitutti, F.; Subramanian, P.; Camhi, S.; Kenyon, V.; Colucci, A.; Serena, G.; et al. Multi-Omics Analysis Reveals the Influence of Genetic and Environmental Risk Factors on Developing Gut Microbiota in Infants at Risk of Celiac Disease. Microbiome 2020, 8, 130. [Google Scholar] [CrossRef]
- King, J.A.; Jeong, J.; Underwood, F.E.; Quan, J.; Panaccione, N.; Windsor, J.W.; Coward, S.; Debruyn, J.; Ronksley, P.E.; Shaheen, A.A.; et al. Incidence of Celiac Disease Is Increasing over Time: A Systematic Review and Meta-Analysis. Am. J. Gastroenterol. 2020, 115, 507–525. [Google Scholar] [CrossRef]
- Abrams, J.A.; Diamond, B.; Rotterdam, H.; Green, P.H.R. Seronegative Celiac Disease: Increased Prevalence with Lesser Degrees of Villous Atrophy. Dig. Dis. Sci. 2004, 49, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, R.P.G. Diagnosing Coeliac Disease: Out with the Old and in with the New? World J. Gastroenterol. 2020, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.M.; Ciaccio, E.J.; Case, S.; Jaffe, N.; Mahadov, S.; Lebwohl, B.; Green, P.H. Celiac Disease in Patients with Type 1 Diabetes: Screening and Diagnostic Practices. Diabetes Educ. 2013, 39, 532–540. [Google Scholar] [CrossRef]
- Volta, U.; Rostami, K.; Tovoli, F.; Caio, G.; Masi, C.; Ruggeri, E.; Cacciari, G.; Bon, I.; De Giorgio, R. Fulminant Type 1 Autoimmune Hepatitis in a Recently Diagnosed Celiac Disease Patient. Arch. Iran Med. 2013, 16, 683–685. [Google Scholar] [PubMed]
- Martín-Masot, R.; Herrador-López, M.; Navas-López, V.M.; Carmona, F.D.; Nestares, T.; Bossini-Castillo, L. Celiac Disease Is a Risk Factor for Mature T and NK Cell Lymphoma: A Mendelian Randomization Study. Int. J. Mol. Sci. 2023, 24, 7216. [Google Scholar] [CrossRef]
- Diaz-Castro, J.; Muriel-Neyra, C.; Martin-Masot, R.; Moreno-Fernandez, J.; Maldonado, J.; Nestares, T. Oxidative Stress, DNA Stability and Evoked Inflammatory Signaling in Young Celiac Patients Consuming a Gluten-Free Diet. Eur. J. Nutr. 2019, 59, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Nestares, T.; Martín-Masot, R.; Flor-Alemany, M.; Bonavita, A.; Maldonado, J.; Aparicio, V.A. Influence of Ultra-Processed Foods Consumption on Redox Status and Inflammatory Signaling in Young Celiac Patients. Nutrients 2021, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Nestares, T.; Martín-Masot, R.; Labella, A.; Aparicio, V.A.; Flor-Alemany, M.; López-Frías, M.; Maldonado, J. Is a Gluten-Free Diet Enough to Maintain Correct Micronutrients Status in Young Patients with Celiac Disease? Nutrients 2020, 12, 844. [Google Scholar] [CrossRef] [Green Version]
- Silano, M.; Volta, U.; Mecchia, A.M.; Dessì, M.; Di Benedetto, R.; De Vincenzi, M. Delayed Diagnosis of Coeliac Disease Increases Cancer Risk. BMC Gastroenterol. 2007, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Viljamaa, M.; Kaukinen, K.; Pukkala, E.; Hervonen, K.; Reunala, T.; Collin, P. Malignancies and Mortality in Patients with Coeliac Disease and Dermatitis Herpetiformis: 30-Year Population-Based Study. Dig. Liver Dis. 2006, 38, 374–380. [Google Scholar] [CrossRef]
- Mearin, M.L.; Catassi, C.; Brousse, N.; Brand, R.; Collin, P.; Fabiani, E.; Schweizer, J.J.; Abuzakouk, M.; Szajewska, H.; Hallert, C.; et al. European Multi-Centre Study on Coeliac Disease and Non-Hodgkin Lymphoma. Eur. J. Gastroenterol. Hepatol. 2006, 18, 187–194. [Google Scholar] [CrossRef]
- Ilus, T.; Kaukinen, K.; Virta, L.J.; Pukkala, E.; Collin, P. Incidence of Malignancies in Diagnosed Celiac Patients: A Population-Based Estimate. Am. J. Gastroenterol. 2014, 109, 1471–1477. [Google Scholar] [CrossRef]
- Kell, D.B.; Goodacre, R. Metabolomics and Systems Pharmacology: Why and How to Model the Human Metabolic Network for Drug Discovery. Drug Discov. Today 2014, 19, 171–182. [Google Scholar] [CrossRef]
- Allen, J.; Davey, H.M.; Broadhurst, D.; Heald, J.K.; Rowland, J.J.; Oliver, S.G.; Kell, D.B. High-Throughput Classification of Yeast Mutants for Functional Genomics Using Metabolic Footprinting. Nat. Biotechnol. 2003, 21, 692–696. [Google Scholar] [CrossRef]
