
The Effects of Specific Gut Microbiota and Metabolites on IgA Nephropathy—Based on Mendelian Randomization and Clinical Validation
Abstract
:1. Background
2. Method
2.1. Data Source of Exposure and Outcome
2.2. The Selection of Instrumental Variables
2.3. Mendelian Randomization Analysis
2.4. Clinical Validation
2.4.1. Study Population
2.4.2. Sample Collection
2.4.3. DNA Extraction and Sequencing of Samples
2.4.4. Statistical Analysis
3. Results
3.1. The Selection of Instrumental Variables
3.2. Two Samples MR Analysis
3.2.1. Gut Microbiota
3.2.2. Gut Metabolites
3.2.3. Bonferroni-Corrected Test and Reverse Causality
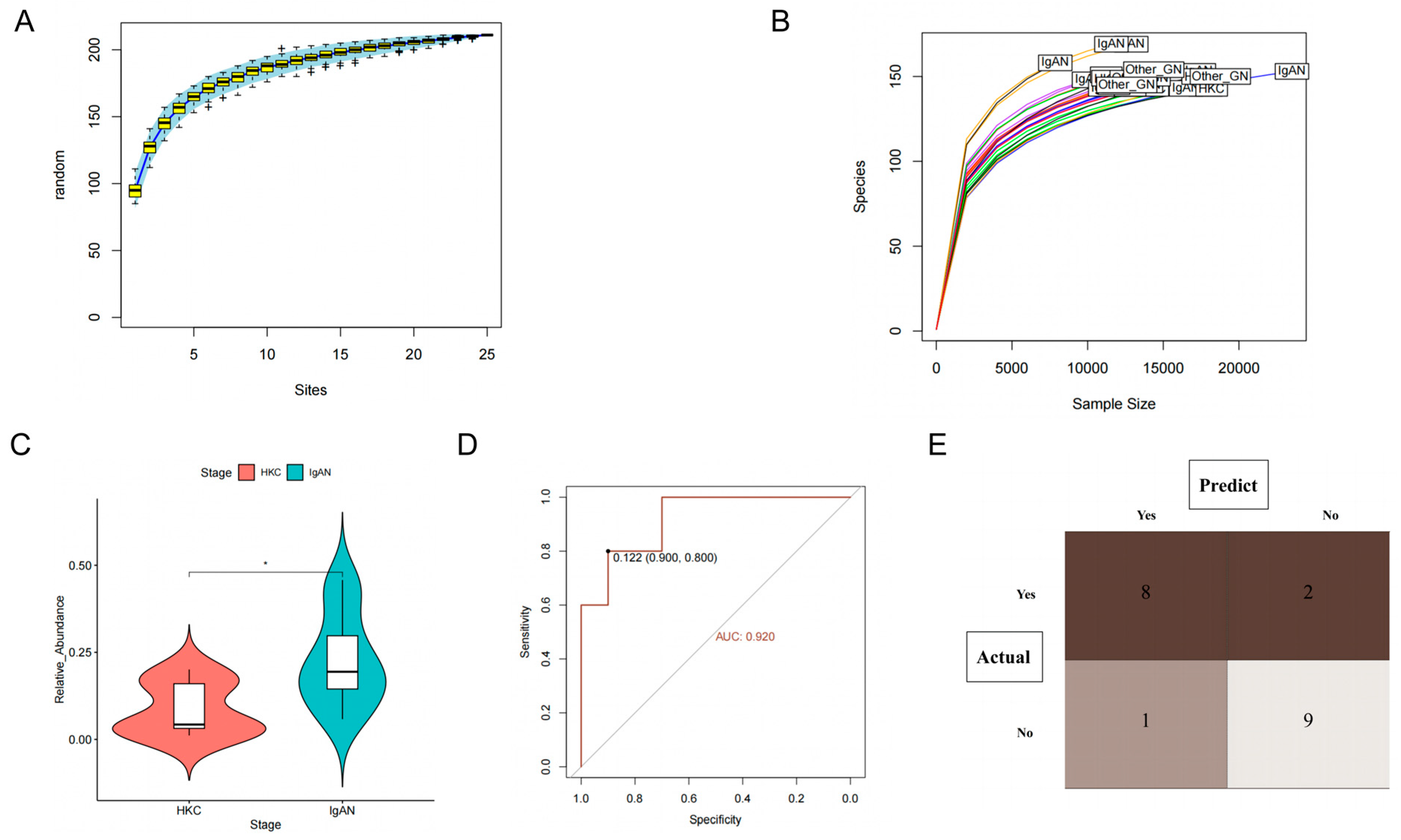
3.3. Clinical Validation
3.3.1. Baseline Characteristic
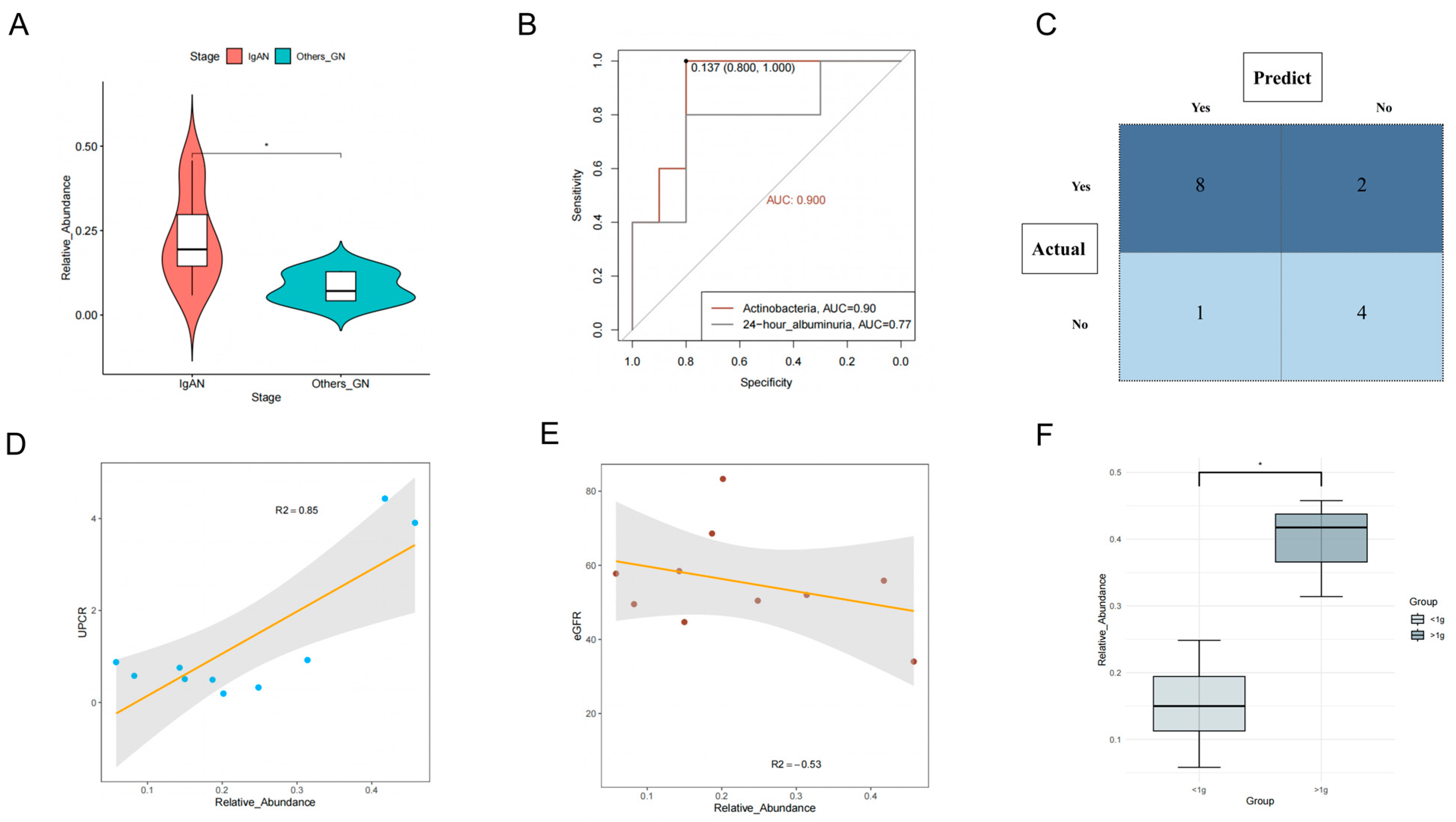
3.3.2. Class Actinobacteria Is Associated with IgAN
3.3.3. Class Actinobacteria Can Be Used to Distinguish IgAN from Other
Glomerular Diseases
3.3.4. Class Actinobacteria Is Associated with the Progression of IgAN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schena, F.P.; Nistor, I. Epidemiology of IgA Nephropathy: A Global Perspective. Semin. Nephrol. 2018, 38, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Canney, M.; Barbour, S.J.; Zheng, Y.; Coppo, R.; Zhang, H.; Liu, Z.-H.; Matsuzaki, K.; Suzuki, Y.; Katafuchi, R.; Reich, H.N.; et al. Quantifying Duration of Proteinuria Remission and Association with Clinical Outcome in IgA Nephropathy. J. Am. Soc. Nephrol. 2021, 32, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Pattrapornpisut, P.; Avila-Casado, C.; Reich, H.N. IgA Nephropathy: Core Curriculum 2021. Am. J. Kidney Dis. 2021, 78, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Coppo, R. The Gut-Renal Connection in IgA Nephropathy. Semin. Nephrol. 2018, 38, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.K.; Rajasekaran, A.; Barratt, J.; Rizk, D.V. An Update on the Current State of Management and Clinical Trials for IgA Nephropathy. J. Clin. Med. 2021, 10, 2493. [Google Scholar] [CrossRef]