- Sellitto, M.; Bai, G.; Serena, G.; Fricke, W.F.; Sturgeon, C.; Gajer, P.; White, J.R.; Koenig, S.S.K.; Sakamoto, J.; Boothe, D.; et al. Proof of Concept of Microbiome-Metabolome Analysis and Delayed Gluten Exposure on Celiac Disease Autoimmunity in Genetically at-Risk Infants. PLoS ONE 2012, 7, e33387. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. “Metabonomics”: Understanding the Metabolic Responses of Living Systems to Pathophysiological Stimuli via Multivariate Statistical Analysis of Biological NMR Spectroscopic Data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, Nontargeted Ultrahigh Performance Liquid Chromatography/Electrospray Ionization Tandem Mass Spectrometry Platform for the Identification and Relative Quantification of the Small-Molecule Complement of Biological Systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B. Current Trends and Future Requirements for the Mass Spectrometric Investigation of Microbial, Mammalian and Plant Metabolomes. Phys. Biol. 2008, 5, 011001. [Google Scholar] [CrossRef] [PubMed]
- Kan, M.; Himes, B.E. Insights into Glucocorticoid Responses Derived from Omics Studies. Pharmacol. Ther. 2021, 218, 107674. [Google Scholar] [CrossRef] [PubMed]
- Bordag, N.; Klie, S.; Jürchott, K.; Vierheller, J.; Schiewe, H.; Albrecht, V.; Tonn, J.-C.; Schwartz, C.; Schichor, C.; Selbig, J. Glucocorticoid (Dexamethasone)-Induced Metabolome Changes in Healthy Males Suggest Prediction of Response and Side Effects. Sci. Rep. 2015, 5, 15954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S. Emerging Applications of Metabolomics in Drug Discovery and Precision Medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Petrick, L.; Edmands, W.; Schiffman, C.; Grigoryan, H.; Perttula, K.; Yano, Y.; Dudoit, S.; Whitehead, T.; Metayer, C.; Rappaport, S. An Untargeted Metabolomics Method for Archived Newborn Dried Blood Spots in Epidemiologic Studies. Metabolomics 2017, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiecki, M.M.; Walker, D.I.; Vermeulen, R.; Chadeau-Hyam, M.; Jones, D.P.; Miller, G.W. The Exposome: Molecules to Populations. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 107–127. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Yu, T.; Strobel, F.H.; Jones, D.P. A Practical Approach to Detect Unique Metabolic Patterns for Personalized Medicine. Analyst 2010, 135, 2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabré, R.; Jové, M.; Naudí, A.; Ayala, V.; Piñol-Ripoll, G.; Gil-Villar, M.P.; Dominguez-Gonzalez, M.; Obis, È.; Berdun, R.; Mota-Martorell, N.; et al. Specific Metabolomics Adaptations Define a Differential Regional Vulnerability in the Adult Human Cerebral Cortex. Front. Mol. Neurosci. 2016, 9, 138. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Sparks, L.M. Metabolic Flexibility in Health and Disease. Cell Metab. 2017, 25, 1027–1036. [Google Scholar] [CrossRef] [Green Version]
- Moreau, R.; Clària, J.; Aguilar, F.; Fenaille, F.; Lozano, J.J.; Junot, C.; Colsch, B.; Caraceni, P.; Trebicka, J.; Pavesi, M.; et al. Blood Metabolomics Uncovers Inflammation-Associated Mitochondrial Dysfunction as a Potential Mechanism Underlying ACLF. J. Hepatol. 2020, 72, 688–701. [Google Scholar] [CrossRef]
- Pang, H.; Jia, W.; Hu, Z. Emerging Applications of Metabolomics in Clinical Pharmacology. Clin. Pharmacol. Ther. 2019, 106, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wang, X. Power of Metabolomics in Biomarker Discovery and Mining Mechanisms of Obesity. Obes. Rev. 2013, 14, 344–349. [Google Scholar] [CrossRef]
- Thomson, T.M.; Balcells, C.; Cascante, M. Metabolic Plasticity and Epithelial-Mesenchymal Transition. J. Clin. Med. 2019, 8, 967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saner, C.; Harcourt, B.E.; Pandey, A.; Ellul, S.; McCallum, Z.; Kao, K.-T.; Twindyakirana, C.; Pons, A.; Alexander, E.J.; Saffery, R.; et al. Sex and Puberty-Related Differences in Metabolomic Profiles Associated with Adiposity Measures in Youth with Obesity. Metabolomics 2019, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Handelman, S.K.; Romero, R.; Tarca, A.L.; Pacora, P.; Ingram, B.; Maymon, E.; Chaiworapongsa, T.; Hassan, S.S.; Erez, O. The Plasma Metabolome of Women in Early Pregnancy Differs from That of Non-Pregnant Women. PLoS ONE 2019, 14, e224682. [Google Scholar] [CrossRef] [PubMed]
- Christian, P.; Smith, E.R.; Lee, S.E.; Vargas, A.J.; Bremer, A.A.; Raiten, D.J. The Need to Study Human Milk as a Biological System. Am. J. Clin. Nutr. 2021, 113, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Mota-Martorell, N.; Jové, M.; Borrás, C.; Berdún, R.; Obis, È.; Sol, J.; Cabré, R.; Pradas, I.; Galo-Licona, J.D.; Puig, J.; et al. Methionine Transsulfuration Pathway Is Upregulated in Long-Lived Humans. Free Radic. Biol. Med. 2021, 162, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Sol, J.; Obis, È.; Mota-Martorell, N.; Pradas, I.; Galo-Licona, J.D.; Martin-Garí, M.; Fernández-Bernal, A.; Ortega-Bravo, M.; Mayneris-Perxachs, J.; Borrás, C.; et al. Plasma Acylcarnitines and Gut-derived Aromatic Amino Acids as Sex-specific Hub Metabolites of the Human Aging Metabolome. Aging Cell 2023, 22, e13821. [Google Scholar] [CrossRef] [PubMed]
- Sas, K.M.; Karnovsky, A.; Michailidis, G.; Pennathur, S. Metabolomics and Diabetes: Analytical and Computational Approaches. Diabetes 2015, 64, 718–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang-Sattler, R.; Yu, Z.; Herder, C.; Messias, A.C.; Floegel, A.; He, Y.; Heim, K.; Campillos, M.; Holzapfel, C.; Thorand, B.; et al. Novel Biomarkers for Pre-diabetes Identified by Metabolomics. Mol. Syst. Biol. 2012, 8, 615. [Google Scholar] [CrossRef]
- Pallares-Méndez, R.; Aguilar-Salinas, C.A.; Cruz-Bautista, I.; del Bosque-Plata, L. Metabolomics in Diabetes, a Review. Ann. Med. 2016, 48, 89–102. [Google Scholar] [CrossRef]
- Danzi, F.; Pacchiana, R.; Mafficini, A.; Scupoli, M.T.; Scarpa, A.; Donadelli, M.; Fiore, A. To Metabolomics and beyond: A Technological Portfolio to Investigate Cancer Metabolism. Signal Transduct. Target Ther. 2023, 8, 137. [Google Scholar] [CrossRef]
- Gatius, S.; Jove, M.; Megino-Luque, C.; Albertí-Valls, M.; Yeramian, A.; Bonifaci, N.; Piñol, M.; Santacana, M.; Pradas, I.; Llobet-Navas, D.; et al. Metabolomic Analysis Points to Bioactive Lipid Species and Acireductone Dioxygenase 1 (ADI1) as Potential Therapeutic Targets in Poor Prognosis Endometrial Cancer. Cancers 2022, 14, 2842. [Google Scholar] [CrossRef]
- Shao, Y.; Le, W. Recent Advances and Perspectives of Metabolomics-Based Investigations in Parkinson’s Disease. Mol. Neurodegener. 2019, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Pamplona, R.; Obis, E.; Sol, J.; Andres-Benito, P.; Martí n-Gari, M.; Mota-Martorell, N.; Daniel Galo-Licona, J.; Piñol-Ripoll, G.; Portero-Otin, M.; Ferrer, I.; et al. Lipidomic Alterations in the Cerebral Cortex and White Matter in Sporadic Alzheimer’s Disease. bioRxiv 2023. [Google Scholar] [CrossRef]
- Centanni, M.; Moes, D.J.A.R.; Trocóniz, I.F.; Ciccolini, J.; van Hasselt, J.G.C. Clinical Pharmacokinetics and Pharmacodynamics of Immune Checkpoint Inhibitors. Clin. Pharmacokinet. 2019, 58, 835–857. [Google Scholar] [CrossRef] [Green Version]
- Alarcon-Barrera, J.C.; Kostidis, S.; Ondo-Mendez, A.; Giera, M. Recent Advances in Metabolomics Analysis for Early Drug Development. Drug Discov. Today 2022, 27, 1763–1773. [Google Scholar] [CrossRef]
- De Castro, F.; Benedetti, M.; Del Coco, L.; Fanizzi, F.P. NMR-Based Metabolomics in Metal-Based Drug Research. Molecules 2019, 24, 2240. [Google Scholar] [CrossRef] [Green Version]
- Lindon, J.C.; Holmes, E.; Nicholson, J.K. Metabonomics in Pharmaceutical R & D. FEBS J. 2007, 274, 1140–1151. [Google Scholar]