- Barratt, J.; Rovin, B.H.; Cattran, D.; Floege, J.; Lafayette, R.; Tesar, V.; Trimarchi, H.; Zhang, H. Why Target the Gut to Treat IgA Nephropathy? Kidney Int. Rep. 2020, 5, 1620–1624. [Google Scholar] [CrossRef]
- Rollino, C.; Vischini, G.; Coppo, R. IgA nephropathy and infections. J. Nephrol. 2016, 29, 463–468. [Google Scholar] [CrossRef]
- Seikrit, C.; Pabst, O. The immune landscape of IgA induction in the gut. Semin. Immunopathol. 2021, 43, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Gesualdo, L.; Di Leo, V.; Coppo, R. The mucosal immune system and IgA nephropathy. Semin. Immunopathol. 2021, 43, 657–668. [Google Scholar] [CrossRef]
- Perše, M.; Večerić-Haler, Ž. The Role of IgA in the Pathogenesis of IgA Nephropathy. Int. J. Mol. Sci. 2019, 20, 6199. [Google Scholar] [CrossRef]
- He, J.W.; Zhou, X.J.; Lv, J.C.; Zhang, H. Perspectives on how mucosal immune responses, infections and gut microbiome shape IgA nephropathy and future therapies. Theranostics 2020, 10, 11462–11478. [Google Scholar] [CrossRef] [PubMed]
- Haniuda, K.; Gommerman, J.L.; Reich, H.N. The microbiome and IgA nephropathy. Semin. Immunopathol. 2021, 43, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Shang, L.; Lu, Y.; Wang, Y. Gut Microbiome Characteristics in IgA Nephropathy: Qualitative and Quantitative Analysis from Observational Studies. Front. Cell. Infect. Microbiol. 2022, 12, 904401. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Chen, D.-Q.; Chen, L.; Liu, J.-R.; Vaziri, N.D.; Guo, Y.; Zhao, Y.