- Uppal, K.; Walker, D.I.; Liu, K.; Li, S.; Go, Y.-M.; Jones, D.P. Computational Metabolomics: A Framework for the Million Metabolome. Chem. Res. Toxicol. 2016, 29, 1956–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, D.; Lee, M.; Kim, S.; Kim, S. Applications of NMR Spectroscopy Based Metabolomics: A Review. J. Korean Magn. Reson. Soc. 2013, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.U.; Shariff, M.I.F.; Crossey, M.M.E.; Gomez-Romero, M.; Holmes, E.; Cox, I.J.; Fye, H.K.S.; Njie, R.; Taylor-Robinson, S.D. Hepatocellular Carcinoma: Review of Disease and Tumor Biomarkers. World J. Hepatol. 2016, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- Emwas, A.-H.M. The Strengths and Weaknesses of NMR Spectroscopy and Mass Spectrometry with Particular Focus on Metabolomics Research. In Metabonomics: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2015; pp. 161–193. [Google Scholar]
- Adamski, J. Introduction to Metabolomics. In Metabolomics for Biomedical Research; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–15. [Google Scholar]
- Bhinderwala, F.; Wase, N.; DiRusso, C.; Powers, R. Combining Mass Spectrometry and NMR Improves Metabolite Detection and Annotation. J. Proteome Res. 2018, 17, 4017–4022. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Keun, H.C.; Athersuch, T.J. Nuclear Magnetic Resonance (NMR)-Based Metabolomics. In Metabolic Profiling: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2011; pp. 321–334. [Google Scholar]
- Letertre, M.P.M.; Dervilly, G.; Giraudeau, P. Combined Nuclear Magnetic Resonance Spectroscopy and Mass Spectrometry Approaches for Metabolomics. Anal. Chem. 2021, 93, 500–518. [Google Scholar] [CrossRef] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The Human Serum Metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [Green Version]
- Junot, C.; Fenaille, F.; Colsch, B.; Bécher, F. High Resolution Mass Spectrometry Based Techniques at the Crossroads of Metabolic Pathways. Mass Spectrom. Rev. 2014, 33, 471–500. [Google Scholar] [CrossRef] [PubMed]
- Yanes, O.; Tautenhahn, R.; Patti, G.J.; Siuzdak, G. Expanding Coverage of the Metabolome for Global Metabolite Profiling. Anal. Chem. 2011, 83, 2152–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piraud, M.; Vianey-Saban, C.; Petritis, K.; Elfakir, C.; Steghens, J.; Morla, A.; Bouchu, D. ESI-MS/MS Analysis of Underivatised Amino Acids: A New Tool for the Diagnosis of Inherited Disorders of Amino Acid Metabolism. Fragmentation Study of 79 Molecules of Biological Interest in Positive and Negative Ionisation Mode. Rapid Commun. Mass Spectrom. 2003, 17, 1297–1311. [Google Scholar] [CrossRef]
- Bruins, A.P. Mass spectrometry—Atmospheric Pressure Ionization Techniques. In Encyclopedia of Analytical Science; Elsevier: Amsterdam, The Netherlands, 2005; pp. 366–373. [Google Scholar]
- Schuhmacher, J.; Zimmer, D.; Tesche, F.; Pickard, V. Matrix Effects during Analysis of Plasma Samples by Electrospray and Atmospheric Pressure Chemical Ionization Mass Spectrometry: Practical Approaches to Their Elimination. Rapid Commun. Mass Spectrom. 2003, 17, 1950–1957. [Google Scholar] [CrossRef]
- Karas, M.; Krüger, R. Ion Formation in MALDI: The Cluster Ionization Mechanism. Chem. Rev. 2003, 103, 427–440. [Google Scholar] [CrossRef]
- Theodoridis, G.; Gika, H.G.; Wilson, I.D. Mass Spectrometry-Based Holistic Analytical Approaches for Metabolite Profiling in Systems Biology Studies. Mass Spectrom. Rev. 2011, 30, 884–906. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global Metabolic Profiling of Animal and Human Tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Špánik, I.; Machyňáková, A. Recent Applications of Gas Chromatography with High-Resolution Mass Spectrometry. J. Sep. Sci. 2018, 41, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Zeki, Ö.C.; Eylem, C.C.; Reçber, T.; Kır, S.; Nemutlu, E. Integration of GC–MS and LC–MS for Untargeted Metabolomics Profiling. J. Pharm. Biomed. Anal. 2020, 190, 113509. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.D.; Powers, R. Beyond the Paradigm: Combining Mass Spectrometry and Nuclear Magnetic Resonance for Metabolomics. Prog. Nucl. Magn. Reason. Spectrosc. 2017, 100, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martias, C.; Baroukh, N.; Mavel, S.; Blasco, H.; Lefèvre, A.; Roch, L.; Montigny, F.; Gatien, J.; Schibler, L.; Dufour-Rainfray, D.; et al. Optimization of Sample Preparation for Metabolomics Exploration of Urine, Feces, Blood and Saliva in Humans Using Combined NMR and UHPLC-HRMS Platforms. Molecules 2021, 26, 4111. [Google Scholar] [CrossRef] [PubMed]
- Vuckovic, D. Sample Preparation in Global Metabolomics of Biological Fluids and Tissues. In Proteomic and Metabolomic Approaches to Biomarker Discovery; Elsevier: Amsterdam, The Netherlands, 2013; pp. 51–75. [Google Scholar]
- Lu, W.; Bennett, B.D.; Rabinowitz, J.D. Analytical Strategies for LC–MS-Based Targeted Metabolomics. J. Chromatogr. B 2008, 871, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Patti, G.J.; Tautenhahn, R.; Siuzdak, G. Meta-Analysis of Untargeted Metabolomic Data from Multiple Profiling Experiments. Nat. Protoc. 2012, 7, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Jacob, M.; Malkawi, A.; Albast, N.; Al Bougha, S.; Lopata, A.; Dasouki, M.; Abdel Rahman, A.M. A Targeted Metabolomics Approach for Clinical Diagnosis of Inborn Errors of Metabolism. Anal. Chim. Acta 2018, 1025, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Puigarnau, S.; Fernàndez, A.; Obis, E.; Jové, M.; Castañer, M.; Pamplona, R.; Portero-Otin, M.; Camerino, O. Metabolomics Reveals That Fittest Trail Runners Show a Better Adaptation of Bioenergetic Pathways. J. Sci. Med. Sport 2022, 25, 6. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for Large-Scale Metabolic Profiling of Serum and Plasma Using Gas Chromatography and Liquid Chromatography Coupled to Mass Spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Molnos, S.; Wahl, S.; Haid, M.; Eekhoff, E.M.W.; Pool, R.; Floegel, A.; Deelen, J.; Much, D.; Prehn, C.; Breier, M.; et al. Metabolite Ratios as Potential Biomarkers for Type 2 Diabetes: A DIRECT Study. Diabetologia 2018, 61, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where Do We Stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Phapale, P. Pharmaco-metabolomics Opportunities in Drug Development and Clinical Research. Anal. Sci. Adv. 2021, 2, 611–616. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, H. Perspectives from Metabolomics in the Early Diagnosis and Prognosis of Gestational Diabetes Mellitus. Front. Endocrinol. 2022, 13, 7191. [Google Scholar] [CrossRef] [PubMed]
- Mickiewicz, B.; Vogel, H.J.; Wong, H.R.; Winston, B.W. Metabolomics as a Novel Approach for Early Diagnosis of Pediatric Septic Shock and Its Mortality. Am. J. Respir Crit. Care Med. 2013, 187, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Kaddurah-Daouk, R.; Krishnan, K.R.R. Metabolomics: A Global Biochemical Approach to the Study of Central Nervous System Diseases. Neuropsychopharmacology 2009, 34, 173–186. [Google Scholar] [CrossRef]
- Mock, A.; Zschäbitz, S.; Kirsten, R.; Scheffler, M.; Wolf, B.; Herold-Mende, C.; Kramer, R.; Busch, E.; Jenzer, M.; Jäger, D.; et al. Serum Very Long-Chain Fatty Acid-Containing Lipids Predict Response to Immune Checkpoint Inhibitors in Urological Cancers. Cancer Immunol. Immunother. 2019, 68, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Paglia, G.; Williams, J.P.; Menikarachchi, L.; Thompson, J.W.; Tyldesley-Worster, R.; Halldórsson, S.; Rolfsson, O.; Moseley, A.; Grant, D.; Langridge, J.; et al. Ion Mobility Derived Collision Cross Sections to Support Metabolomics Applications. Anal. Chem. 2014, 86, 3985–3993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paglia, G.; Kliman, M.; Claude, E.; Geromanos, S.; Astarita, G. Applications of Ion-Mobility Mass Spectrometry for Lipid Analysis. Anal. Bioanal. Chem. 2015, 407, 4995–5007. [Google Scholar] [CrossRef] [PubMed]