-Y. Microbiome–metabolome reveals the contribution of gut–kidney axis on kidney disease. J. Transl. Med. 2019, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Lauriero, G.; Abbad, L.; Vacca, M.; Celano, G.; Chemouny, J.M.; Calasso, M.; Berthelot, L.; Gesualdo, L.; De Angelis, M.; Monteiro, R.C. Fecal Microbiota Transplantation Modulates Renal Phenotype in the Humanized Mouse Model of IgA Nephropathy. Front. Immunol. 2021, 12, 694787. [Google Scholar] [CrossRef]
- Aguilera, M.; Cerdà-Cuéllar, M.; Martínez, V. Antibiotic-induced dysbiosis alters host-bacterial interactions and leads to colonic sensory and motor changes in mice. Gut Microbes 2015, 6, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Davey, S.G.; Hemani, G. Mendelian randomization: Genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 2014, 23, R89–R98. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Garay, J.A.R.; Finnicum, C.T.; Liu, X.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef]
- Rhee, E.P.; Ho, J.E.; Chen, M.-H.; Shen, D.; Cheng, S.; Larson, M.G.; Ghorbani, A.; Shi, X.; Helenius, I.T.; O’donnell, C.J.; et al. A Genome-wide association study of the human metabolome in a community-based cohort. Cell Metab. 2013, 18, 130–143. [Google Scholar] [CrossRef]
- Sakaue, S.; Kanai, M.; Tanigawa, Y.; Karjalainen, J.; Kurki, M.; Koshiba, S.; Narita, A.; Konuma, T.; Yamamoto, K.; Akiyama, M.; et al. A cross-population atlas of genetic associations for 220 human phenotypes. Nat. Genet. 2021, 53, 1415–1424. [Google Scholar] [CrossRef]
- Staley, J.R.; Blackshaw, J.; Kamat, M.A.; Ellis, S.; Surendran, P.; Sun, B.B.; Paul, D.S.; Freitag, D.; Burgess, S.; Danesh, J.; et al. PhenoScanner: A database of human genotype-phenotype associations. Bioinformatics 2016, 32, 3207–3209. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Verbanck, M.; Chen, C.-Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.C.; Berthelot, L. Role of gut–kidney axis in renal diseases and IgA nephropathy. Curr. Opin. Gastroenterol. 2021, 37, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Sugurmar, A.N.K.; Mohd, R.; Shah, S.A.; Neoh, H.-M.; Cader, R.A. Gut microbiota in Immunoglobulin A Nephropathy: A Malaysian Perspective. BMC Nephrol. 2021, 22, 145. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Tan, J.; Tan, L.; Tang, Y.; Qiu, Z.; Pei, G.; Qin, W. Modifications of gut microbiota are associated with the severity of IgA nephropathy in the Chinese population. Int. Immunopharmacol. 2020, 89, 107085. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.; Lu, R.; Li, H.; Pang, Y.; Fu, H.; Fang, G.; Chen, Q.; Liu, B.; Wu, J.; et al. Integrated Fecal Microbiome and Serum Metabolomics Analysis Reveals Abnormal Changes in Rats with Immunoglobulin A Nephropathy and the Intervention Effect of Zhen Wu Tang. Front. Pharmacol. 2021, 11, 606689. [Google Scholar] [CrossRef]
- Liu, F.; Xu, X.; Chao, L.; Chen, K.; Shao, A.; Sun, D.; Hong, Y.; Hu, R.; Jiang, P.; Zhang, N.; et al. Alteration of the Gut Microbiome in Chronic Kidney Disease Patients and Its Association With Serum Free Immunoglobulin Light Chains. Front. Immunol. 2021, 12, 609700. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Qin, Y.; Yu, Z.; Zhang, Y.; Ning, X.; Sun, S. The Specific Alteration of Gut Microbiota in Diabetic Kidney Diseases—A Systematic Review and Meta-Analysis. Front. Immunol. 2022, 13, 908219. [Google Scholar] [CrossRef]
- He, J.-W.; Zhou, X.-J.; Li, Y.-F.; Wang, Y.-N.; Liu, L.-J.; Shi, S.-F.; Xin, X.-H.; Li, R.-S.; Falchi, M.; Lv, J.-C.; et al. Associations of Genetic Variants Contributing to Gut Microbiota Composition in Immunoglobin A Nephropathy. Msystems 2021, 6, e00819-20. [Google Scholar] [CrossRef]