- Paglia, G.; Smith, A.J.; Astarita, G. Ion Mobility Mass Spectrometry in the Omics Era: Challenges and Opportunities for Metabolomics and Lipidomics. Mass Spectrom. Rev. 2022, 41, 722–765. [Google Scholar] [CrossRef]
- Heuillet, M.; Bellvert, F.; Cahoreau, E.; Letisse, F.; Millard, P.; Portais, J.-C. Methodology for the Validation of Isotopic Analyses by Mass Spectrometry in Stable-Isotope Labeling Experiments. Anal. Chem. 2018, 90, 1852–1860. [Google Scholar] [CrossRef]
- Winter, G.; Krömer, J.O. Fluxomics—Connecting ‘omics Analysis and Phenotypes. Environ. Microbiol. 2013, 15, 1901–1916. [Google Scholar] [CrossRef]
- Emwas, A.-H.; Szczepski, K.; Al-Younis, I.; Lachowicz, J.I.; Jaremko, M. Fluxomics—New Metabolomics Approaches to Monitor Metabolic Pathways. Front. Pharmacol. 2022, 13, 5782. [Google Scholar] [CrossRef]
- Jang, C.; Chen, L.; Rabinowitz, J.D. Metabolomics and Isotope Tracing. Cell 2018, 173, 822–837. [Google Scholar] [CrossRef] [Green Version]
- Eggers, L.F.; Schwudke, D. Shotgun Lipidomics Approach for Clinical Samples. Methods Mol. Biol. 2018, 1730, 163–174. [Google Scholar] [CrossRef]
- Chekmeneva, E.; dos Santos Correia, G.; Chan, Q.; Wijeyesekera, A.; Tin, A.; Young, J.H.; Elliott, P.; Nicholson, J.K.; Holmes, E. Optimization and Application of Direct Infusion Nanoelectrospray HRMS Method for Large-Scale Urinary Metabolic Phenotyping in Molecular Epidemiology. J. Proteom. Res. 2017, 16, 1646–1658. [Google Scholar] [CrossRef]
- Taylor, M.J.; Lukowski, J.K.; Anderton, C.R. Spatially Resolved Mass Spectrometry at the Single Cell: Recent Innovations in Proteomics and Metabolomics. J. Am. Soc. Mass Spectrom. 2021, 32, 872–894. [Google Scholar] [CrossRef] [PubMed]
- Miura, D.; Fujimura, Y.; Wariishi, H. In Situ Metabolomic Mass Spectrometry Imaging: Recent Advances and Difficulties. J. Proteom. 2012, 75, 5052–5060. [Google Scholar] [CrossRef]
- Abu Sammour, D.; Cairns, J.L.; Boskamp, T.; Marsching, C.; Kessler, T.; Ramallo Guevara, C.; Panitz, V.; Sadik, A.; Cordes, J.; Schmidt, S.; et al. Spatial Probabilistic Mapping of Metabolite Ensembles in Mass Spectrometry Imaging. Nat. Commun. 2023, 14, 1823. [Google Scholar] [CrossRef] [PubMed]
- Auricchio, R.; Galatola, M.; Cielo, D.; Amoresano, A.; Caterino, M.; De Vita, E.; Illiano, A.; Troncone, R.; Greco, L.; Ruoppolo, M. A Phospholipid Profile at 4 Months Predicts the Onset of Celiac Disease in At-Risk Infants. Sci. Rep. 2019, 9, 14303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, P.; Carlsson, C.; Virtanen, S.M.; Simell, S.; Hyöty, H.; Ilonen, J.; Toppari, J.; Veijola, R.; Hyötyläinen, T.; Knip, M.; et al. Persistent Alterations in Plasma Lipid Profiles Before Introduction of Gluten in the Diet Associated with Progression to Celiac Disease. Clin. Transl. Gastroenterol. 2019, 10, 44. [Google Scholar] [CrossRef]
- Orešič, M.; Hyötyläinen, T.; Kotronen, A.; Gopalacharyulu, P.; Nygren, H.; Arola, J.; Castillo, S.; Mattila, I.; Hakkarainen, A.; Borra, R.J.H.; et al. Prediction of Non-Alcoholic Fatty-Liver Disease and Liver Fat Content by Serum Molecular Lipids. Diabetologia 2013, 56, 2266–2274. [Google Scholar] [CrossRef] [Green Version]
- Kirchberg, F.F.; Werkstetter, K.J.; Uhl, O.; Auricchio, R.; Castillejo, G.; Korponay-Szabo, I.R.; Polanco, I.; Ribes-Koninckx, C.; Vriezinga, S.L.; Koletzko, B.; et al. Investigating the Early Metabolic Fingerprint of Celiac Disease—A Prospective Approach. J. Autoimmun. 2016, 72, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Martín-Masot, R.; Galo-Licona, J.D.; Mota-Martorell, N.; Sol, J.; Jové, M.; Maldonado, J.; Pamplona, R.; Nestares, T. Up-Regulation of Specific Bioactive Lipids in Celiac Disease. Nutrients 2021, 13, 2271. [Google Scholar] [CrossRef] [PubMed]