- Shah, N.B.; Nigwekar, S.U.; Kalim, S.; Lelouvier, B.; Servant, F.; Dalal, M.; Krinsky, S.; Fasano, A.; Tolkoff-Rubin, N.; Allegretti, A.S. The Gut and Blood Microbiome in IgA Nephropathy and Healthy Controls. Kidney360 2021, 2, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain–gut–kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef]
- Barba, C.; Soulage, C.O.; Caggiano, G.; Glorieux, G.; Fouque, D.; Koppe, L. Effects of Fecal Microbiota Transplantation on Composition in Mice with CKD. Toxins 2020, 12, 741. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Liebert, A.; Bicknell, B.; Chen, X.-M.; Huang, C.; Pollock, C.A. Faecal Microbiota Transplantation and Chronic Kidney Disease. Nutrients 2022, 14, 2528. [Google Scholar] [CrossRef]
- Luo, S.; Yang, M.; Han, Y.; Zhao, H.; Jiang, N.; Li, L.; Chen, W.; Li, C.; Yang, J.; Liu, Y.; et al. β-Hydroxybutyrate against Cisplatin-Induced acute kidney injury via inhibiting NLRP3 inflammasome and oxidative stress. Int. Immunopharmacol. 2022, 111, 109101. [Google Scholar] [CrossRef] [PubMed]
- Tajima, T.; Yoshifuji, A.; Matsui, A.; Itoh, T.; Uchiyama, K.; Kanda, T.; Tokuyama, H.; Wakino, S.; Itoh, H. β-hydroxybutyrate attenuates renal ischemia-reperfusion injury through its anti-pyroptotic effects. Kidney Int. 2019, 95, 1120–1137. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Jin, X.; Yuan, B.; Lv, Y.; Yan, G.; Liu, M.; Xie, C.; Liu, J.; Tang, Y.; Gao, H.; et al. G Protein-Coupled Receptor 109A Maintains the Intestinal Integrity and Protects Against ETEC Mucosal Infection by Promoting IgA Secretion. Front. Immunol. 2021, 11, 583652. [Google Scholar] [CrossRef]
- Lau, W.L.; Savoj, J.; Nakata, M.B.; Vaziri, N.D. Altered microbiome in chronic kidney disease: Systemic effects of gut-derived uremic toxins. Clin. Sci. 2018, 132, 509–522. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