- Sedda, S.; Dinallo, V.; Marafini, I.; Franzè, E.; Paoluzi, O.A.; Izzo, R.; Giuffrida, P.; Di Sabatino, A.; Corazza, G.R.; Monteleone, G. MTOR Sustains Inflammatory Response in Celiac Disease. Sci. Rep. 2020, 10, 10798. [Google Scholar] [CrossRef]
- Martín-Masot, R.; Mota-Martorell, N.; Jové, M.; Maldonado, J.; Pamplona, R.; Nestares, T. Alterations in One-Carbon Metabolism in Celiac Disease. Nutrients 2020, 12, 3723. [Google Scholar] [CrossRef] [PubMed]
- Girdhar, K.; Dogru, Y.D.; Huang, Q.; Yang, Y.; Tolstikov, V.; Raisingani, A.; Chrudinova, M.; Oh, J.; Kelley, K.; Ludvigsson, J.F.; et al. Dynamics of the Gut Microbiome, IgA Response, and Plasma Metabolome in the Development of Pediatric Celiac Disease. Microbiome 2023, 11, 9. [Google Scholar] [CrossRef]
- Auricchio, R.; Galatola, M.; Cielo, D.; Rotondo, R.; Carbone, F.; Mandile, R.; Carpinelli, M.; Vitale, S.; Matarese, G.; Gianfrani, C.; et al. Antibody Profile, Gene Expression and Serum Cytokines in At-Risk Infants before the Onset of Celiac Disease. Int. J. Mol. Sci. 2023, 24, 6836. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Leonard, M.M.; Kenyon, V.; Valitutti, F.; Pennacchio-Harrington, R.; Piemontese, P.; Francavilla, R.; Norsa, L.; Passaro, T.; Crocco, M.; et al. Cohort Profile: Celiac Disease Genomic, Environmental, Microbiome and Metabolome Study; a Prospective Longitudinal Birth Cohort Study of Children at-Risk for Celiac Disease. PLoS ONE 2023, 18, e2082739. [Google Scholar] [CrossRef]
- Leonard, M.M.; Valitutti, F.; Karathia, H.; Pujolassos, M.; Kenyon, V.; Fanelli, B.; Troisi, J.; Subramanian, P.; Camhi, S.; Colucci, A.; et al. Microbiome Signatures of Progression toward Celiac Disease Onset in At-Risk Children in a Longitudinal Prospective Cohort Study. Proc. Natl. Acad. Sci. USA 2021, 118, e2020322118. [Google Scholar] [CrossRef]
- Lomash, A.; Prasad, A.; Singh, R.; Kumar, S.; Gupta, R.; Dholakia, D.; Kumar, P.; Batra, V.V.; Puri, A.S.; Kapoor, S. Evaluation of the Utility of Amino Acid Citrulline as a Surrogate Metabolomic Biomarker for the Diagnosis of Celiac Disease. Nutr. Metab. Insights 2021, 14, 603. [Google Scholar] [CrossRef]
- Crenn, P.; Messing, B.; Cynober, L. Citrulline as a Biomarker of Intestinal Failure Due to Enterocyte Mass Reduction. Clin. Nutr. 2008, 27, 328–339. [Google Scholar] [CrossRef]
- Olivares, M.; Albrecht, S.; De Palma, G.; Ferrer, M.D.; Castillejo, G.; Schols, H.A.; Sanz, Y. Human Milk Composition Differs in Healthy Mothers and Mothers with Celiac Disease. Eur. J. Nutr. 2015, 54, 119–128. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.; et al. Duodenal and Faecal Microbiota of Celiac Children: Molecular, Phenotype and Metabolome Characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drabinska, N.; Jarocka-Cyrta, E.; Ratcliffe, N.M.; Krupa-Kozak, U. The Profile of Urinary Headspace Volatile Organic Compounds After 12-Week Intake of Oligofructose-Enriched Inulin by Children and Adolescents with Celiac Disease on a Gluten-Free Diet: Results of a Pilot, Randomized, Placebo-Controlled Clinical Trial. Molecules 2019, 24, 1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drabińska, N.; Azeem, H.A.; Krupa-Kozak, U. A Targeted Metabolomic Protocol for Quantitative Analysis of Volatile Organic Compounds in Urine of Children with Celiac Disease. RSC Adv. 2018, 8, 36534–36541. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Nuclear Magnetic Resonance (NMR) | Mass Spectrometry (MS) | |
|---|---|---|
| Sensitivity | Low | High |
| Dynamic range | Moderate | High |
| Reproducibility | Very high | Moderate |
| Detectable metabolites | 30–100 | 300–5000 or more |
| Metabolite identification | Well categorized | Labor intensive |
| Targeted analysis | Not optimal | Better than NMR |
| Sample destructive | Non-destructive | Destructive to sample |
| Sample preparation | Minimal | More complex than NMR |
| Tissue extraction | Not required | Required |
| Sample analysis time | Fast (<10 min) | Longer than NMR (>10 min) |
| Instrument cost | High | Cheaper than NMR |