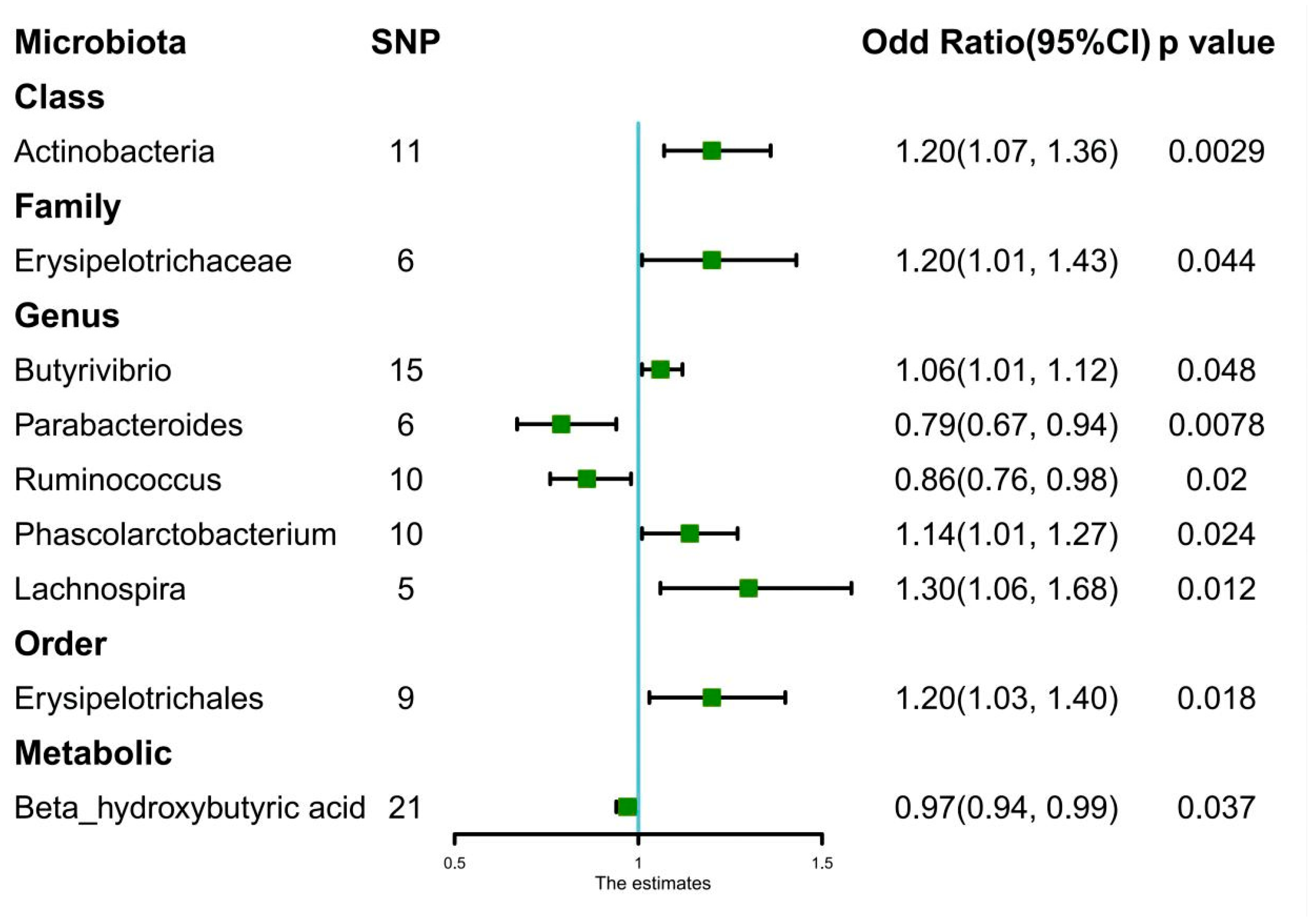
| Exposure | Odd Ratio | 95% CI | p-Value |
|---|---|---|---|
| Class.Actinobacteria | |||
| Weighted mode | 1.27 | 1.05–1.53 | 0.036 * |
| Weighted median | 1.2 | 1.03–1.41 | 0.024 * |
| IVW | 1.2 | 1.07–1.36 | 0.0029 ** |
| MR-Egger | 1.15 | 0.83–1.61 | 0.42 |
| Family.Erysipelotrichaceae | |||
| Weighted mode | 1.27 | 0.88–1.84 | 0.26 |
| Weighted median | 1.2 | 0.95–1.51 | 0.12 |
| IVW | 1.2 | 1.01–1.43 | 0.044 * |
| MR-Egger | 1.34 | 0.61–2.98 | 0.5 |
| Order.Erysipelotrichales | |||
| Weighted mode | 1.21 | 0.88–1.67 | 0.28 |
| Weighted median | 1.2 | 0.99–1.46 | 0.06 |
| IVW | 1.2 | 1.03–1.40 | 0.018 * |
| MR-Egger | 1.32 | 0.68–2.58 | 0.43 |
| Genus.Lachnospira | |||
| Weighted mode | 1.22 | 0.84–1.77 | 0.36 |
| Weighted median | 1.24 | 0.94–1.62 | 0.12 |
| IVW | 1.3 | 1.06–1.68 | 0.012 * |
| MR-Egger | 1.18 | 0.34–4.09 | 0.81 |
| Genus.Parabacteroides | |||
| Weighted mode | 0.81 | 0.61–1.08 | 0.21 |
| Weighted median | 0.81 | 0.64–1.02 | 0.07 |
| IVW | 0.79 | 0.67–0.94 | 0.0078 * |
| MR-Egger | 0.93 | 0.55–1.55 | 0.78 |
| Genus.Butyrivibrio | |||
| Weighted mode | 1.03 | 0.92–1.16 | 0.63 |
| Weighted median | 1.03 | 0.96–1.11 | 0.47 |
| IVW | 1.06 | 1.01–1.12 | 0.048 * |
| MR-Egger | 1.11 | 0.86–1.42 | 0.44 |
| Genus.Phascolarctobacterium | |||
| Weighted mode | 1.16 | 0.93–1.46 | 0.23 |
| Weighted median | 1.13 | 0.98–1.31 | 0.1 |
| IVW | 1.14 | 1.01–1.27 | 0.024 * |
| MR-Egger | 1.1 | 0.67–1.82 | 0.71 |
| Genus.Ruminococcus | |||
| Weighted mode | 0.83 | 0.63–1.05 | 0.21 |
| Weighted median | 0.84 | 0.71–1.01 | 0.055 |
| IVW | 0.86 | 0.76–0.98 | 0.02 * |
| MR-Egger | 0.73 | 0.52–1.01 | 0.13 |
| Beta_hydroxybutyric acid | |||
| Weighted mode | 0.98 | 0.93–1.02 | 0.3 |