| Sample cost | Low | High |
| Study Reference | Groups (N) | Age | Sample | Methodology | Key Results |
|---|---|---|---|---|---|
| [102] | CD progressors (30) vs. HC with similar genetic profiles (20) | 0–8 years | Serum | LC-MS, MRM | Altered serum phospholipid profile, even before gluten intake, in CD progressors: Elevated lyso- and PC and PC-O serum levels; Decreased PE 34:1 and PE 36:1 serum levels. |
| [103] | CD progressors (23) vs. HC matched for HLA risk, sex, and age (23) | 0–6 years | Plasma | MS | Altered serum lipid profile in CD progressors: Elevated TGs of low carbon number and double-bond count plasma levels. Decreased PC, cholesterol esters, endogenous TGs and total essential TG plasma levels (these latter after gluten intake). |
| [105] | CD progressors (33) vs. HC matched for HLA risk (197) | 4 month–8 years | Serum | LC-MS/MS | No significant differences; decreased serum phospholipids levels in CD progressors. No influence of HLA genotype on the serum metabolic profile. |
| [106] | T-CD (17) vs. HC (siblings) (17) | 4–17 years | Plasma | LC-MS/MS | Altered plasma lipid profile in T-CD: elevated carboxylic acids and ceramides, diacylglycerides and lysophospholipid plasma levels. Other altered molecules: fatty acyls, glycerolipids, glycerophospholipids, organoxygen compound, sphingolipids, steroid metabolism, molecules involved in bilirubin metabolism. |
| [108] | T-CD (17) vs. HC (siblings) (17) | 4–17 years | Plasma | LC-MS/MS | Altered one-carbon metabolism in T-CD: Trans-sulphuration pathway down-regulation (decreased cysteine and cystathionine plasma levels), with glutathione and vitamin B6 normal levels. |
| [109] | CD progressors (7 (plasma samples)) vs. HC matched for age, HLA genotype, breastfeeding duration and gluten exposure duration (9 (plasma samples)) | 2.5–5 years | Plasma, stool | MS, GC-MS, LC-MS, HR-MS | Altered plasma cytokine profile (and other metabolites) in CD progressors: Elevated IFNA2, IL-1a, IL-17E/(IL25)), MIP-1b/CCl4, 2-Methyl-3-ketovalric acid, TDCA, Glucono-D-lactone and Isoburyryl-L-carnitine; Decreased oleic acid plasma levels. |
| [113] | T-CD (558) vs. HC (FDRs) (1565) | 1–18 years (T-CD) | Plasma, DBS | LC-MS, DBS | Decreased plasma citrulline levels in T-CD. Decreased plasma citrulline levels in HLA DQ 2.5-positive patients. Inverse correlation between citrulline levels and anti-tTG IgA levels. Value of citrulline levels as predictors of histopathological damage (Marsh 3b and above). |
| Study Reference | Groups (N) | Age | Sample | Methodology | Key Results |
|---|---|---|---|---|---|
| [116] | T-CD (19) vs. HC (15) | 6–12 years | Stool, urine | GC-MS/SPME, H-NMR | Altered VOCs and free-amino-acid levels:
|
| [117] | T-CD Synergy 1 (11) vs. T-CD placebo (12) | 4–18 years | Urine | GC-MS/SPME | No significant changes in VOC urine profiles, except for benzaldehyde concentrations (36% decrease after 12 weeks of intervention). |
| [118] | T-CD (9) vs. HC (9) | 4–14 years | Urine | GC-MS/SPME | Altered VOC levels:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Masot, R.; Jiménez-Muñoz, M.; Herrador-López, M.; Navas-López, V.M.; Obis, E.; Jové, M.; Pamplona, R.; Nestares, T. Metabolomic Profiling in Children with Celiac Disease: Beyond the Gluten-Free Diet. Nutrients 2023, 15, 2871. https://doi.org/10.3390/nu15132871
Martín-Masot R, Jiménez-Muñoz M, Herrador-López M, Navas-López VM, Obis E, Jové M, Pamplona R, Nestares T. Metabolomic Profiling in Children with Celiac Disease: Beyond the Gluten-Free Diet. Nutrients. 2023; 15(13):2871. https://doi.org/10.3390/nu15132871
Chicago/Turabian StyleMartín-Masot, Rafael, María Jiménez-Muñoz, Marta Herrador-López, Víctor Manuel Navas-López, Elia Obis, Mariona Jové, Reinald Pamplona, and Teresa Nestares. 2023. "Metabolomic Profiling in Children with Celiac Disease: Beyond the Gluten-Free Diet" Nutrients 15, no. 13: 2871. https://doi.org/10.3390/nu15132871