| Weighted median | 0.97 | 0.93–1.01 | 0.17 |
| IVW | 0.97 | 0.94–0.99 | 0.037 * |
| MR-Egger | 0.98 | 0.92–1.05 | 0.57 |
| GROUP | HKC | IgAN | Others_GN | p-Value |
|---|---|---|---|---|
| N | 10 | 10 | 5 | |
| Age(years) | 52.4 ± 21.2 | 52.8 ± 17.0 | 67.8 ± 16.5 | 0.205 |
| SBP (mmHg) | 129.2 ± 16.8 | 136.0 ± 15.9 | 132.1 ± 23.6 | 0.8 |
| BMI (Kg/m2) | 23.1 ± 3.9 | 25.0 ± 5.7 | 24.2 ± 3.5 | 0.715 |
| HB (g/L) | 14.2 ± 1.8 | 13.8 ± 1.8 | 11.0 ± 1.2 | 0.016 * |
| ALB (g/L) | 4.3 ± 0.7 | 4.0 ± 0.5 | 3.9 ± 0.4 | 0.093 |
| eGFR (ml/min/1.73 m2) | 93.5 ± 1.9 | 55.5 ± 13.4 | 17.9 ± 10.3 | <0.001 ** |
| 24 h proteinuria | 0.1 ± 0.0 | 1.3 ± 1.6 | 3.1 ± 2.3 | <0.001 ** |
| Gender | 0.867 | |||
| Male | 6 (60.0%) | 7 (70.0%) | 4 (80.0%) | |
| Female | 4 (40.0%) | 3 (30.0%) | 1 (20.0%) | |
| Hypertension | 0.025 | |||
| No | 6 (60.0%) | 1 (10.0%) | 0 (0.0%) | |
| Yes | 4 (40.0%) | 9 (90.0%) | 5 (100.0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Li, N.; Ni, S.; Min, Y.; Wei, K.; Sun, H.; Fu, Y.; Liu, Y.; Lv, D. The Effects of Specific Gut Microbiota and Metabolites on IgA Nephropathy—Based on Mendelian Randomization and Clinical Validation. Nutrients 2023, 15, 2407. https://doi.org/10.3390/nu15102407
Wang F, Li N, Ni S, Min Y, Wei K, Sun H, Fu Y, Liu Y, Lv D. The Effects of Specific Gut Microbiota and Metabolites on IgA Nephropathy—Based on Mendelian Randomization and Clinical Validation. Nutrients. 2023; 15(10):2407. https://doi.org/10.3390/nu15102407
Chicago/Turabian StyleWang, Fang, Ning Li, Siming Ni, Yu Min, Kang Wei, Hongbin Sun, Yuqi Fu, Yalan Liu, and Dan Lv. 2023. "The Effects of Specific Gut Microbiota and Metabolites on IgA Nephropathy—Based on Mendelian Randomization and Clinical Validation" Nutrients 15, no. 10: 2407. https://doi.org/10.3390/nu15102407